转录因子NtMYC2b参与烟草烟碱含量杂种优势形成的机制研究
2019-11-21吴宇瑶杨友成刘坤华
吴宇瑶,谢 瑞,杨友成,刘坤华,聂 琼
(1.贵州大学 农学院,贵州 贵阳 550025; 2.贵州大学 贵州省烟草品种研究重点实验室,贵州 贵阳 550025; 3.遵义市农业科学研究院,贵州 遵义563000; 4.遵义市烟草公司,贵州 遵义 563000)
转录因子(Transcription factor)又名反式作用因子,是一种行使调控基因表达功能且具有特殊结构的蛋白质分子,能特异性地识别并结合顺式元件启动子区域的核心序列,从而调控靶基因的转录[1-2]。转录因子的调控过程是植物信号传导的关键环节,广泛参与植物初生和次生代谢产物合成[3-4]等各种生物学过程,调控植物的生长发育。目前,植物中已经鉴定出的转录因子有多种[5-7],其中MYC2属于bHLH转录因子家族,在茉莉酸(JA)信号传导途径中处于核心地位,发挥着重要作用[8]。ZHANG等[9]在烟草中克隆得到2个MYC2基因,命名为NtMYC2a和NtMYC2b,研究发现,这2个转录因子基因影响尼古丁的合成,在体外也能特异性结合烟碱合成关键酶N-甲基转移酶基因(PMT)启动子的G-box区域,调控PMT基因的表达。
烟碱由2个不同的含氮环即具有2种合成途径的吡咯环和吡啶环合成[10]。在吡咯环途径中,鸟氨酸脱羧酶(ODC)可直接使鸟氨酸脱羧形成腐胺[11],或者间接通过精氨酸脱羧酶(ADC)介导精氨酸脱羧形成腐胺[12]。继而,腐胺被N-甲基转移酶(PMT)催化,形成N-甲基腐胺[13]。最后,N-甲基腐胺被N-甲基腐胺氧化酶(MPO)催化,形成烟碱的直接前体物质。在吡啶环途径,喹啉酸磷酸核糖转移酶(QPT)是烟碱合成代谢中的限速酶[14]。研究表明,PMT过量表达可以增加烟叶中的烟碱含量[15],而通过反义RNA技术沉默PMT表达可以降低烟叶中的烟碱含量[16]。烟碱含量的高低是评价烟草品质的重要指标之一[17],也是将其应用于医药、化工及农药防治[18-20]等方面的依据。杂种优势利用是作物品种改良的一种重要手段,已逐渐成为进一步提高动植物产量、品质、抗病性及社会经济效益的有效措施[21],利用杂种优势选育不同烟碱含量的烟草品种,将其应用于烟叶生产、医药及农药防治有着实践意义。目前,烟碱含量杂种优势的形成机制尚未得到阐明,且在转录因子水平上关于烟碱杂种优势的研究报道还比较少。
转录因子可以作为分子开关与结构基因启动子特异结合,通过调控结构基因的表达,高效诱导或抑制目标代谢产物的积累[22-24]。因此,开展烟碱生物合成相关转录因子及结构基因的表达研究,进一步研究转录因子参与调控烟碱生物合成的分子机制和烟碱含量杂种优势的形成机制,对于提高烟草的品质和工业应用价值具有重要意义。
1 材料和方法
1.1 材料
烟草亲本VA116、GDH88、巴斯玛及其杂交种(VA116×GDH88和VA116×巴斯玛)均由贵州大学烟草实验室提供。其中,VA116×巴斯玛和VA116×GDH88分别为贵州大学烟草实验室在前期研究杂交组配中筛选鉴定出来的烟碱含量杂种优势的强优势组合和弱优势组合。材料栽种于贵州大学烟草试验基地,分别在烟株移栽后66 d和移栽后80 d采样,每次选取长势一致的3株,取其根和中部叶片立即放入液氮冷冻,于-80 ℃冰箱保存备用。
1.2 方法
1.2.1 烟碱含量测定及杂种优势计算 烟碱含量测定采用紫外分光光度法[25]进行,计算公式:
烟碱含量=
式中,1.059是与硅乌酸重量法相比的校正系数,A259、A236、A282分别表示在波长为259、236、282 nm处测得的吸光值,V1、V2、V3分别表示原测液体积、原测液稀释后的待测液体积、空白对照的参比液体积,m表示原测液制备中所称取的样品质量。
杂种优势的计算方法:
正向超亲优势=(F1-HP)/HP×100%,
中亲优势=(F1-MP)/MP×100%,
负向超亲优势=(F1-LP)/LP×100%。
式中,HP为双亲中的最高值,MP为双亲平均值,LP为双亲中的最低值,F1为杂交1代值。
1.2.2 RNA提取与cDNA第一链合成 取不同组织(根与叶)在液氮下充分研磨成粉末,按照OMEGA Totle RNA KitⅠ试剂盒(京工生物公司)说明书提取总RNA。总RNA经过Nanodrop2000紫外分光光度计和琼脂糖凝胶电泳检测浓度和完整性后,根据反转录试剂盒Prime Script TMⅡ1st Strand cDNA Synthesis Kit(TaKaRa)的说明书进行cDNA第一链合成,-20 ℃保存备用。
1.2.3 RT-qPCR与分析 采用RT-qPCR分析NtMYC2b及烟碱合成关键酶基因ODC、PMT、ADC、QPT、MPO的表达,以烟草持家基因Actin-2、HSC70-1为内参基因。各关键酶基因和持家基因的PCR引物参照文献[26]。在NCBI数据库中查找NtMYC2b,根据Real-time PCR引物设计原则,利用Primer Premier 5.0软件设计目的基因引物,上游引物为5′-AGAAGGTTCTACGGGAGC-3′,下游引物为5′-TAACAAACGATTGGGTCA-3′。所有引物由上海生物工程股份有限公司合成。RT-qPCR的反应体系20 μL:Trans Start Green qPCR Super Mix 10 μL,上下游引物各0.8 μL,cDNA模板2 μL,ddH2O 6.4 μL,每个反应重复3次。反应程序为:95 ℃预变性15 s、95 ℃变性10 s,60 ℃退火20 s,72 ℃延伸15 s,40个循环。采用2-△△Ct法计算并分析RT-qPCR的数据。
1.2.4 烟碱合成关键酶基因启动子序列中MYC结合位点分析 利用Plant CARE在线软件分析关键酶基因的启动子序列是否含有MYC结合位点,预测MYC2b调控的酶基因。
1.3 数据处理与分析
所有原始数据均来自3个生物学重复,即重复3次的试验结果。采用 2-△△Ct法计算RT-qPCR的数据,分析NtMYC2b与5个关键酶基因在移栽后66、80 d不同组织(根与中部叶片)中的表达情况。运用Excel和SPSS 21.0软件,统计并分析NtMYC2b的相对表达量、烟碱含量及酶基因表达量间的相关性。
2 结果与分析
2.1 烟草杂交种及其亲本的烟碱含量与杂种优势表现分析
VA116×GDH88和VA116×巴斯玛及其亲本VA116、GDH88、巴斯玛烟株不同时期、不同组织的烟碱含量及其杂种优势表现见表1和表2。从表1可见,移栽后66 d,烟草根中的烟碱含量高于叶中,而移栽后80 d,叶中的烟碱含量高于根中;与移栽后66 d相比,移栽后80 d除VA116根中烟碱含量稍有降低外,其余材料相同部位的烟碱含量均增加;无论不同取样时间,还是不同部位,VA116×巴斯玛的烟碱含量均高于2个亲本,而VA116×GDH88的烟碱含量均介于2个亲本之间。
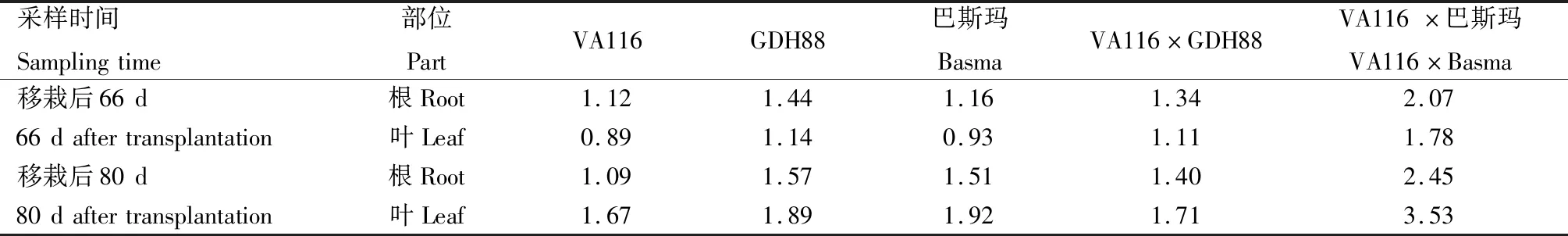
表1 不同时期烟草亲本和杂交种根与叶中的烟碱含量
由表2可知,移栽后66 d和80 d,VA116×巴斯玛和VA116×GDH88根与叶烟碱含量的杂种优势表现一致,且VA116×巴斯玛根与叶中的烟碱含量均表现为正向超亲优势,而VA116×GDH88均表现为弱的中亲优势。
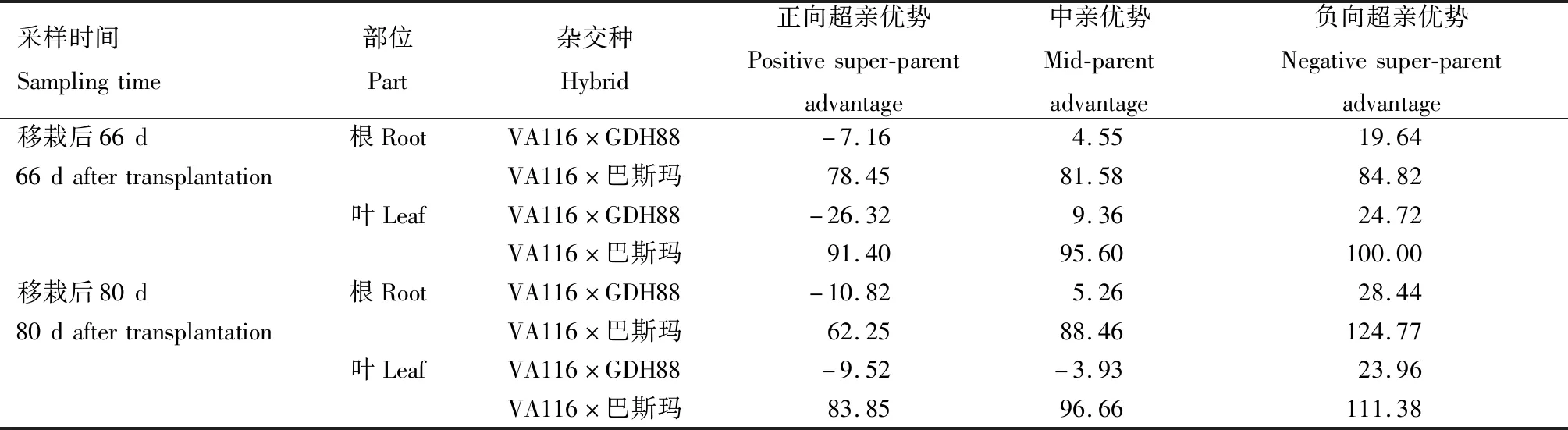
表2 不同时期烟草杂交种烟碱含量的杂种优势表现Tab.2 Heterosis of nicotine content in tobacco hybrid lines at different stages %
2.2 烟草NtMYC2b以及烟碱合成途径关键酶基因的表达分析
由图1可见,移栽后80 d烟草各亲本根中NtMYC2b、PMT和ADC的表达量相对于移栽后66 d明显上调,而QPT表达量则明显下调。其中,VA116、GDH88、巴斯玛根中NtMYC2b的表达量分别是移栽后66 d的2.97倍、11.19倍、4.25倍。巴斯玛根中ODC的表达量明显上调,VA116和GDH88根中ODC的表达量略有下调;GDH88和巴斯玛根中MPO的表达量略有上调,VA116根中MPO的表达量略有下调。移栽后80 d烟草各亲本叶片中ODC、PMT、ADC、QPT的表达量相对于移栽后66 d明显上调;VA116、GDH88、巴斯玛叶片中NtMYC2b的表达量分别是移栽后66 d的0.92倍、1.02倍、0.39倍;VA116和GDH88叶片中MPO的表达量较移栽后66 d上调,而巴斯玛叶片中MPO的表达量下调。

A、 B分别表示根、叶
由图2显示,移栽后80 d,杂交种根中NtMYC2b、ODC、PMT、ADC、MPO的表达量均较2个亲本平均表达量明显上调,其中NtMYC2b在弱优势组合VA116×GDH88和强优势组合VA116×巴斯玛根中的表达量分别是亲本平均表达量的18.96倍、69.75倍;QPT在VA116×GDH88根中的表达量较亲本平均表达量略有下调,而在VA116×巴斯玛根中则明显上调。杂交种叶片中NtMYC2b与PMT、ADC的表达量较2个亲本平均表达量也明显上调,其中NtMYC2b在弱优势组合VA116×GDH88和强优势组合VA116×巴斯玛叶中的表达量分别是亲本平均表达量的1.41倍、73.75倍;ODC、MPO和QPT的相对表达量在VA116×GDH88中分别表现为下调、明显上调和下调,而在VA116×巴斯玛中的表现则相反,表现为明显上调、下调和明显上调。
RT-qPCR结果表明,NtMYC2b和5个关键酶基因在3个亲本和2个杂交种的根部和叶片中均有表达。图1A和图2A表明,NtMYC2b在烟草杂交种和亲本根中的相对表达量均高于5个关键酶基因,表达量较移栽后66 d上调,较亲本平均值也上调。此外,由图2可知,无论在根或叶中,除MPO外,NtMYC2b与其他4个关键酶基因在强优势组合VA116×巴斯玛中的相对表达量都明显高于弱优势组合VA116×GDH88,且在VA116×巴斯玛根与叶中的相对表达量较亲本平均表达量均表现出明显上调,而在VA116×GDH88中QPT下调且其他基因上调也不明显。结合烟碱含量分析,NtMYC2b可能是烟碱合成的调控因子,其表达在烟碱含量杂种优势的形成上扮演着一定角色。
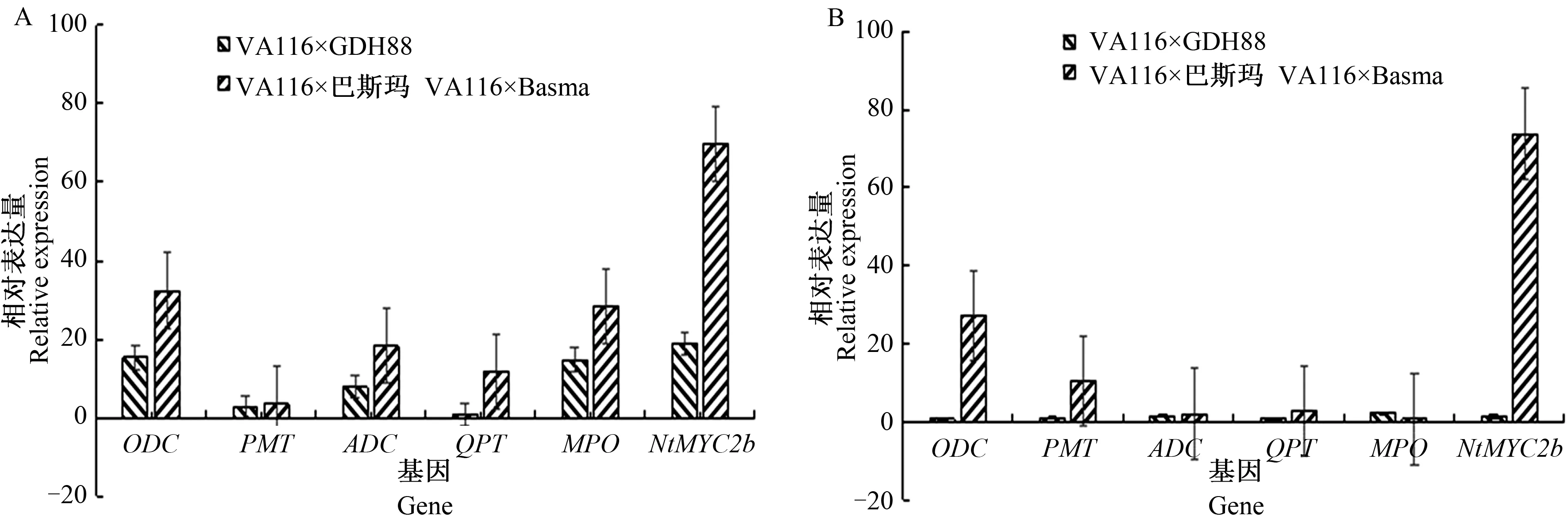
A、 B分别表示根、叶
2.3 NtMYC2b、关键酶基因表达量与烟碱含量及杂种优势的相关分析
表3显示,在亲本根部,NtMYC2b表达量与烟碱含量呈正相关(P>0.05),而与PMT表达量呈显著正相关,相关系数为0.88;此外,ADC表达量与烟碱含量呈显著正相关,相关系数为0.94。在杂交种根部(表4),NtMYC2b表达量与烟碱含量及其杂种优势呈正相关(P>0.05),但与ADC、MPO表达量呈显著正相关,相关系数分别为0.96、0.97;此外,ADC表达量与MPO表达量呈显著正相关,相关系数为0.98。
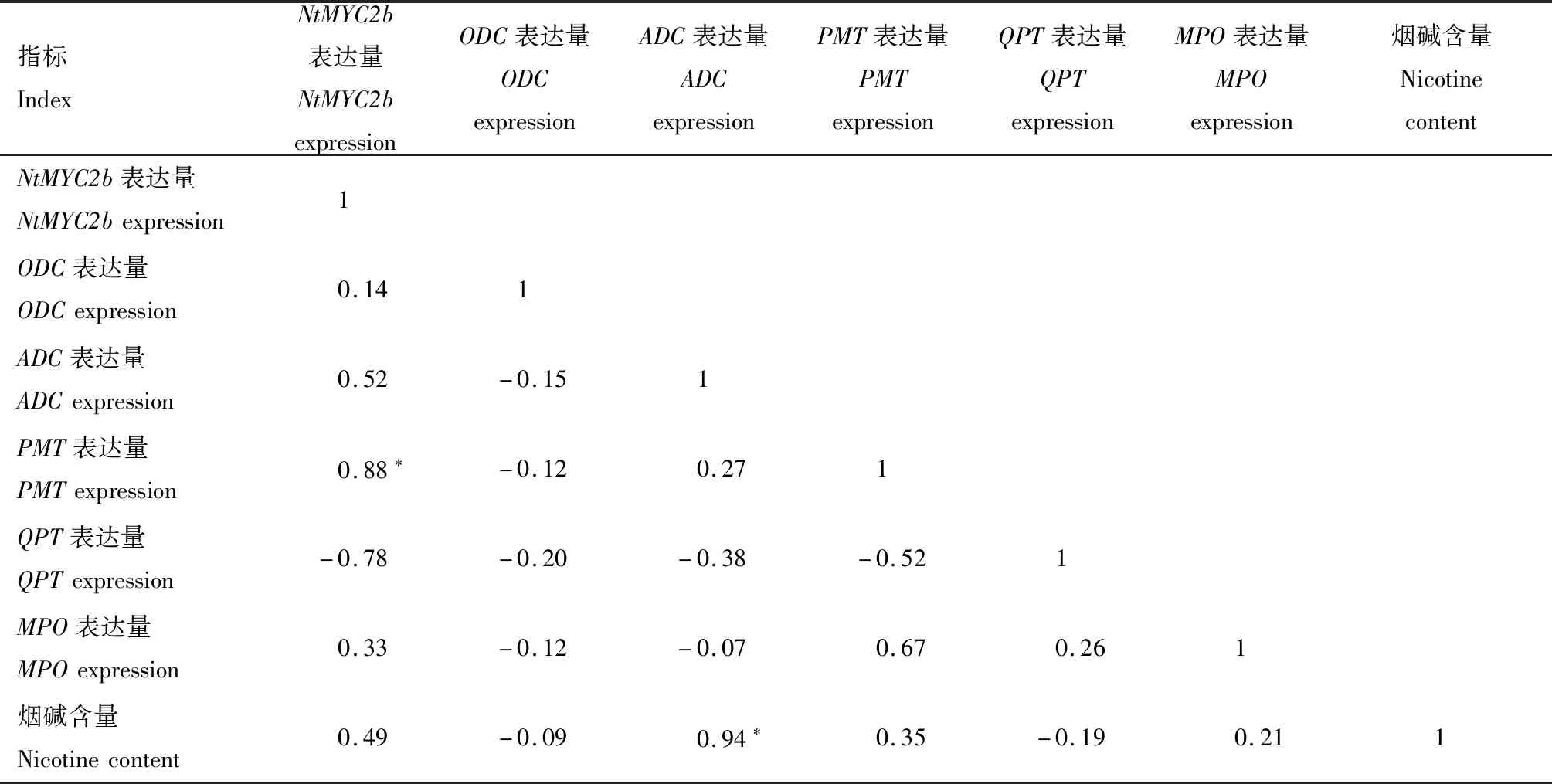
表3 烟草亲本根中NtMYC2b、关键酶基因表达量与烟碱含量间的相关系数Tab.3 Correlation coefficient between expression of NtMYC2b,key enzyme gene and nicotine content in tobacco parental roots
注:*表示显著相关(P<0.05)。
Note:*indicated significant correlation(P<0.05).
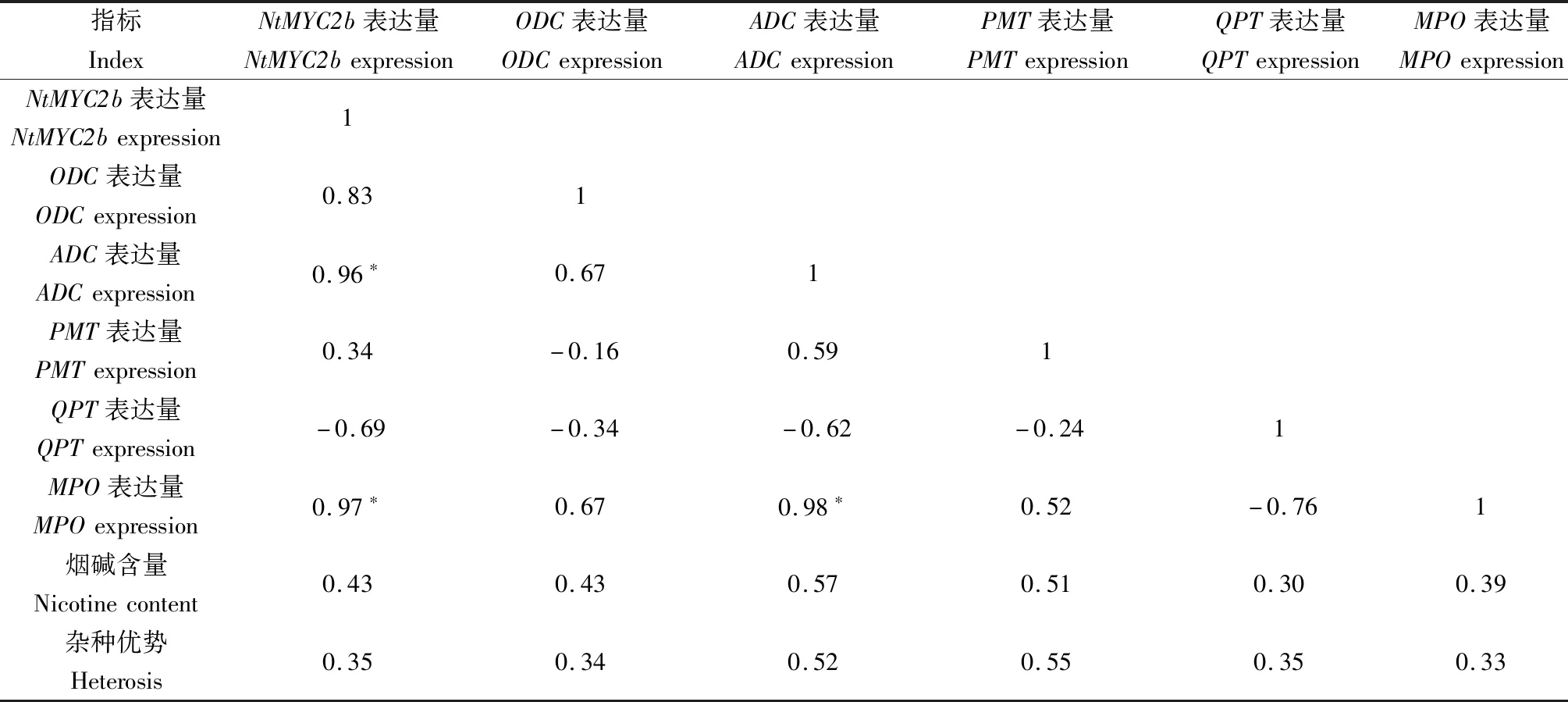
表4 烟草杂交种根中NtMYC2b、关键酶基因表达量与烟碱含量间的相关系数Tab.4 Correlation coefficient between expression of NtMYC2b,key enzyme gene and nicotine content in tobacco hybrid roots
在杂交种叶片(表5)中,NtMYC2b表达量与杂种优势显著正相关,相关系数为0.97;PMT表达量与ODC、ADC表达量呈显著正相关,相关系数均为0.98。可见,PMT、ADC、MPO3个关键酶基因在烟碱合成过程中可能受到NtMYC2b的调控,且关键酶基因间也可能存在互作关系;杂交种叶片中NtMYC2b表达量与烟碱含量杂种优势关系密切。
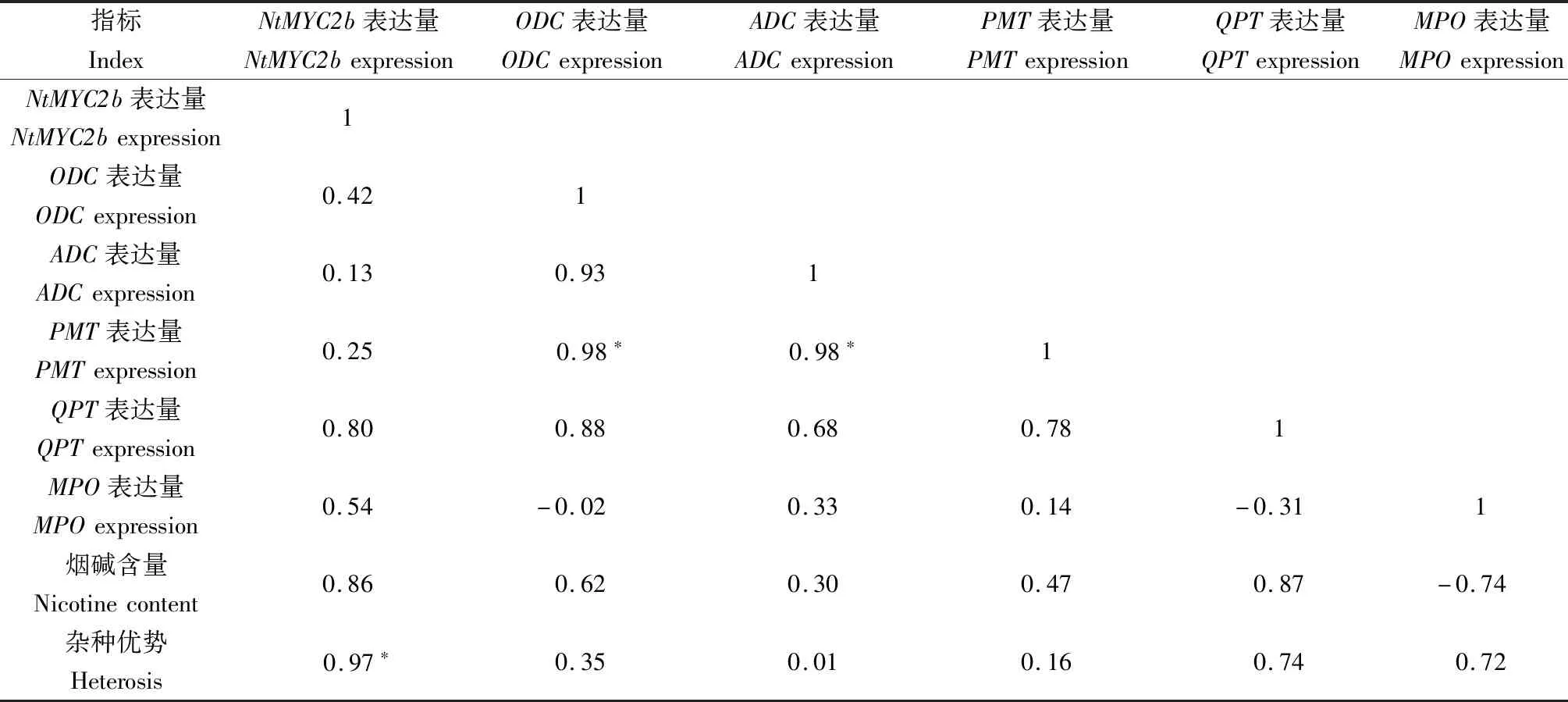
表5 烟草杂交种叶中NtMYC2b、关键酶基因表达量与烟碱含量间的相关系数Tab.5 Correlation coefficient between expression of NtMYC2b,key enzyme gene and nicotine content in tobacco hybrid leaves
2.4 烟碱合成关键酶基因启动子序列中MYC结合位点分析
G-box被证明是与MYC2转录因子结合的核心位点[9,27-28]。在NCBI数据库中查找ADC、PMT的启动子序列,利用Plant CARE在线软件分析关键酶基因ADC、PMT的生物学功能,结果见图3。由图3可见,ADC与PMT的启动子序列中都含有MYC结合位点和G-box区域。其中,在ADC启动子反义链700 bp处含有1个MYC结合位点(CATTTG),正义链1 957 bp处有1个G-box元件(CACGTT)。PMT启动子中含有7个MYC结合位点,分别有3个CATTTG、3个CATGTG、1个CAATTG,其中,在正义链824 bp处有1个CATTTG,在反义链521 bp和986 bp处有2个CATTTG,在正义链1 015 bp和1 985 bp处有2个CATGTG,在反义链1 566 bp处有1个CATGTG,在正义链544 bp处有1个CAATTG;反义链1 991 bp处有1个G-box元件(TACGTG)。由此推测,NtMYC2b可能与ADC、PMT的顺式元件MYC结合位点或G-box元件特异性结合调控ADC、PMT的表达,进而促进烟碱的合成。

A、B分别表示ADC、PMT的启动子序列;+、- 分别表示正义链和反义链;方框表示MYC结合位点;椭圆框表示G-box元件A and B represent promoter sequences of ADC and PMT;+ and - represent justice chain and antisense chain,respectively;The box represents MYC binding site,and the oval box represents G-box element
3 结论与讨论
MYC类转录因子具有多种调节功能,如MYC2调控着植物根[29]、花[30]、果实[31]、种子[32]及叶脉[33]的变化以及烟碱合成[9]等。MYC2转录因子得以发挥调控作用,主要是通过结合结构基因的核心位点G-box区域[9,27-28]。ZHANG等[9]研究表明,转录因子基因NtMYC2a和NtMYC2b在体外也能特异性结合PMT基因启动子的G-box区域,调控PMT基因的表达。本研究发现,ADC、PMT启动子序列中有MYC结合位点及G-box区域,且在亲本根部和杂交种根部,NtMYC2b表达量分别与PMT、ADC表达量显著相关,其表达也与烟碱含量呈正相关,表明NtMYC2b可能与ADC、PMT启动子序列中MYC结合位点或G-box区域特异性结合来调控该基因的表达,从而参与调控烟碱的合成。
烟碱主要在根部合成,经过木质部运输到地上部,存储在液泡中。本研究发现,移栽后80 d,NtMYC2b在烤烟亲本根中的表达量相对于移栽后66 d明显上调,在杂交种根中相对于亲本平均表达量也明显上调,ADC、PMT表达量也明显上调,GDH88和巴斯玛根部烟碱含量增加,但NtMYC2b在亲本叶片中的相对表达量低于根中的相对表达量,而叶片中的烟碱含量高于根中烟碱含量,这可能是因为烟碱在根部合成后向地上部运输,在叶中积累的结果[34]。
杂种优势是指杂交种在生长势、产量、抗性等性状方面优于双亲的遗传现象,在生物界普遍存在[35-37],受多基因控制,遗传机制复杂。目前,主要有显性[36]、超显性[37]和上位性假说[38]来解释杂种优势的形成机制。在烟碱合成代谢的2种途径中,QPT参与吡啶环[39]的形成,ADC、ODC[40]、PMT[41]和MPO[42]参与吡咯环的合成。本研究发现,NtMYC2b与ODC、ADC、PMT、QPT在烟碱含量杂种优势较强的组合中表达量都明显上调,而在弱优势组合中QPT下调且其他基因表达量上调也不明显,说明NtMYC2b参与了烟碱含量杂种优势的形成,且在杂交种中表现为超显性表达。TIAN等[43]对杂交种(VA116×Basma)进行转录组分析,发现烟碱合成代谢相关基因(ADC、PMT、MPO、QPT、AO、QS、QPT、A622、BBLs)和烟碱转运相关基因(JAT2、MATE1、MATE2、NUP1)在杂交种中上调表达,其中大部分被证明为超显性表达,且烟碱合成代谢在杂交种中受到诱导。
本研究探讨了NtMYC2b与烟草烟碱含量及其杂种优势的关系,结果显示,移栽后80 d,NtMYC2b在烟草亲本和杂交种根中的表达量分别较移栽后66 d和亲本平均表达量呈明显上调趋势,且其根和叶中烟碱含量较移栽后66 d增加。NtMYC2b在强优势组合VA116×巴斯玛中的相对表达量均明显高于弱优势组合VA116×GDH88,根和叶中烟碱含量及其杂种优势表现也具有相同的趋势。另外,NtMYC2b表达量与ADC、PMT、MPO表达量及烟碱含量杂种优势呈显著正相关,ADC与PMT基因中都含有MYC结合位点和G-box元件。因此,推测NtMYC2b可能通过与ADC、PMT的顺式元件MYC结合位点或G-box元件特异性结合来调控ADC、PMT的表达,进而促进烟碱的合成,并且NtMYC2b基因的超显性表达参与了烟碱含量杂种优势的形成。
