添加竹笋对酱油制曲以及成品酱油品质的影响
2020-03-02蒋立新葛青张强章兴肖竹钱毛建卫泮国荣
蒋立新,葛青*,张强 ,章兴,肖竹钱 ,毛建卫,泮国荣
(1.浙江科技学院 生物与化学工程学院 浙江省农产品化学与生物加工技术重点实验室, 浙江省农业生物资源生化制造协同创新中心,杭州 310023; 2.浙江圣氏生物科技有限公司,浙江 湖州 313300)
酿造酱油是以植物蛋白和淀粉等为原料,在适宜的条件下经过一段时间的发酵,从而生产出含有多种氨基酸、碳水化合物等有机物以及具有特殊色、香、味的液体调味品[1]。随着社会不断的发展进步,新型原料作为辅料酿造的酱油开始出现,如詹超群等[2]通过研究添加秘鲁鱿鱼骨粉来酿造酱油,发现在酱油制曲过程中添加适量的鱿鱼骨粉,会促进酱油制曲过程中微生物的生长以及其酶系的分泌,并且会使酱油曲中的挥发性成分高达36.31%。而张昕等[3]研究发现紫苏酱的调味、增香和保健功能更好。同时牟灿灿等[4]用薏仁碎米代替麸皮酿造酱油是可行的,且优于传统发酵酱油。
在传统方法生产的酱油中,无论是制曲还是发酵生产中,微生物起到了无法替代的作用,微生物含有多种酶系,其中米曲霉作为发酵酱油必不可少的菌种,其所分泌的蛋白酶和淀粉酶对酱油酿造过程中所产生的风味和营养具有重要作用[5]。
制曲作为提高酿造酱油质量的关键工序,直接影响酿造酱油的原料利用及品质形成,这一过程主要由可分泌多种酶系的曲霉完成[6]。作为制曲的主要原料性质(高蛋白含量)决定了制曲过程中酶活力的高低,进而决定了制曲的好坏。蛋白酶的主要作用是将原料中的大分子蛋白质分子分解为小分子物质如氨基酸、低分子肽、多肽和胨,进一步被微生物生长所利用;淀粉酶通过将辅料中的淀粉分解成可以被微生物吸收的小分子的葡萄糖、双糖、三糖、糊精等[7],这些被分解成小分子的氨基酸和糖类不仅易于被发酵过程中的微生物吸收,也是酿造酱油中非挥发性物质及挥发性风味物质的前体[8],也为后阶段其他微生物的生长创造了条件。
竹笋营养齐全,一直以来是许多学者研究的热点,竹笋富含各种营养素[9];并且食用竹笋中的蛋白含量高达其干重的30%[10]。前期研究发现:食用竹笋中含有大量蛋白质和丰富的呈味氨基酸,是一类极具开发潜力的蛋白质资源。目前,我国竹笋的产品加工主要集中于咸菜笋、清水笋和笋干等。为缓解竹笋出笋期销售的压力,在产笋旺季时进行适合的加工,能提高笋产品的附加值,将会有很好的市场开发前景。基于这一状况,本文研究了竹笋对发酵酱油的制曲工艺、活性成分和抗氧化性活性等的影响。本研究也首次将竹笋应用于酿造酱油中,这不仅有望为传统酱油提供一种新的酱油酿造思路,而且酱油中还有竹笋中的功能性成分,是一种全新的功能性酱油。
1 材料与方法
1.1 材料
食用竹笋、黄豆、食用麦麸、酱油曲精:市售;福林试剂、干酪素、L-酪氨酸:国药集团化学试剂有限公司;冰乙酸、乙酸钠、甲醛、酒石酸钾钠、没食子酸、芦丁、3,5-二硝基水杨酸、苯酚:分析纯;碘标准液;2,2′-联氮-二(3-乙基-苯并噻唑-6-磺酸)二铵盐(ABTS)。
1.2 设备
HZT-B3002电子称;PTX-FA210电子天平 福州华志科学仪器有限公司;GZX-9140MBE电热恒温鼓风干燥箱、SPX-B-Z生化培养箱 上海博讯实业有限公司医疗设备厂;KQ-300E型超声波清洗器 昆山市超声仪器有限公司;UV-5500PC型紫外分光光度计 上海市元析仪器有限公司;Allegra X-12R型离心机 贝克曼库尔特有限公司;DF-101S磁力加热搅拌器 巩义市予华仪器有限责任公司。
1.3 试验方法
1.3.1 制曲
将大豆和竹笋用清水清洗3遍后,浸泡6~8 h。最后将泡好的大豆、竹笋和食用麦麸一起放入高压灭菌锅内,在121 ℃,0.1 MPa条件下,灭菌30 min后取出,放置过夜。
在放置过夜的黄豆和竹笋中,拌入麦麸,搅拌均匀后加入酱油曲精,进行恒温培养制曲。翻曲的目的是为了避免局部温度过高从而导致的烧曲,使蛋白酶活力下降。最佳翻曲时间为在一定温度下恒温培养12 h,翻曲1次,继续恒温培养12 h后再翻曲1次;然后培养48~72 h,直到制曲原料表面长满黄绿色孢子即制曲成功。
1.3.2 酱醪发酵
采用高盐稀态发酵,添加竹笋制曲发酵的成品酱油为A,不加竹笋制曲发酵的成品酱油为B。将优化好的曲料入罐发酵,添加浓度为20%的盐水,每个样品平行酿造3缸。培养发酵180 d。第1天15 ℃发酵,每天升温1 ℃,达到30 ℃后,持续30 ℃发酵,发酵时间总计6个月[11]。
1.3.3 酶液的提取
向干燥的250 mL锥形瓶中加入成曲10.0 g左右,然后加入磷酸缓冲溶液80 mL (磷酸缓冲溶液pH 7.2),混匀,40 ℃条件下磁力搅拌恒温提取1 h,立即计时,取出后立即沸水浴5 min,通过高温来破坏蛋白酶的活力,水浴过后自然冷却至室温。用4层纱布过滤定容至100 mL,得到粗酶液,离心后取上清液于4 ℃冰箱中待测。
1.3.4 相关酶活力的测定
参考GB 1886/174-2016制作酪氨酸标准曲线,得到线性回归方程:y=0.0105x-0.0007(R2=0.9996)。综合酶活力的测定和淀粉酶活力的测定参考潘雁红等[12]的方法,糖化酶活力和制作葡萄糖标准曲线,得到葡萄糖线性回归方程:y=1.129x+0.0076(R2=0.9975)。
1.4 制曲工艺的单因素试验
将放置过夜的大豆、竹笋、麦麸和酱油曲精按一定比例混合后,恒温培养一定时间后,每隔一段时间取样(本文以间隔时间为12 h),通过测定其蛋白酶活力和其他相应的酶活力来检验其制曲的结果,每个样品做3个平行。考察制曲时间(24,36,48,60,72 h)、制曲温度(28,30,32,34,36 ℃)、酱油曲精添加量(0.10%、0.15%、0.20%、0.25%、0.30%)以及竹笋添加量(5%、10%、15%、20%、40%、60%、80%、100%)对制曲过程中酶活力的影响。
1.5 响应面优化制曲工艺
根据Box-Behnken试验设计及单因素试验结果确定响应面因素水平,利用Design Expert 7.11软件,以制曲时间、制曲温度、酱油曲精添加量、竹笋的添加量进行四因素三水平的响应面试验设计(见表1),考察4个因素之间的相互作用对制曲过程中酶活力的影响,优化制取工艺的最佳配方。制曲的成功与失败可以通过曲料的颜色和曲味等感官因素评定,选择蛋白酶活力为其主要的评价指标。

表1 响应面试验因素与水平Table 1 Factors and levels of response surface experiment
1.6 酱油成品中基本理化指标的测定
根据GB 18186-2000和参考邱露等[13]的方法并略作修改测定氨基酸态氮。利用全自动凯氏定氮仪测定总氮。根据GB 5009.7-2016,利用全自动还原糖测定仪(SGD-IV型)测定还原糖。
1.7 酱油成品中总酚、总黄酮以及抗氧化能力的测定
1.7.1 总酚的测定
参考张欢欢等[14]的方法,略有修改。分别移取1 mL各浓度的没食子酸标准溶液和样品于50 mL离心管中,加入5 mL 10%福林酚试剂,摇匀,3 min后,加入4 mL 7.5% Na2CO3溶液,振荡混匀,120 r/min 25 ℃避光震荡反应1 h。以没食子酸空白为参比溶液,在波长为765 nm分光光度计下读取吸光值,以没食子酸质量浓度为横坐标,吸光值为纵坐标绘制标准曲线。得到回归方程为:y=0.0117x+0.0096(R2=0.999)。
1.7.2 总黄酮的测定
参考郑有飞等[15]的方法,略有修改。取2 mL样品于离心管中,用甲醇稀释至6 mL,摇匀后取上层清液1 mL,加入30%乙醇溶液至5 mL,再加入5%亚硝酸钠溶液0.3 mL,混匀后放置6 min,加入1.0 mol/L氢氧化钠溶液4 mL,用30%乙醇溶液定容至10 mL,放置10 min,以零管为空白,在510 nm的波长处测定吸光度。以芦丁为标品,按照上述方法测定吸光度值,以芦丁质量浓度为横坐标,吸光值为纵坐标绘制标准曲线。得到回归方程为:y=0.1368x+0.0083(R2=0.999)。
1.7.3 抗氧化能力的测定
1.7.3.1 还原力
参考李丹等[16]的方法,略作修改。将5,10,15,20,25 μL样品加磷酸缓冲液(0.2 mol/L,pH 6.6)至2.5 mL,然后加入2.5 mL 1%铁氰化钾(W/V),混合均匀,50 ℃下反应30 min,再加入10%三氯乙酸(W/V) 2.5 mL,混合均匀,静止 10 min,立即取2.5 mL上清液,加入2.5 mL去离子水和0.5 mL 0.1%三氯化铁(W/V),混合均匀,以去离子水为参比溶液,在700 nm下测定。吸光度值越高,还原力越强。
1.7.3.2 ABTS自由基清除试验
参考李莹等[17]的方法,略有修改。取1 mL样品,稀释至50 mL,取50,100,150,200,250,300 μL分别用磷酸缓冲液(5 mmol/L,pH 7.4)补至300 μL,再加入5 mL上述稀释液,在30 ℃下,反应1 h。以去离子水为参比溶液,在734 nm下测定吸光度。
式中:A0为空白对照液的吸光度,即去离子水+ABTS自由基PBS溶液;A1为样品测定管的吸光度,样品+ABTS自由基PBS溶液;A2为样品本底管的吸光度,样品+PBS溶液。
1.8 统计分析
用Origin 8.0软件对数据进行处理和图表的绘制。
2 结果与分析
2.1 单因素试验
2.1.1 制曲时间的确定
制曲时间长短决定了微生物是否得到了充分的生长繁殖,如果制曲时间不足,则会导致微生物达不到最佳的生长效果,也不能够达到对原料的充分利用,致使原料的浪费,其相应的蛋白酶活力也会降低;相反,如果制曲时间过长,会导致其原料的消耗,使微生物在生长发育过程中受阻,同样地其酶活力也会下降,达不到理想的结果,同时增加了原料的消耗。因此,以高蛋白酶活力为依据确定最佳制曲时间。制曲时间对酶活力的影响见图1,在制曲温度为30 ℃、酱油曲精用量为0.20%、竹笋添加量为5%的条件下,在单因素制曲过程中,蛋白酶活力、糖化酶活力和综合酶活力随着制曲时间的增加,呈现出的趋势均为先增大后减小。当制曲时间为60 h时,三者同时达到最大值,分别为669.10 U/g干基,640.14 U/g干基,0.44 U/100 g干基,因此选择最佳的制曲时间为60 h。
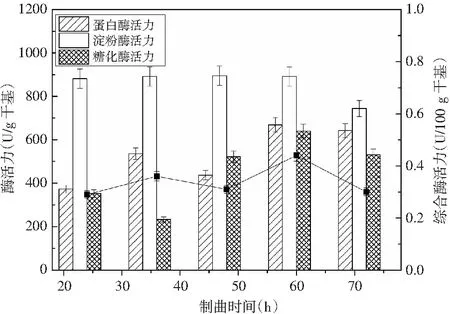
图1 制曲时间对酶活力的影响Fig.1 Effect of koji-making time on enzyme activity
2.1.2 制曲温度的确定
温度是微生物生长繁殖至关重要的因素,一般情况下,微生物的生长温度在低于25 ℃时,其生长代谢都比较缓慢;当其生长温度高于38 ℃时,则非常不利于微生物的生长代谢,反而适合细菌的发育繁殖,并且易染杂菌,影响制曲[18]。选定制曲时间为60 h、接种量为0.2%、竹笋添加量为5%的条件下,考察制曲温度对酶活力的影响。随着制曲温度的逐步升高,蛋白酶活力和糖化酶活力呈现先增大后减小的趋势,而淀粉酶活力和综合酶活力则呈现先增大后减小再增大的趋势,见图2。
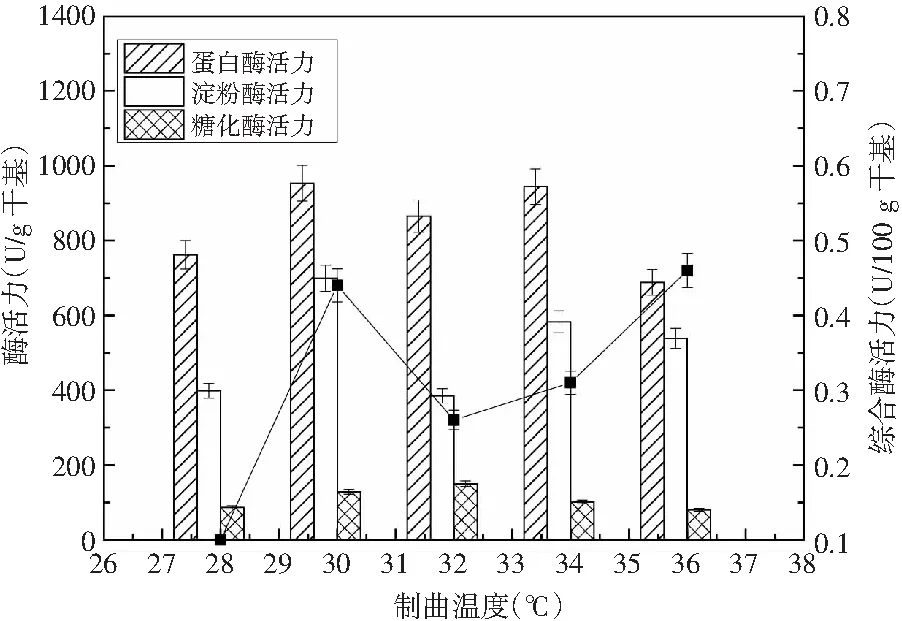
图2 制曲温度对酶活力的影响 Fig.2 Effect of koji-making temperature on enzyme activity
当制曲温度为30 ℃时,蛋白酶活力、淀粉酶活力、糖化酶活力和综合酶活力同时达到最高。因此,选择最佳的制曲温度为30 ℃。
2.1.3 酱油曲精接种量的确定
酱油制曲的周期直接受酱油曲精接种量的影响。酱油曲精接种量大可以缩短制曲的周期,减少被杂菌污染的机率,但如果酱油曲精接种量过大,制曲过程中,其本来用于生长繁殖的主要营养物质会被菌体细胞生长所利用,曲精生长时营养物质的缺乏,从而导致其不能很好地生长繁殖,也会使其酶活力大大地降低,并且在菌体细胞生长利用营养物质的同时会产生大量的代谢废物,使微生物原所处的生长环境得到恶化,菌体细胞也会较早衰老,维持微生物生长所需的营养物质也得不到满足,就会形成大量休眠孢子,降低酶活力;倘若接种量过小,微生物生长繁殖不足,使得营养过剩,造成浪费,增加生产成本[19]。选定制曲时间为60 h、制曲温度为30 ℃、竹笋添加量为5%的条件下,考察曲精量对成曲质量的影响,结果见图3。
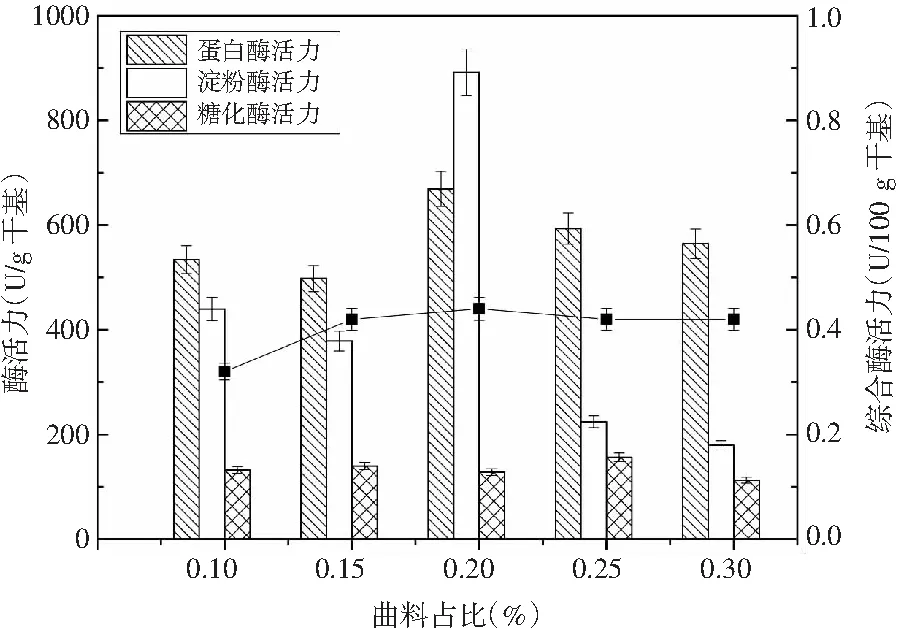
图3 酱油曲精用量对制曲过程中酶活力的影响Fig.3 Effect of the amount of soy sauce koji extract on the enzyme activity in the koji-making process
蛋白酶活力和α-淀粉酶活力随接种曲精量的增加而增大,达到最大值时的曲精量为0.20%,随着接种量的持续增加,酶活力呈逐渐减小趋势。因此,控制接种曲精量为0.20%。
2.1.4 竹笋添加量的确定
竹笋高纤低脂,营养齐全,蛋白含量较高,在制曲过程中竹笋中的蛋白质可被微生物所利用,提高蛋白酶活力,但同时由于竹笋中水分含量高,添加太多对微生物的生长不利,添加太少又达不到利用竹笋制曲的目的。因此选定在制曲时间为60 h、制曲温度为30 ℃、酱油曲精用量为0.20%的条件下,考察竹笋的最佳添加量对成曲质量的影响,结果见图4。
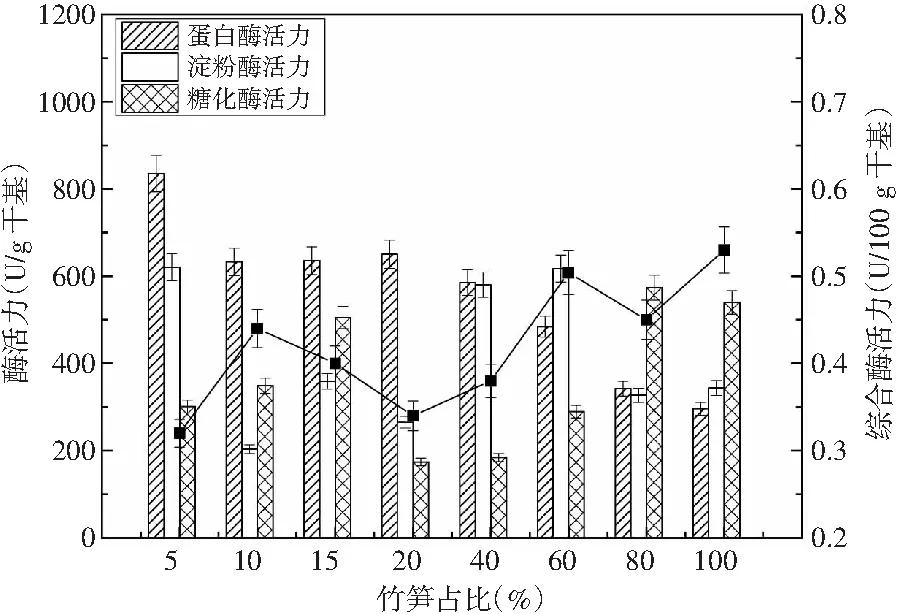
图4 竹笋占比对酶活力的影响Fig.4 Effect of bamboo shoots proportion on enzyme activity
由图4可知,当竹笋添加量为5%时,其蛋白酶活力最高为834.86 U/g干基。当竹笋添加量增大时,其蛋白酶活力明显下降,其下降幅度为24.20%;当竹笋添加量继续增加时,蛋白酶活力会继续减小,减小幅度最大为64.53%。因此,选择最佳的竹笋添加量为5%。
2.2 响应面优化试验
结合单因素试验,以制曲时间(A)、制曲温度(B)、酱油曲精用量(C)和竹笋占比(D)4个条件为自变量,蛋白酶活力为响应值,进行响应面优化,试验设计及结果见表2。

表2 Box-Behnken设计矩阵响应数据结果Table 2 Response data results of Box-Behnken design matrix
2.2.1 回归方程的建立与显著性分析
通过对表2中的试验数据进行回归拟合,得到蛋白酶活力对制曲时间、制曲温度、酱油曲精用量和竹笋占比的回归模型:
R=617.5260-28.89833A-22.04333B-5.35583C-45.84250D-18.42750AB+12.7225AC+73.1200AD+81.54000BC+44.39750BD+9.63000CD-75.32842A2-34.29842B2-79.21717C2-106.45217D2。
表3 回归模型方差分析Table 3 Analysis of variance for the regression model

续 表
注:P<0.01表示差异高度显著,用“**”表示;P<0.05表示差异显著,用“*”表示。
由表3模型方差分析可知,回归方程的各项系数显著性,在制曲过程4个单因素中,对蛋白酶活力具有极显著影响的是B(制曲温度),而对蛋白酶活力具有显著性影响的为A(制曲时间)和C(酱油曲精接种量)。在各交互相中,制曲时间和制曲温度之间的交互作用、制曲温度和酱油曲精接种量之间的交互作用、制曲温度和竹笋占比之间的交互作用对蛋白酶活力有极显著性影响;酱油曲精接种量和竹笋占比之间的交互作用有显著性影响;对于蛋白酶活力值各二次项均有显著影响。
通过对表3中模型方程进行方差分析可知:采用F值对模型的显著性进行分析,F值为22.85,P值<0.0001,该结果显示所选取的模型合适,R2(模型确定系数)=0.9528,表明有95.28%的变化可以用该模型所解释。表明该模型对试验的拟合程度良好,进一步说明该模型对优化制曲条件的试验很合适[20],因此在预测和分析酿造酱油制曲过程中蛋白酶活力变化值时,该模型是可以使用的。模型值为22.85,P值<0.0001,具有高度显著性;模型调整确定系数RAdj2=0.9161,表明此回归模型与实测值拟合较好,可用于制曲过程中蛋白酶活力的预测及分析。
2.2.2 响应面分析
在研究制曲过程中交互项对蛋白酶活力的影响时,在其他因素不变的情况下,任意选择2个因素观察其对蛋白酶活力的影响,对表2的数据进行拟合,即可得到等高线图和响应面图。制曲时间(A)和制曲温度(B)之间的交互作用(见图5)、制曲温度(B)和接种量(C)之间的交互作用(见图6)、制曲温度(B)和竹笋占比(D)之间的交互作用(见图7)对制曲过程中蛋白酶活力具有显著的影响,这一结果与回归方程的交互项显著性分析结果相符。
可通过响应曲面的响应图对蛋白酶活力产生两两交互影响的试验因素进行评价,进而确定各因素的最佳水平范围,响应曲面顶点附近的区域即为最佳水平范围。如果响应值的大小对于影响因素的改变不是很灵敏,那么该响应面相对应的曲面的坡度应较为平缓,相反,若响应值对于影响因素的改变较敏感,那么该响应面相对应的曲面的坡度应较为陡峭。同时也可以用等高线的形状来说明交互作用对响应值影响的强弱,一般情况下两因素交互作用显著,则其形状接近椭圆形,若交互作用不显著则为圆形。根据回归模型做出相应的响应面图和等高线图,见图5~图7。

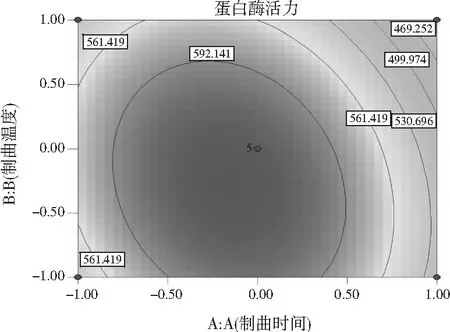
图5 制曲时间和制曲温度之间交互作用的 响应面图和等高线图Fig.5 Response surface and contour maps of the interaction between koji-making time and koji-making temperature


图6 制曲温度和接种量之间交互作用的 响应面图和等高线图Fig.6 Response surface and contour maps of the interaction between koji-making temperature and inoculum amount
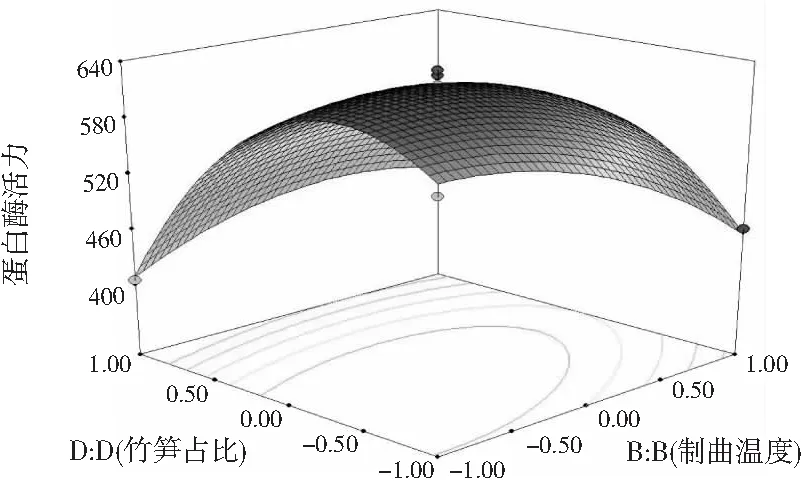
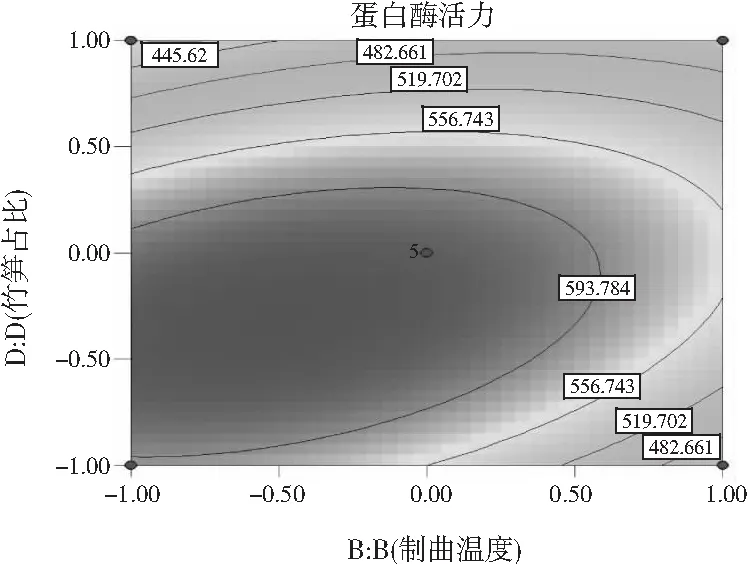
图7 制曲温度和竹笋占比之间交互作用的 响应面图和等高线图Fig.7 Response surface and contour maps of the interaction between koji-making temperature and bamboo shoots proportion
2.3 模型的检验
通过采用蛋白酶活力最大响应值相对应的响应因素值来进行检验预测值与真实值之间的拟合程度以及方程的合适性、有效性,制曲时间62.5 h,制曲温度31.5 ℃,曲精用量0.23%,竹笋占比6.50%为最佳制曲工艺条件,经过3组平行试验,得到其蛋白酶活力658.35 U/g干基,预计值为660.96 U/g干基,与计算值误差<0.39%,说明该模型能良好预测实际情况。
2.4 酱油发酵过程中总氮和氨基酸态氮的变化
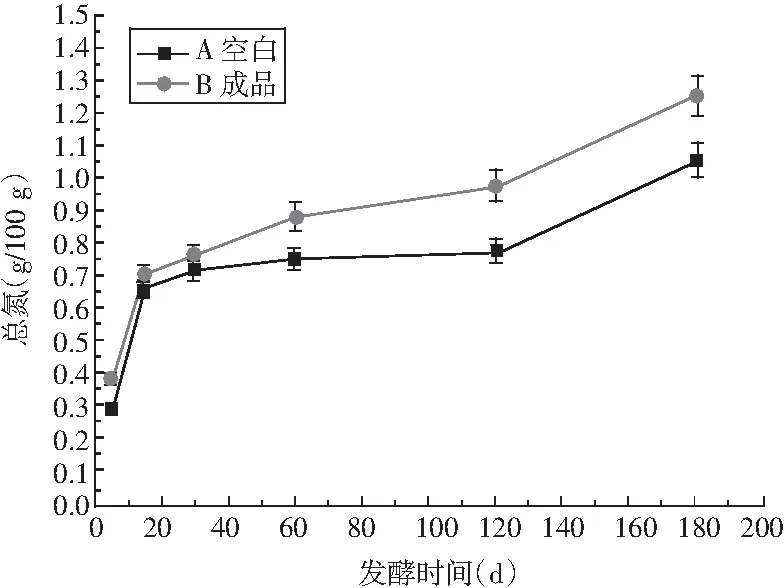
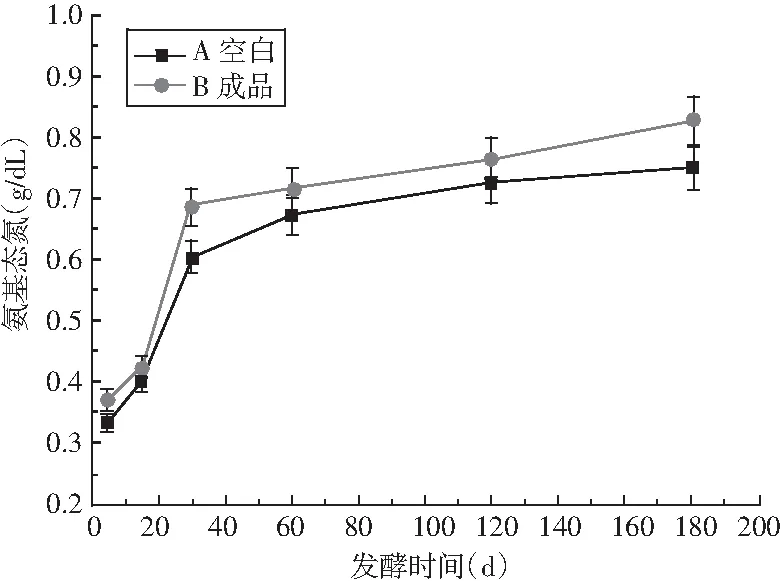
图8 酱油发酵过程中总氮和氨基酸态氮的变化Fig.8 Changes of total nitrogen and amino acid nitrogen in soy sauce during fermentation
酱油在发酵的同时也进行着复杂的生化反应,酱油的品质受这些反应产物的影响。由图8可知,竹笋制酱油的总氮和氨基酸态氮与传统酱油的变化趋势相同,均为在5~30 d内显著增加,之后趋于缓慢增加。试验过程中总氮增加的原因主要是原料蛋白质不断被蛋白酶水解成可溶性的肽类及氨基酸成分[21];酱油A和B总氮含量和氨基酸态氮含量在后2个月的增长速度相同,但A中的总氮与氨酸基态氮含量高于B,氨基酸态氮是原料蛋白质的分解产物,由于酱油A中蛋白质越高,蛋白酶活力越高,其可溶出氮含量就越高。
2.5 总酚和总黄酮含量变化

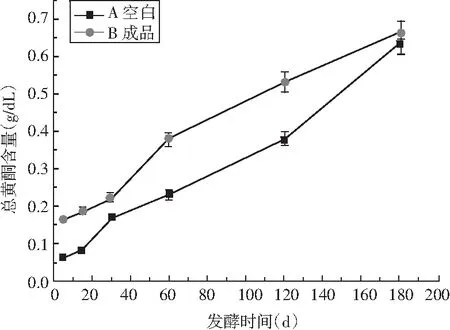
图9 发酵过程中酱油总酚和总黄酮含量含量的变化Fig.9 Changes of total phenolics and flavonoids content in soy sauce during fermentation
多酚类化合物之所以被称为高效的天然氧化物,是因为多酚类化合物中含有多酚羟基,而多酚羟基又具有较强的抗氧化能力以及清除自由基的能力[22],主要包括酚酸、黄酮和单宁类物质等。由图9可知,在整个发酵阶段,前30 d A类酱油和B类酱油中总酚含量迅速增加,在后5个月的发酵过程中,B类酱油呈现缓慢持续增长的趋势,而A类酱油则在第2个月增长速度远大于之后4个月的增加速度。竹笋中含有多酚类物质,在发酵过程中,通过微生物作用,溶进酱油中,说明添加竹笋制曲发酵的酱油高于传统发酵酱油。另外,酱油中酪氨酸含酚羟基,福林酚法测得的酱油总酚含量稍有偏高。总黄酮一直处于稳定增加的趋势,A类酱油比B类酱油中总黄酮含量高,可能是由于添加了竹笋的原因,另外蛋白酶也起到了主导作用,酶活力高,对原料的分解越多,则能够使原料中的黄酮类物质更好地溶解于酱油中。
2.6 发酵过程中酱油抗氧化活性的变化
采用还原力、ABTS法体外抗氧化方法来评价酱油的抗氧化活性,见图10。
由图10可知,成品酱油A和B的抗氧化活性随发酵时间延长而显著上升,主要是因为在发酵过程中,随着发酵的不断进行,原料中酚类、多肽类等活性物质不断溶出,美拉德产物、呋喃酮等活性物质也不断生成。在2种抗氧化指标中,还原力和ABTS在整个发酵过程中A类成品酱油抗氧化活性一直高于B类酱油,造成该现象的主要原因为酱油中对不同抗氧化评价方法相应的抗氧化活性物质不同。该研究结果与刘静怡等[23]的研究结果类似。
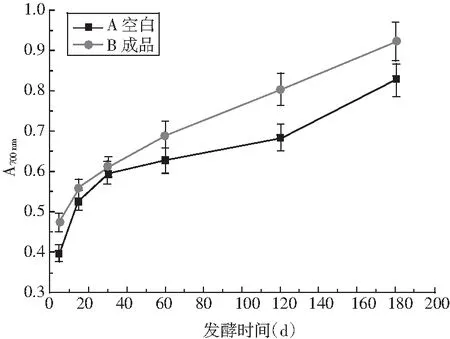
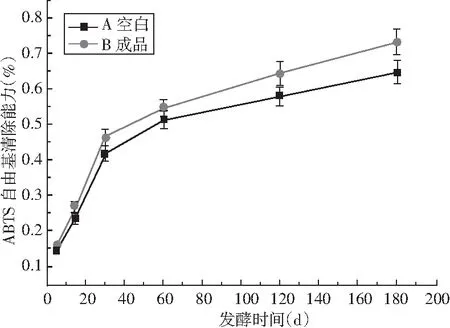
图10 发酵过程中酱油抗氧化活性的变化Fig.10 Changes of antioxidant activity of soy sauce during fermentation
3 结果与讨论
在本试验制曲过程中,制曲因素(制曲时间、制曲温度、曲精量和竹笋添加量)对制曲过程中蛋白酶活力均有显著影响,制曲因素中影响蛋白酶活力的顺序为制曲温度>制曲时间>曲精量>竹笋添加量,而且各因素存在一定的交互作用。
以酱油曲精为发酵菌种,在单因素的基础上,通过响应面分析法研究了酿造酱油制曲过程中添加竹笋的最优制曲工艺条件为制曲时间62.5 h,制曲温度31.50 ℃,接种曲精量0.23%,竹笋占比6.50%,在此条件下蛋白酶活力最高可达658.35 U/g干基。
在制曲的最优条件下,采用高盐稀态工艺酿造竹笋酱油,研究发现竹笋添加能不同程度地提高酱油总氮、氨基酸态氮、总酚、总黄酮含量,其增加百分比分别为9.24%、15.96%、4.55%、3.83%。经体外抗氧化评价方法证实,与传统酱油相比,竹笋酱油抗氧化活性随着发酵时间延长不断上升。在成品竹笋酱油中其还原力和ABTS清除自由基能力提高百分比分别为10.08%和11.63%。 因此,在高盐稀态酱油酿造过程中,竹笋可部分替代黄豆,作为功能性更强的新型酱油,该研究为新产品酱油的研发提供了一定的理论支持。
