转录因子Krüppel样因子15通过下调活化T细胞核因子胞质1型保护足细胞
2019-12-31窦曹帅柯贵宝连智雯陈雪芹1梁馨刘双信1
窦曹帅 张 鸿 柯贵宝 连智雯 陈雪芹1, 史 伟 梁馨 刘双信1,
蛋白尿通常与足细胞损伤密切相关[1],足细胞损伤及数量减少可直接导致蛋白尿和肾小球硬化[2],但其损伤机制尚不完全清楚。Krüppel样因子15(KLF15)是KLFs转录因子家族成员,C端包含与DNA区域连接的锌指结构,N端是一段高度保守的区域[3],在肾脏中表达丰富[4],国内外研究发现,KLF15可减弱足细胞损伤[5],缺少KLF15的小鼠加速慢性足细胞损伤进展[6],糖皮质激素可显著提高足细胞中KLF15的表达,并促进足细胞分化[7],但具体分子机制尚不明确。
核转录因子——活化T细胞核因子胞质1型(Nuclear factor of activated T cytoplasmic 1,NFATc1)静息状态下以磷酸化形式存在于胞质中,Ca2+流入后活化钙调磷酸酶介导NFATc1脱磷酸入核介导下游基因转录,从而引起足细胞损伤[8]。我们课题组既往研究发现,NFATc1活化在足细胞损伤、凋亡以及肾小球硬化的发生发展中发挥着至关重要的作用。近来有文献报道,糖皮质激素可显著提高KLF15的表达,并通过生物信息学分析预测发现KLF15与NFATc1启动子区域有结合[7]。因此,本研究拟通过体外培养足细胞实验探究KLF15是否通过影响NFATc1保护足细胞,并进一步完善糖皮质激素保护足细胞的分子机制。
材料和方法
实验试剂RMPI 1640培养基(CORNING);重组小鼠γ干扰素(IFN-γ)(ProSpec);胎牛血清(FBS);0.05%胰酶(Gibco);核浆蛋白提取试剂盒、凋亡试剂盒(凯基);ChIP试剂盒、荧光二抗488、荧光二抗555(Thermo);NFATc1抗体(Abcam);GAPDH 抗体(Bioworld);BAX、KLF15抗体(Santa);BCL2、Histone 抗体(CST);逆转录试剂盒、Trizol、lipofectamine 2000(LifeTechnologies);SYBR GREEN(Takara);增强绿色荧光蛋白(EGFP)。
细胞培养条件永生化小鼠足细胞系由美国Jochen Reiser教授(Rush University Medical Center,Chicago,IL,USA)惠赠。足细胞复苏后在含10% FBS 及(2~10)×104U/L 的 IFN-γ培养基、33 ℃含5%CO2培养箱中增殖,细胞融合达到70%~80%时转入 5% CO2、37 ℃培养箱中在5% FBS 及不含IFN-γ的培养基中分化,在37 ℃培养10~12d,足细胞分化成熟。本研究所做实验均在分化成熟后进行。
实验分组
免疫荧光分组 肾癌患者周边正常肾组织(Normal组);糖尿病肾病患者肾穿组织标本(DN组);微小病变型肾病患者肾穿组织标本(MCD组);局灶节段性肾小球硬化患者肾穿组织标本(FSGS组)。
足细胞损伤刺激分组 CON组:空白对照组;脂多糖(LPS)组:含100 μg/ml的LPS培养基培养48h;HG组:含30 mmol/L葡萄糖的培养基培养48h;阿霉素(ADR)组:含0.25 μg/ml ADR培养基培养24h。
足细胞过表达及过表达损伤处理分组 EGFP组:腺病毒瞬时感染过表达对照组;KLF15组:腺病毒瞬时感染过表达KLF15组;EGFP+LPS组:过表达对照后加入LPS干预24h;EGFP+HG组:过表达对照后加入高糖干预24h;EGFP+ADR组:过表达对照后加入ADR干预24h;KLF15+LPS组:过表达KLF15后加入LPS干预24h。KLF15+HG组:过表达KLF15后加入高糖干预24h;KLF15+ADR组:过表达KLF15后加入ADR干预24h。
足细胞地塞米松处理分组 0h:空白对照组;6h:地塞米松(10 μmol/L)处理6h组;12h:地塞米松处理12h组;24h:地塞米松处理24h组;48h:地塞米松处理48h组。
病毒感染足细胞分化成熟后,在5 ml不含血清RMPI 1640培养基中加入5 μl病毒,24h后换为含5%血清的RMPI 1 640培养基,再做其他干预。
地塞米松处理8 ml 5%血清的RMPI 1 640培养基加入2 μl 40 mmol/L地塞米松后培养6h、12h、24h、48h。
WesternBlot 细胞干预到时间后,每皿加入100 μl蛋白裂解液刮取蛋白,BCA法测得蛋白浓度后按照20∶ 1加入变性液,98℃变性10 min,按照每孔60 μg上样蛋白,80V恒压电泳分离后,200 mA恒流120 min湿转至PVDF膜上,用5%牛奶封闭1h,一抗4℃孵育过夜,用相应二抗4℃孵育2h,用电化学发光液(ECL)曝光,Image J软件分析灰度值。
实时定量PCR(RT-PCR)Trizol法提取细胞总RNA,按照逆转录试剂盒说明逆转录RNA为cDNA,按照SYBR-Green PCR试剂盒说明检测目的基因表达。引物序列:GAPDH正义链5′-A ̄G ̄G ̄T ̄C ̄G ̄G ̄T ̄G ̄T ̄G ̄A ̄A ̄C ̄G ̄G ̄A ̄T ̄T ̄T ̄G-3′,反义链5′-T ̄G ̄T ̄A ̄G ̄A ̄C ̄C ̄A ̄T ̄G ̄T ̄A ̄G ̄T ̄T ̄G ̄A ̄G ̄G ̄T ̄C ̄A-3; NFATc1正义链5’-G ̄G ̄A ̄G ̄A ̄G ̄T ̄C ̄C ̄G ̄A ̄G ̄A ̄A ̄T ̄C ̄G ̄A ̄G ̄A ̄T-3’反义链5’-T ̄T ̄G ̄C ̄A ̄G ̄C ̄T ̄A ̄G ̄G ̄A ̄A ̄G ̄T ̄A ̄C ̄G ̄T ̄C ̄T-3’;KLF15正义链5’-G ̄A ̄G ̄A ̄C ̄C ̄T ̄T ̄C ̄T ̄C ̄G ̄T ̄C ̄A ̄C ̄C ̄G ̄A ̄A ̄A-3’,反义链5’-G ̄C ̄T ̄G ̄G ̄A ̄G ̄A ̄C ̄A ̄T ̄C ̄G ̄C ̄T ̄G ̄T ̄C ̄A ̄T-3’;BAX正义链5’-A ̄G ̄A ̄C ̄A ̄G ̄G ̄G ̄G ̄C ̄C ̄T ̄T ̄T ̄T ̄T ̄G ̄C ̄T ̄A ̄C-3’反义链5’-A ̄A ̄T ̄T ̄C ̄G ̄C ̄C ̄G ̄G ̄A ̄G ̄A ̄C ̄A ̄C ̄T ̄C ̄G-3’; BCL2 正义链5’-G ̄C ̄T ̄A ̄C ̄C ̄G ̄T ̄C ̄G ̄T ̄G ̄A ̄C ̄T ̄T ̄C ̄G ̄C-3’反义链 5’-C ̄C ̄C ̄C ̄A ̄C ̄C ̄G ̄A ̄A ̄C ̄T ̄C ̄A ̄A ̄A ̄G ̄A ̄A ̄G ̄G ̄F ̄Z ̄D9-3’;FZD9正义链5’-C ̄G ̄C ̄A ̄C ̄G ̄C ̄A ̄C ̄T ̄C ̄T ̄G ̄T ̄A ̄T ̄G ̄G ̄A ̄G-3’,反义链 5’-G ̄C ̄C ̄G ̄A ̄G ̄A ̄C ̄C ̄A ̄G ̄A ̄A ̄C ̄A ̄C ̄C ̄TC-3’;RCAN1正义链 5’-C ̄T ̄C ̄C ̄T ̄C ̄C ̄C ̄G ̄T ̄T ̄G ̄G ̄C ̄T ̄G ̄G ̄A ̄A ̄A-3’,反义链 5’-C ̄T ̄G ̄G ̄G ̄A ̄G ̄T ̄G ̄G ̄T ̄G ̄T ̄C ̄T ̄G ̄T ̄C ̄G ̄C-3’; PLARU正义链 5’-G ̄A ̄C ̄T ̄A ̄C ̄C ̄G ̄T ̄G ̄C ̄T ̄T ̄C ̄G ̄G ̄G ̄A ̄A ̄T ̄G -3’,反义链5’-A ̄T ̄G ̄G ̄T ̄C ̄C ̄T ̄G ̄T ̄T ̄G ̄G ̄T ̄C ̄T ̄T ̄T ̄T ̄C ̄G-3’。
免疫荧光肾组织先进行冰甲醇固定15~20 min、0.5% trionx-100透化10 min、5%BSA封闭30 min,再孵育一抗4℃过夜,然后用相应的荧光二抗避光孵育15 min,DAPI染核15 min,封片,共聚焦显微镜下观察,所用图片均在400倍油镜下拍摄。
染色质免疫共沉淀(chromatin Immunoprecipitation,CHIP):甲醛处理并收集足细胞,超声破碎,然后加入KLF15抗体,与KLF15蛋白-NFATc1 DNA复合物相互结合,加入Protein A结合KLF15抗体-KLF15蛋白-NFATc1 DNA复合物,并沉淀,清洗,得到富集的KLF15蛋白-NFATc1 DNA复合物,加热解交联,纯化富集的NFATc1 DNA片断通过PCR分析,PCR特异扩增NFATc1启动子-984/-1 861 bp区域,引物序列NFATc1正义链5’-T ̄C ̄C ̄C ̄C ̄A ̄A ̄G ̄G ̄T ̄C ̄T ̄C ̄G ̄G ̄G ̄T ̄G ̄G ̄G-3’,反义链5’-T ̄T ̄C ̄C ̄G ̄T ̄C ̄C ̄C ̄C ̄G ̄C ̄C ̄C ̄T ̄T ̄C ̄C ̄C ̄T-3’。
流式细胞仪检测细胞凋亡收集各组足细胞,用Binding Buffer重悬足细胞,在足细胞悬液中先加入Annexin V-APC避光孵育15 min,再加入PI后,在1h内用流式细胞仪检测细胞凋亡。
统计学方法利用《SPSS 22.0》进行统计分析,符合正态分布计量资料用均数±标准差表示,组间比较用单因素方差分析,两组间比较用LSD检验,以P<0.05为差异有统计学意义。
结 果
KLF15在肾小球疾病患者的足细胞中表达降低用免疫荧光染色通过synaptopodin定位足细胞,观察正常肾小球足细胞与病变肾小球足细胞中KLF15的表达,发现与正常肾小球相比,病变肾小球足细胞中的KLF15的表达明显降低(图1)。
足细胞KLF15在损伤刺激后表达明显降低足细胞用含0.25 μg/ml ADR、含30 mmol/L葡萄糖、含100 μg/L LPS的培养基分别培养24h、48h、48h,通过RT-PCR和Western Blot检测KLF15的mRNA和蛋白表达,发现与空白对照组相比,ADR组、LPS组以及HG组中KLF15的表达明显降低(图2)。
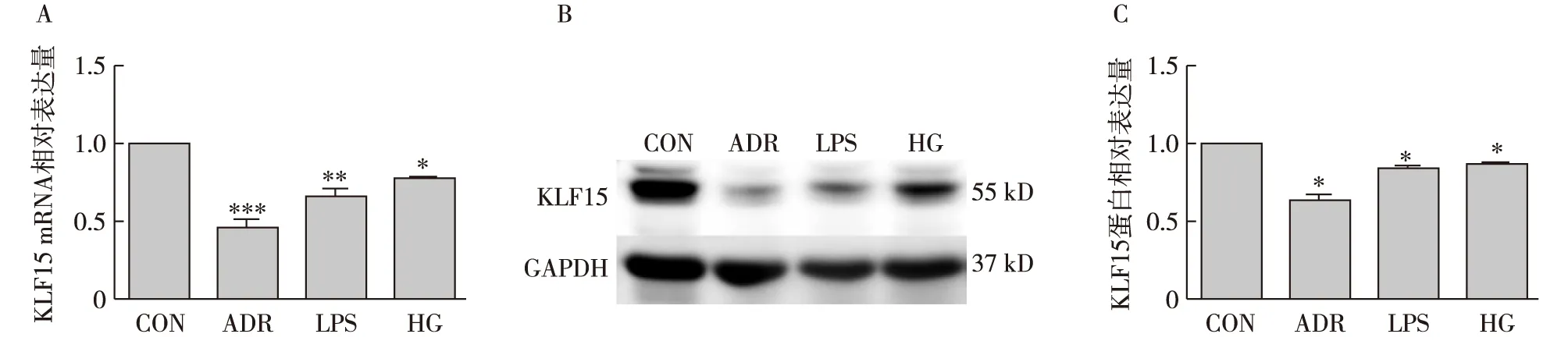
图2 KLF15在ADR、LPS、HG刺激体外培养的足细胞中表达降低A:KLF15的mRNA水平表达;B、C:KLF15的蛋白水平表达;KLF15:Krüppel样因子15;CON:空白对照组;ADR:阿霉素;LPS:脂多糖;HG:高糖;*:与CON组相比P<0.05,**: 与CON组相比P<0.01,***: 与CON组相比P<0.001
过表达KLF15对足细胞的保护作用通过腺病毒感染足细胞过表达KLF15后,通过RT-PCR、Western Blot验证KLF15的过表达效率,结果显示过表达效率很高(图3A、B);足细胞在过表达KLF15后通过Western Blot、RT-PCR检测发现与对照组相比,在mRNA及蛋白水平,促凋亡蛋白BAX表达降低(P<0.01),抗凋亡蛋白BCL2表达升高(P<0.05)(图3C、D);分别在含有LPS(100 μg/ml)、ADR(0.25 μg/ml)、HG(30 mmol/L)培养基中培养24h后,检测细胞凋亡情况,结果发现,过表达KLF15组比对照组凋亡减少(P<0.05)见图3E、图3F,过表达KLF15后给予ADR、LPS、HG等培养后与对照组给予ADR、LPS、HG培养相比凋亡减少(P<0.05)(图3G~L)。
转录因子KLF15对NFATc1表达的调控足细胞过表达KLF15后,通过RT-PCR、 western blot检测NFATc1的表达,结果发现,过表达KLF15后NFATc1的表达降低(P<0.05)(图4A、B);足细胞LPS(100 μg/ml)处理48h后,按照ChIP试剂盒的操作流程,通过普通琼脂糖凝胶电泳检测、RT-PCR定量分析发现,转录因子KLF15与NFATc1的启动子区域存在结合,并且在LPS刺激后结合减少(P<0.001)(图4C、D);KLF15可通过结合NFATc1抑制其下游靶基因FZD9、RCAN1、PLAUR表达,从而抑制NFATc1活性(图4E)。
地塞米松通过KLF15保护足细胞的机制有研究发现[7],糖皮质激素可通过升高KLF15的表达促进足细胞分化,本研究通过对足细胞在含10umol/L地塞米松培养基中培养不同时间段后用RT-PCR检测KLF15的表达发现,与对照组相比,在含地塞米松培养基中培养6h、12h、24h、48h后,KLF15的表达明显上调(P<0.05)(图5A);通过RT-PCR及Western Blot检测不同时间点足细胞中NFATc1的表达,发现NFATc1的表达明显下调(P<0.05),其中以24h最为明显(P<0.01)(图5B、C)。
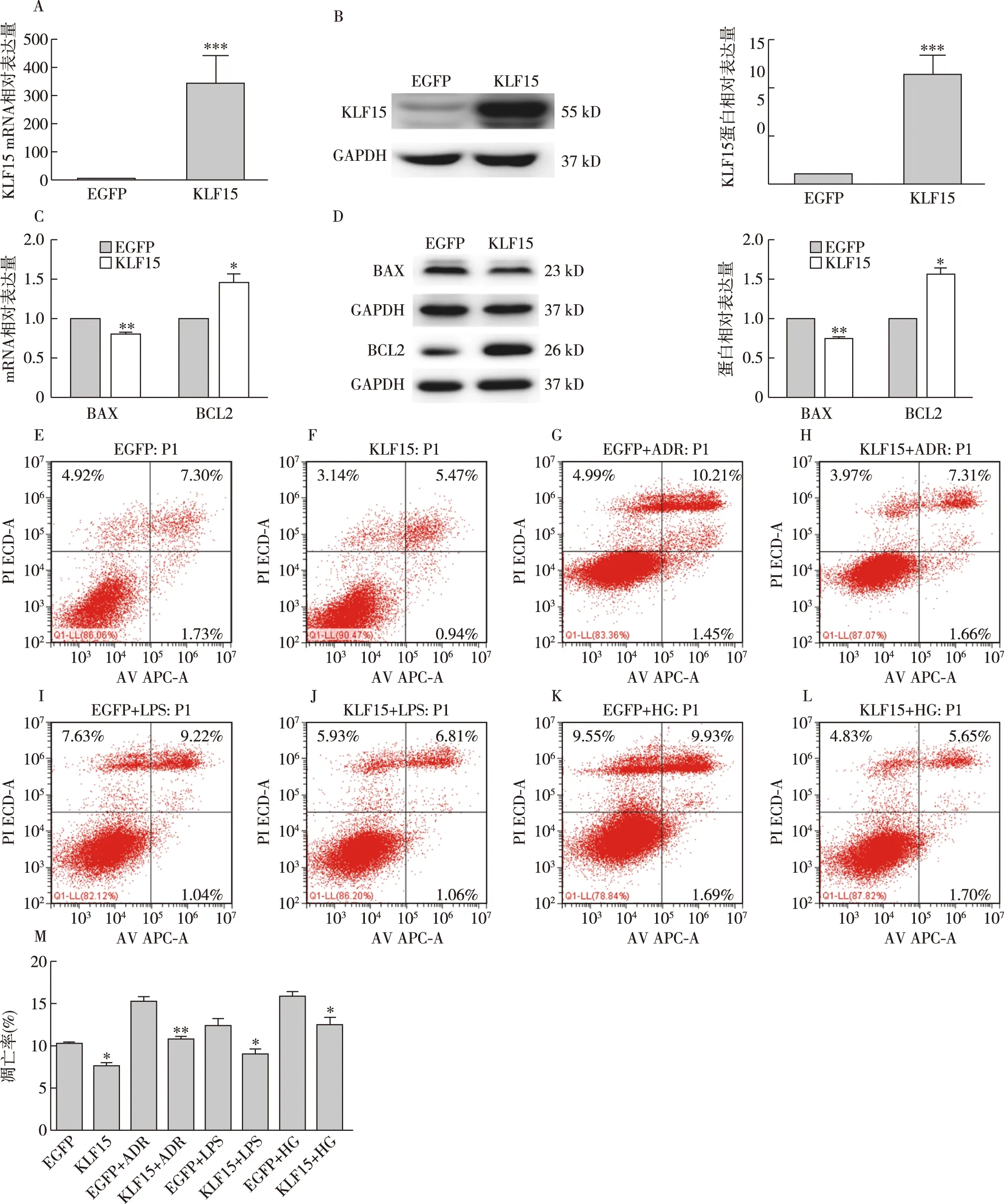
图3 过表达KLF15对足细胞凋亡的影响A、B:mRNA、蛋白水平验证过表达KLF15效率;C:过表达KLF15后BAX、BCL2的mRNA表达;D:过表达KLF15后BAX、BCL2的蛋白表达;E:EGFP组流式凋亡图;F:KLF15组流式凋亡图;G:EGFP+ADR组流式凋亡图;H:KLF15+ADR组流式凋亡图;I:EGFP+LPS组流式凋亡图;J:KLF15+LPS组流式凋亡图;K:EGFP+HG组流式凋亡图;L:KLF15+HG组流式凋亡图;M:细胞凋亡定量统计结果;KLF15:Krüppel样因子15;BAX:促凋亡蛋白;BCL2:抗凋亡蛋白;EGFP:增强绿色荧光蛋白;ADR:阿霉素;LPS:脂多糖;HG:高糖; *:与对照组相比P<0.05,**: 与对照组相比P<0.01,***: 与对照组相比P<0.001
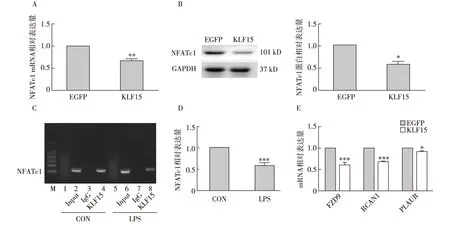
图4 转录因子KLF15对NFATc1表达的调控CON:空白对照组,LPS:脂多糖处理组,Input:阳性对照组,IgG组抗体为IgG(阴性对照组),KLF15组抗体为KLF15;A:RT-PCR示过表达KLF15后NFATc1的mRNA表达;B:Western Blot示过表达KLF15后NFATc1蛋白表达;C:染色质免疫共沉淀(CHIP)后普通琼脂糖凝胶电泳,引物为针对NFATc1的启动子区域序列;D:CHIP针对NFATc1启动子区域进行的RT-PCR定量分析(C、D两者为不同类型实验);E:NFATc1下游靶基因mRNA表达; *:与对照组相比,P<0.05;**:与对照组相比,P<0.01;***:与对照组相比,P<0.001;NFATc1:活化T细胞核因子胞质1型
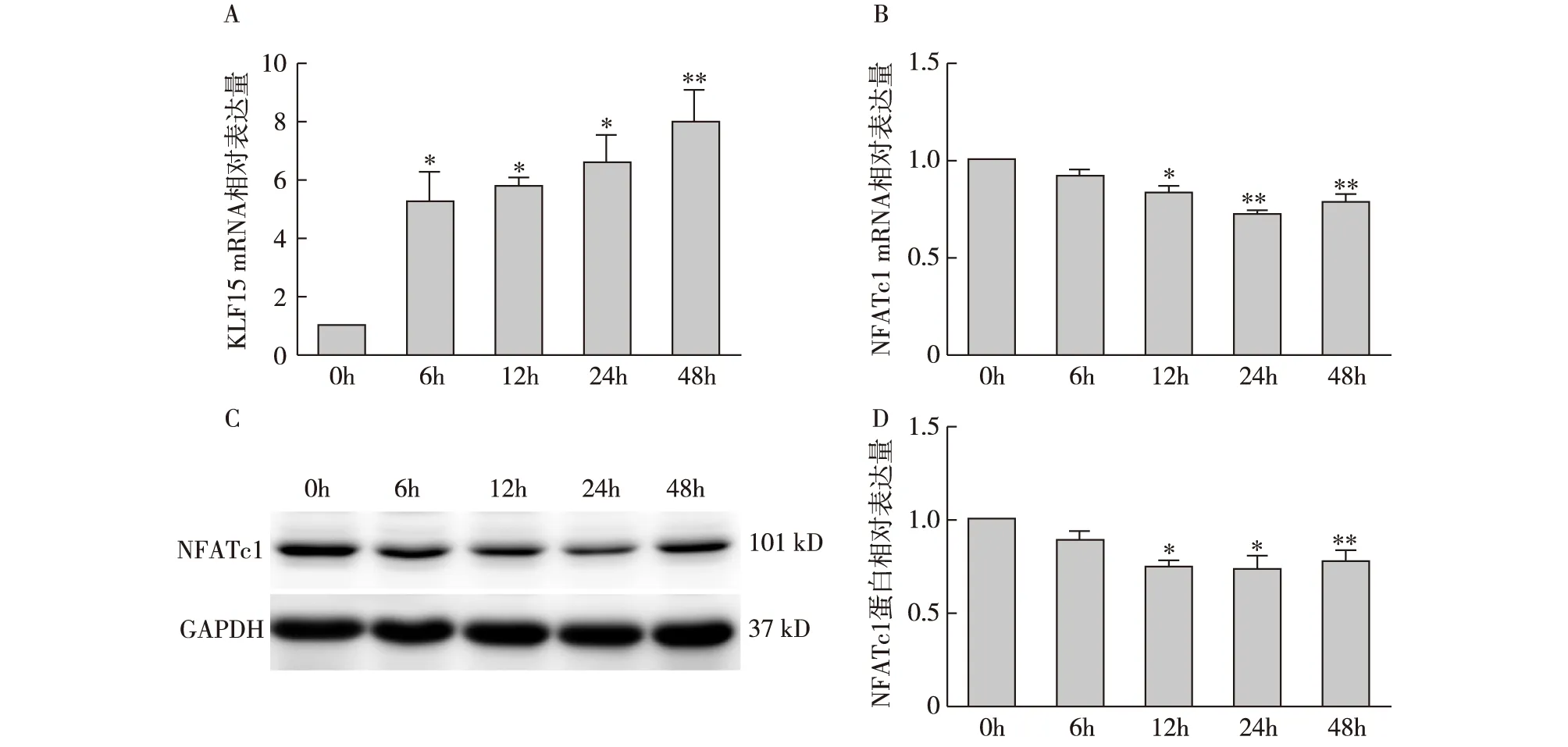
图5 地塞米松处理不同时间点NFATc1、KLF15表达NFATc1:活化T细胞核因子胞质1型;KLF15:Krüppel样因子15;A:地塞米松处理不同时间点足细胞中KLF15的mRNA表达水平;B: 地塞米松处理不同时间点足细胞中NFATc1 mRNA表达;C、D:地塞米松处理不同时间点足细胞中NFATc1的蛋白表达;*:与对照组(0h)相比,P<0.05,**:与对照组(0h)相比,P<0.01
讨 论
蛋白尿是导致终末期肾病的重要危险因素之一[9],足细胞作为肾脏滤过功能的最后屏障,足细胞损伤是蛋白尿产生的重要病理基础。足细胞损伤在糖尿病肾病及其他导致蛋白尿肾病中常表现为足突融合[10],进而导致足细胞的死亡、脱落及数量减少,但足细胞的损伤机制尚不完全清楚,是近年来慢性肾病的研究热点。
KLF15是转录因子Krüppel-like factors家族中的一员,最早发现于果蝇体内,与果蝇的Krüppel蛋白结合[3]。人体内已被发现的KLFs包含KLF1-KLF18,已知KLF2、KLF4、KLF5、KLF6、KLF15参与到肾脏疾病调节中。有研究发现,KLF15是促进果蝇肾原细胞分化的关键调节因子[11];KLF15参与调节足细胞的分化,敲除KLF15的小鼠与野生型相比有更严重的蛋白尿,并且足细胞分化减少[12]。本研究发现在肾小球疾病患者足细胞中KLF15的表达降低;通过给予足细胞ADR、LPS、HG三种损伤处理发现KLF15的表达明显下调;通过对足细胞过表达KLF15后发现,促凋亡蛋白BAX表达降低,抗凋亡蛋白BCL2表达升高,并通过流式细胞术检测发现,过表达KLF15后,足细胞凋亡降低,而且在损伤刺激下的足细胞,过表达KLF15后相比于对照组,凋亡明显下降,进一步说明KLF15可显著减少足细胞损伤,这提示,KLF15在足细胞中是一个保护性因子,而KLF15保护足细胞的具体机制尚不明确。
以往研究发现[13],生理状态的NFAT磷酸化位于细胞浆中,经典的CaN-NFATc1调节通路中,在损伤刺激时,Ca2+流入后活化钙调磷酸酶(CaN)介导NFATc1脱磷酸入核从而介导下游靶基因FZD9、RCAN1、PLARU转录,FZD9、RCAN1的转录可直接导致足细胞功能紊乱[14],PLAUR编码蛋白uPAR增加足细胞的活动性可促进足细胞的脱落,引起足细胞损伤[15];NFATc1的活化可通过升高促凋亡蛋白BAX促进足细胞凋亡[16],导致肾小球硬化[14],除此之外,NFATc1还被发现在糖尿病肾病中活性增强,参与到糖尿病肾病足细胞损伤进程[17];国内外研究及本课题组[8,17-19]发现,RANK,ATF3,SOS2等分子都可通过调节NFATc1影响足细胞损伤;有研究通过生物信息学分析发现[7],KLF15与NFATc1的启动子区域可能存在结合。本研究通过对足细胞过表达KLF15观察NFATc1的表达发现,过表达KLF15的足细胞NFATc1的表达明显下调,并通过CHIP实验证实,KLF15与NFATc1的启动子区域结合,并在损伤刺激下结合减弱,足细胞过表达KLF15后NFATc1的下游靶基因FZD9、RCAN1、PLAUR表达减少,表明KLF15可通过抑制NFATc1的表达而保护足细胞。
糖皮质激素因为其强大的抗炎作用而被广泛用于各种肾脏疾病的治疗中,研究证实,人类足细胞中存在糖皮质激素受体[20],糖皮质激素可以直接对足细胞起作用并对足细胞分化有着重要作用[21-22],但糖皮质激素对于足细胞分化调节的具体分子机制尚不明确。近年研究发现[7],糖皮质激素可通过升高KLF15的表达来促进足细胞分化,在LPS诱导小鼠肾损伤模型中,地塞米松可明显改善小鼠蛋白尿水平,而在KLF15敲除鼠中,地塞米松不能改善小鼠蛋白尿,说明KLF15表达受抑制后地塞米松的治疗效果明显降低,KLF15是地塞米松的直接治疗靶点。本研究中,通过用地塞米松处理足细胞6h、12h、24h、48h发现,KLF15的表达水平显著升高,并且NFATc1的表达降低,表明糖皮质激素有可能通过KLF15-NFATc1通路保护足细胞。
本研究首次探讨了转录因子KLF15通过抑制NFATc1信号通路保护足细胞的可能机制,丰富了NFATc1的分子调节机制,在经典CaN-NFATc1通路之外从基因转录水平探索了NFATc1调节新思路,为临床治疗蛋白尿及慢性肾小球疾病提供新策略;KLF15是否还通过其他分子途径保护足细胞尚待进一步探索;本研究还初步探索了糖皮质激素保护足细胞可能的新的分子机制,丰富了糖皮质激素的作用机制,但糖皮质激素是否通过KLF15抑制NFATc1、糖皮质激素在ADR、HG诱导的足细胞损伤的治疗效果尚待进一步研究。
