咖啡酸对铜绿假单胞菌群体感应的抑制及毒力因子降低的研究
2019-09-13于福浩娄在祥王洪新寇兴然
于福浩 , 娄在祥 *, 王洪新 , 寇兴然
(1.江南大学 食品学院,江苏 无锡 214122;2.江南大学 国家功能食品工程技术研究中心,江苏 无锡 214122)
铜绿假单胞菌是临床上一种常见的条件致病菌,它能引起因生物膜和群体感应介导的毒力因子而产生的各种感染,被认为是医院中的第三大致病菌[1]。铜绿假单胞菌也是食物中的普遍致病菌,经常引起食物腐败和食源性疾病。因为抗生素的过度使用,许多铜绿假单胞菌对抗生素已经形成了很高的抗药性。因此,采用非抗生素、安全的天然化合物取代传统的药物,对控制致病菌来说是必要的。
群体感应是一种细胞密度依赖的基因表达现象。细菌依赖于扩散性的信号分子监测周围同种或异种细菌的密度,当信号分子达到一定浓度阈值时可调控特定基因的表达。革兰氏阴性菌的群体感应系统中主要有合成酶蛋白和受体蛋白参与,合成酶蛋白分泌N-酰基高丝氨酸内酯(AHL)作为自体诱导物,当这种自体诱导物密度积累到阈值时就与转录受体蛋白结合,启动相关基因的转录[2]。作为一个重要的调节系统,群体感应系统调控着一系列细菌的行为,如生物膜形成,毒力因子分泌,免疫逃逸和抗药性的产生[3-5]。研究发现,铜绿假单胞菌的生物膜形成和毒力因子分泌都是由群体感应系统调控的[6],因此,以群体感应为靶点研发群体感应抑制剂已成为一种新的趋势[7-8]。这种新型群体感应抑制剂不会抑制细菌的生长,在降低细菌致病力的同时不使其产生抗药性,这对细菌耐药性的问题具有极重要的意义[9-11]。
绝大多数传统的群体感应抑制剂(如卤代呋喃酮等)被发现有毒害作用[12-13]。相反,从可食植物中筛选安全、无毒的活性成分作为群体感应抑制剂来干扰群体感应系统已经引起了越来越多的关注。咖啡酸普遍存在于植物中,主要来自柠檬皮、毛莨科植物升麻属根茎、缬草属植物根等。因此,本实验选用广泛存在于植物中的咖啡酸作为群体感应抑制剂,研究其对铜绿假单胞菌群体感应的抑制作用及抑制毒力因子分泌和生物膜形成的能力。
1 材料与方法
1.1 材料
咖啡酸(≥98%),购于西安小草植物科技有限责任公司;铜绿假单胞菌CMCC(B)10104,购于中国微生物菌种保藏中心 (中国,北京);菌种Choromobacterium violaceum(CV026),购于西班牙典型培养物保藏中心。
1.2 最小抑菌质量浓度MIC的测定
采用二倍稀释法测定咖啡酸对紫色色杆菌和铜绿假单胞菌的MIC。将配置好的LB培养液(牛肉膏 10 g,胰蛋白胨 5 g,氯化钠 10 g,水 1000 mL,调整pH为7.2)分装到各个试管中,每管1 mL。向第1管中加入1 mL药物原液,混匀后取出1 mL依次加到下一管中,最后一管弃掉1 mL,形成咖啡酸一系列浓度,最后向试管中加入10 μL活化好的菌液。阳性对照不加药物,阴性对照组不加菌液。37℃恒温培养24 h,观察细菌生长情况,以抑制细菌生长所含的最低药物质量浓度为最小抑菌质量浓度MIC。
1.3 咖啡酸对细菌生长的影响
将紫色色杆菌于LB培养液中过夜培养,将已活化好的菌种分别接种于新鲜的LB培养液中,加入适量的咖啡酸,使咖啡酸的终质量浓度分别为0.200、0.100、0.050、0.025 mg/mL, 对照组不添加药物。实验组和对照组菌液的起始OD600为0.05,37℃振荡培养24 h,每隔2 h分别取样测OD600。
1.4 对群体感应活性的影响
1.4.1 指示菌平板法定性检测咖啡酸的群体感应活性 咖啡酸对群体感应的抑制效果用群体感应指示菌C.violaceum测定,并做适当的修改[8]。将群体感应指示菌C.violaceum活化后按2%的接种量(体积分数)接种到新鲜LB培养液中,恒温振荡培养至OD600为1.0。LB琼脂固体培养基融化后待冷却至50℃,将对数期的菌液缓缓加入,轻轻混匀后倒平板。凝固后将20 μL浓度为40 μmol/L的信号分子C6-HSL涂布于平板表面,用滤纸片法检测,每个滤纸片(直径6 mm)上分别滴加10 μL不同质量浓度的咖啡酸,37℃培养24 h,观察纸片周围无色圈的直径。
1.4.2 定量检测咖啡酸的群体感应活性 咖啡酸对紫色杆菌素产量的影响根据已报道的方法测定,并做适当的修改[14-15]。将100 μL处于生长对数期的C.violaceum接种到4 mL含有信号分子C6-HSL的新鲜LB培养液中,同时添加不同质量浓度的咖啡酸,对照组不添加咖啡酸。37℃恒温振荡培养24 h后,取1 mL于离心管中,13000 r/min离心10 min,弃掉上清液,沉淀加入500 μL DMSO使紫色杆菌素溶解出来,再在13000 r/min的转速下离心10 min,上清液中则含有紫色杆菌素,取上清液测其OD585。
1.5 对群体感应调控的生物膜的影响
1.5.1 结晶紫染色法测定咖啡酸对生物膜抑制作用的影响 将铜绿假单胞菌于LB培养液中过夜培养。取50 μL菌液接于含1 mL LB培养液的24孔板中,同时加入不同浓度的咖啡酸,使其终质量浓度为 0.4,0.2,0.1,0.05 mg/mL, 对照组不添加药物,37℃恒温静置培养36 h。培养完吸出孔中液体,每孔加入1 mL的无菌PBS缓冲液清洗板孔3次,待干燥后每孔加1 mL 1 g/dL的结晶紫染色5 min,吸出结晶紫染液,用流水洗掉多余染料,静置干燥完全,之后每孔加1 mL 95%的乙醇,37℃烘箱中作用30 min以溶解结晶紫,570 nm 处测其吸光值[5,16]。
1.5.2 激光共聚焦显微镜观察生物膜 将铜绿假单胞菌于LB培养液中过夜培养。按取500 μL菌液接入10 mL新鲜LB培养液中,加入不同质量浓度的咖啡酸,使其终质量浓度为0.40、0.20、0.10、0.05 mg/mL,同时将灭过菌的盖玻片作为粘附载体放入培养皿中,对照组不添加药物,37℃恒温静置培养36 h。培养结束后将盖玻片从培养皿中取出,用去离子水轻轻洗涤,除去表面的游离细菌,将盖玻片放入已配好的染色剂(SYTO9和PI)中避光染色30 min,后置于激光共聚焦显微镜下观察生物膜的形成情况[17]。
1.6 对群体感应调控的毒力因子产量的影响
将铜绿假单胞菌于LB培养液中过夜培养。将活化好的菌种接于新鲜LB培养液中,同时加入不同质量浓度的咖啡酸,使其终质量浓度为0.40、0.20、0.10、0.05 mg/mL,对照组不添加药物(只含培养基和菌液),37℃振荡培养24 h。培养后的菌液经不同处理测其对绿脓菌素和鼠李糖产量的影响。
1.6.1 对绿脓菌素产量的影响 培养后的菌液经离心,取5 mL上清液于15 mL离心管中,加入3 mL氯仿充分振荡以使两者充分混匀,静置后将氯仿层转入干净的EP管中,加入1 mL 0.2 mol/L的盐酸充分振摇,静置分层后取上层含有绿脓菌素的粉红色有机相测其OD520[18]。
1.6.2 对鼠李糖脂产量的影响 首先制作出鼠李糖标准曲线。配制鼠李糖标准品的梯度质量浓度溶液:0、50、100、200、300、400、500 μg/mL。 取 1 mL 鼠李糖标准品加入到已配好的0.19 g/dL的苔黑酚—浓硫酸溶液中,定容至10 mL。80℃下水浴半小时后取出置室温保持15 min,测其在421 nm处的OD值,并制作鼠李糖标准曲线,1 mg鼠李糖相当于2.5 mg鼠李糖脂。
培养后的菌液经6000 r/min离心10 min,除去菌体,过滤后取上清液用浓盐酸调至pH为2,取1 mL加入3倍体积的乙酸乙酯萃取2次,将乙酸乙酯层混匀,旋转蒸发至干,最后加入1 mL的去离子水溶解。取1 mL按上述步骤测其在421 nm处的吸光值[19-21]。
1.6.3 对游动能力的影响 游动能力的测定采用的是双层平板法[22-23]。在平板底层倒入含质量分数2%琼脂的LB琼脂培养基,上层倒入含咖啡酸的质量分数0.3%琼脂的软琼脂培养基,咖啡酸的终质量浓度为 0.40、0.20、0.10、0.05 mg/mL,对照组不添加药物。在平板的中央接种2 μL的已活化好的菌液,37℃恒温培养24 h,观察致病菌形成的浮游圈大小。
1.7 对信号分子的抑制作用
将铜绿假单胞菌于LB培养液中过夜培养。将活化好的菌液加到6 mL新鲜LB液体培养基中,同时加入不同质量浓度的咖啡酸,使其终质量浓度为0.40、0.20、0.10、0.05 mg/mL,对照组不添加药物(只含培养基和菌液),37℃振荡培养24 h。培养后将培养液6000 r/min离心10 min,取上清液用等量含0.02 mol/L冰乙酸的乙酸乙酯萃取3次,合并有机相。将上述得到的乙酸乙酯提取液在旋蒸仪内旋转蒸干,后用体积分数50%的甲醇水溶液将其溶解,经0.22 μm膜过滤,采用超高效液相-串联质谱定性定量分析群体感应信号分子[24]。
2 结果与分析
2.1 最小抑菌浓度的测定结果
通过试管中细菌的生长情况判断药物对细菌的最小抑菌浓度(MIC)。观察阳性对照组明显浑浊,阴性对照组澄清,所以,未见试管浑浊的最低药物质量浓度即为最小抑菌质量浓度。如表1所示,咖啡酸对紫色色杆菌的最小抑菌质量浓度为0.4 mg/mL,对铜绿假单胞菌的最小抑菌质量浓度为0.8 mg/mL。
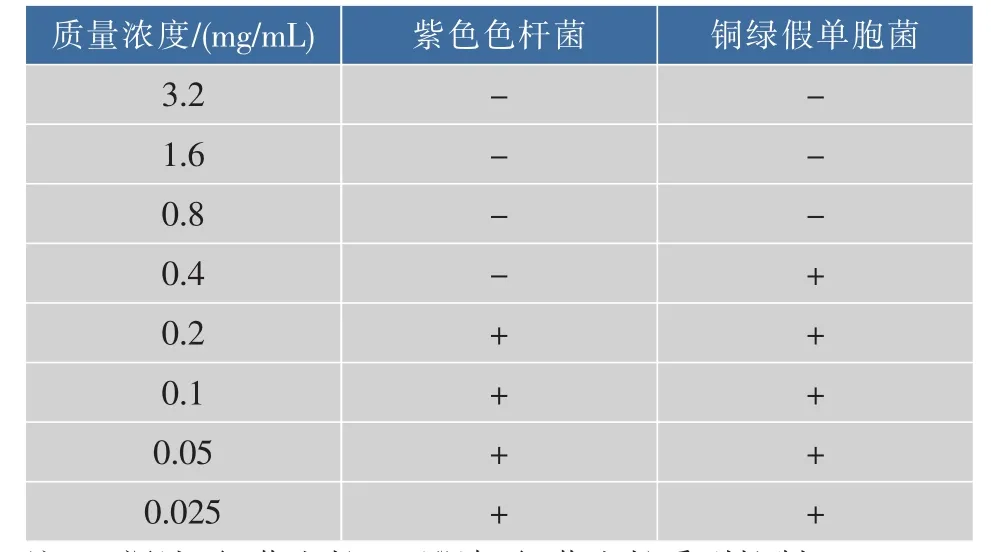
表1 咖啡酸对细菌的MICTable 1 MIC of caffeic acid on bacteria
2.2 咖啡酸对细菌生长的影响
为了研究咖啡酸对群体感应的抑制作用及对其调控的毒力因子的产生量的影响,实验应在亚抑菌质量浓度下进行。咖啡酸对紫色色杆菌的MIC为0.4 mg/mL,因此实验采用最小抑菌质量浓度以下的4个质量浓度梯度,验证在此质量浓度下对紫色色杆菌的生长是否会产生影响,实验结果如图1。从图中结果可以看出,在亚抑菌质量浓度下,咖啡酸对紫色色杆菌的生长几乎无明显影响。因此,接下来的实验可在亚抑菌质量浓度下进行。
2.3 咖啡酸的群体感应活性结果
群体感应的定性实验中,滤纸片周围无色圈的大小代表了咖啡酸抑制群体感应能力的强弱。从图2可以看出,含咖啡酸的滤纸片周围的紫色素的产量明显低于不加药物的滤纸片,且不影响细菌的生长。当质量浓度为0.2 mg/mL时,咖啡酸滤纸片周围无色圈的大小达到了1.32 cm,表明咖啡酸有抑制紫色色杆菌群体感应的能力。

图1 紫色色杆菌的生长曲线Fig.1 Growth curves of C.violaceum

图2 咖啡酸对紫色色杆菌群体感应的定性实验Fig.2 Qualitative QS inhibition assay of caffeic acid
由于紫色色杆菌其紫色素的产生是其群体感应系统调控的,因此检测紫色杆菌素的产量可以确定药物对细菌群体感应的作用。群体感应的定量实验结果如图3,表明咖啡酸明显的抑制了紫色杆菌其紫色杆菌素的产量,抑制能力呈质量浓度依赖性。随着质量浓度的增大,咖啡酸对紫色杆菌素的抑制率从17.9%增加到61.1%,在质量浓度为0.2 mg/mL时,抑制率最大为61.1%。

图3 定量检测咖啡酸对紫色色杆菌群体感应的抑制作用Fig.3 Inhibition effectofcaffeic acid on violaceinproduction in C.violaceum by quantitative assay
2.4 咖啡酸对生物膜的抑制作用结果
细菌生物膜对其产生抗药性具有重要作用。经培养后,24孔板内壁上形成了一圈薄膜,用结晶紫染色后可通过吸光值的大小反映生物膜的形成量。如图4所示,在亚抑菌质量浓度下,随着咖啡酸质量浓度的增加,生物膜的形成量显著减少。且在质量浓度为0.4 mg/mL即1/2 MIC时,对生物膜的抑制率达到最大,为65.3%。说明咖啡酸能不同程度的抑制铜绿假单胞菌生物膜的形成。

图4 咖啡酸对铜绿假单胞菌生物膜的影响Fig.4 Effect of caffeic acid on biofilm in P.aeruginosa
用激光共聚焦观察铜绿假单胞菌生物膜的形成情况。对比不同质量浓度的咖啡酸对生物膜形成的影响,结果如图5所示。加入咖啡酸后铜绿假单胞菌形成生物膜的能力显著降低,且随着咖啡酸质量浓度的增加,抑制效果明显增强。在不添加药物的对照组中,铜绿假单胞菌形成的生物膜很致密,颜色很亮,在添加质量浓度为0.1 mg/mL咖啡酸的实验组中,菌体密度变得较疏松,亮度也减弱,在添加质量浓度为0.4 mg/mL咖啡酸的实验组中,效果更明显。
铜绿假单胞菌的生物膜无处不在,它是细菌产生抗药性的最重要的原因,生物膜是高度结构化的片状复合物,绝大多数细菌在适宜的条件下都能形成生物膜。生物膜的形成显著增加了膜内细菌对抗生素的耐药性,这对人体健康造成了严重的伤害。Sandasi等[25]报道出药草、香料和重要药用植物中提取物能减少初始细胞粘附。Zeng等[26]研究表明黄芩素,另一个天然黄酮醇,是一种能对铜绿假单胞菌生物膜产生剂量依赖性抑制作用的群体感应抑制剂。本实验也得出咖啡酸能不同程度的抑制铜绿假单胞菌生物膜的形成,对抑制细菌耐药性的产生提供了一定的理论依据。

图5 激光共聚焦观察咖啡酸对铜绿假单胞菌生物膜的抑制作用Fig.5 Confocal Laser Scanning Microscopy (CLSM)analysis:the inhibition of caffeic acid on biofilm formation of P.aeruginosa
2.5 咖啡酸对毒力因子的抑制作用结果
2.5.1 对绿脓菌素的抑制结果 绿脓菌素是铜绿假单胞菌的重要毒力因子之一,反映菌株的毒力大小,它涉及到正常铁离子的转运及多种细胞的功能,过量的绿脓菌素会导致细胞死亡。咖啡酸对铜绿假单胞菌的绿脓菌素产量的抑制结果如图6所示。在亚抑菌质量浓度下,随着咖啡酸质量浓度的增加,绿脓菌素的产量明显减少。咖啡酸在质量浓度为0.05 mg/mL即1/16 MIC时,对绿脓菌素的抑制率达到了14.7%;在质量浓度为0.4 mg/mL即1/2 MIC时,对绿脓菌素的抑制率高达66.4%,而对铜绿假单胞菌的生长无影响。说明咖啡酸抑制了群体感应调控的绿脓菌素的产生。

图6 咖啡酸对铜绿假单胞菌绿脓菌素的影响Fig.6 Effect of caffeic acid on pyocyanin in P.aeruginosa
2.5.2 对鼠李糖脂的抑制结果 鼠李糖脂作为生物表面活性剂,可以增强细菌间的交流进而增强细菌的致病力,是铜绿假单胞菌群体感应调控的另一毒力因子。经咖啡酸作用的铜绿假单胞菌其鼠李糖脂含量减少。如图7所示,当咖啡酸质量浓度为0.4 mg/mL时,鼠李糖脂的抑制率可达31.8%。表明咖啡酸对鼠李糖脂起到了一定的抑制作用。

图7 咖啡酸对铜绿假单胞菌鼠李糖脂的影响Fig.7 Effect of caffeic acid on rhamnolipid in P.aeruginosa
2.5.3 对游动能力的抑制结果 由鞭毛调节的细菌游动能力和生物膜的形成紧紧相关,生物膜的黏附过程是通过鞭毛运动开始的。由图8可以看出,未加药物的对照组,铜绿假单胞菌的游动能力良好,在加入咖啡酸的实验组中,细菌不能充分的扩散开,围绕接种位置形成菌圈,且随着质量浓度的增加,对游动能力的抑制作用增强。药物质量浓度均小于MIC,因此咖啡酸对细菌的生长无抑制,表明咖啡酸对细菌鞭毛的抑制导致了游动能力的减弱。
2.6 对信号分子的抑制
用HPLC-MS对铜绿假单胞菌的信号分子进行分析。在亚抑菌质量浓度下,即咖啡酸质量浓度低于0.8 mg/mL时,铜绿假单胞菌的生长几乎不受影响。因此,培养后经HPLC-MS检测发现,咖啡酸处理过的铜绿假单胞菌其信号分子的量与对照组相比都有不同程度的降低。且随着咖啡酸质量浓度的增加,信号分子的产量也降低。如图9所示,当咖啡酸质量浓度为0.4 mg/mL时,对C4-HSL,C6-HSL,C8-HSL和3-oxo-C12-HSL的抑制率分别为59.6%,49.2%,53.9%,52.2%。当咖啡酸在质量浓度为0.05 mg/mL时,对C4-HSL的抑制率也达到了22.7%。C4-HSL信号分子调控着绿脓菌素和鼠李糖脂这2个毒力因子的产生,由于信号分子降低趋势同毒力因子的降低趋势一致,且信号分子是由信号分子合成酶合成,因此可推测药物通过抑制信号分子合成酶的分泌来抑制细菌毒力因子的产生,从而降低细菌毒力[27]。
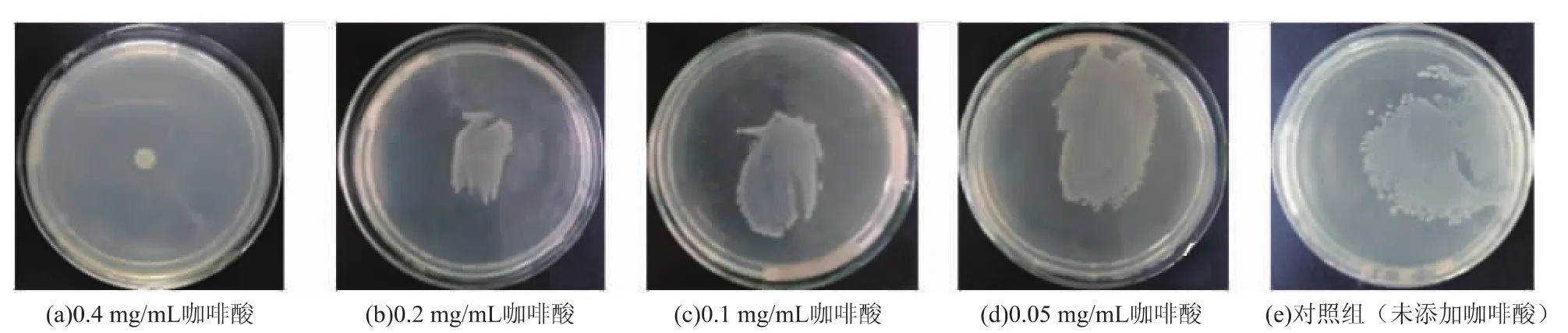
图8 咖啡酸对铜绿假单胞菌游动能力的抑制作用Fig.8 Inhibition effect of caffeic acid on swarming motility of P.aeruginosa
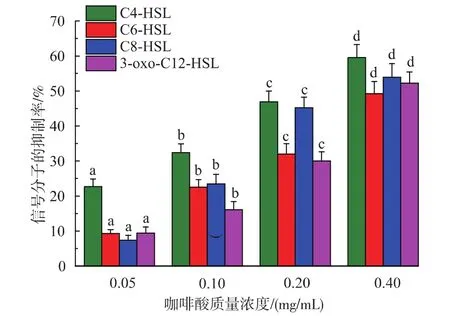
图9 咖啡酸对铜绿假单胞菌信号分子的抑制作用Fig.9 Effect of caffeic acid on AHLs production in P.aeruginosa
3 结 语
研究了咖啡酸对铜绿假单胞菌群体感应及其调控的毒力因子和生物膜的抑制作用,发现其能显著抑制生物膜的形成,对毒力因子包括绿脓菌素、鼠李糖脂、游动能力等有显著的作用,也明显地抑制了群体感应信号分子的产量,推测其抑制机制可能是咖啡酸通过抑制信号分子合成酶的活性,进而抑制信号分子的合成。咖啡酸主要来源于植物中,且对群体感应具有良好的抑制效果。因此,咖啡酸作为一种安全、无毒害具有抑制群体感应潜能的天然化合物,在降低细菌致病力及细菌抗药性方面有很强的应用价值。
