白桦脂醇对AGEs诱导H9C2心肌细胞自噬的影响①
2019-08-13杜娟娟
罗 萍 杜娟娟 杜 松
(河南省人民医院心内科,郑州450003)
细胞自噬是一种存在于真核生物中高度保守的代谢过程[1]。通过清除体内异常的蛋白质和受损细胞器,参与细胞物质代谢平衡的调节[2]。细胞自噬在机体炎症、免疫、肿瘤、神经退行性疾病和心血管疾病的发生发展过程中发挥着重要的作用[3]。心血管疾病是引起人类死亡的最主要原因,患病的风险随着年龄增长而增加。据统计2012年有1 750万人死于心血管疾病,占全球死亡人数的30%以上[4]。越来越多的证据表明,在心血管系统中,正常的自噬对维持血管和心脏的稳态和功能至关重要,然而自噬缺陷或过度自噬会造成心血管疾病的发生,如心力衰竭、心肌肥大和动脉粥样硬化[5]。晚期糖基化终产物(Advanced glycation end products,AGEs)是糖尿病血管并发症的一大特征[6]。AGEs及其受体相互作用,在体内诱发氧化应激,促进炎性反应,加速心血管疾病的发生[7]。大量研究发现,AGEs通过损伤组织细胞,能显著提高细胞自噬水平[8]。白桦脂醇是来源于白桦树皮的一种羽扇烷型三萜类化合物。近年来,白桦脂醇在抗炎、抑菌、抗肿瘤和抗HIV等方面效果明显,因而广受关注[9,10]。近期有研究发现,白桦脂醇对受损的心肌具有保护作用[11],但是白桦脂醇对心肌细胞自噬的影响还未见相关报道。本文使用AGEs诱导H9C2心肌细胞自噬,探究白桦脂醇对细胞自噬的影响。
1 材料与方法
1.1材料 白桦脂醇(纯度>98%,cas:473-98-3)购自道斯夫生物;AGE-BSA(2221-10)购自艾美捷科技有限公司;巴佛洛霉素A(Bafilomycin,Baf-A1)购自美国Selleck公司;DMEM培养基、胎牛血清(FBS)和胰酶购自Hyclone公司;盘尼西林(Penicillin)和链霉素(Streptomycin)购自Thermo Fisher Scientific;CCK8试剂盒购自默沙克生物;肌红蛋白(Myoglobin,Mb)、肌酸激酶同工酶(Creatine kinase isoenzyme,CK-MB)和心肌肌钙蛋白(Myocardial troponin,cTnⅠ)ELISA检测试剂盒购自罗氏;BCA试剂盒购自北京天根生物;Beclin1、P62、LC3Ⅱ、LC3Ⅰ、PI3K、p-PI3K、AKT、p-AKT兔抗大鼠一抗和羊抗兔二抗及显影液购自美国Abcam;H9C2细胞购自美国ATCC公司。
1.2方法
1.2.1细胞的培养及分组 H9C2细胞复苏后,加入DMEM+10%FBS+100 U/ml盘尼西林+100 μg/ml链霉素的培养基,于37℃、5%CO2的生化培养箱中培养。细胞随机分成正常H9C2组,AGEs组(100 mg/L),低剂量加药组(5 μmol/L),中剂量加药组(10 μmol/L),高剂量加药组(20 μmol/L)。
1.2.2CCK8检测细胞活性 取对数生长期的H9C2心肌细胞,消化后,离心弃上清,用培养基将细胞悬液的浓度调整为5×104个/ml,96孔板中加100 μl 细胞悬液。同时加入不同相应浓度白桦脂醇和(或)AGEs,空白组使用不加药物的溶液。培养箱中贴壁生长24 h,加入10 μl CCK8。将培养板在培养箱中孵育2 h。用酶标仪测定450 nm处的吸光度。实验重复3次。
1.2.3酶联免疫吸附实验 收集H9C2细胞,计数后,用预冷的PBS洗3次,加细胞裂解液,超声后破碎,12 000 r/min离心15 min,收集上清备用。抗体包被反应孔,加稀释后的待检测样品,设置空白组和对照组,置于37℃孵育1 h。加入稀释后的酶标抗体,37℃孵育1 h;加底物于37℃反应15 min后,终止反应。以空白组调零后,酶标仪下检测450 nm处光吸收值,根据标准曲线计算酶的含量。
1.2.4Western blot 取对数生长期的H9C2细胞,稀释后接种于含AGEs和(或)不同浓度的白桦脂醇的培养基中培养48 h后。收集细胞并计数,使用含蛋白酶抑制剂的RIPA细胞裂解液处理H9C2细胞。BCA法测定蛋白浓度,并调整蛋白浓度,制备蛋白样品。等量蛋白样品经12%的SDS-PAGE凝胶分离后转移至PVDF膜上。5%脱脂牛奶室温封闭1 h,一抗4℃孵育过夜,TBST洗3次,每次5 min,室温下孵育二抗1 h,TBST洗膜后加显色液显影。凝胶成像系统中观察结果并拍照,使用Image-pro plus软件分析蛋白条带灰度。
1.2.5免疫荧光染色 在培养板中将已爬好细胞的玻片用PBS洗3次,每次3 min,用4%的多聚甲醛固定15 min,PBS清洗;用PBS配制的0.5% Triton X-100室温下通透20 min。山羊血清室温封闭30 min;除去封闭液,加稀释好的一抗,4℃孵育过夜。PBST清洗,加稀释好的带荧光标签的二抗,室温下孵育1 h。滴加DAPI避光孵育5 min,用PBST清洗除去DAPI,含抗荧光淬灭剂的封片液封片,于荧光显微镜下观察采集图像。
1.3统计学分析 使用SPSS18.0统计软件和GraphPad Prism 5分析数据和作图。数据是3次实验的平均标准偏差(SD)。两组之间的差异用独立的t检验。P<0.05表示差异具有统计学意义。
2 结果
2.1白桦脂醇促进AGEs作用下H9C2心肌细胞的活性 AGEs组细胞增殖倍数明显低于正常H9C2组(图1,P<0.01)。白桦脂醇与AGEs共同作用于H9C2细胞后,细胞的增殖倍数显著升高,且增殖倍数随着白桦脂醇浓度的增加而升高(图1,P<0.01)。实验结果表明,白桦脂醇促进AGEs作用下H9C2心肌细胞的活性。
2.2白桦脂醇对心肌酶表达的影响 ACEs组与H9C2组相比较,细胞中Mb、CK-MB和cTnⅠ的表达显著上升(图2,P<0.01)。将白桦脂醇与ACEs共同作用于H9C2细胞后,细胞中Mb、CK-MB和cTnⅠ的表达明显下降;且白桦脂醇的浓度越高,降低的效果越明显(图2,P<0.01)。实验结果表明,白桦脂醇抑制AGEs诱导的H9C2心肌细胞损伤。
2.3白桦脂醇对自噬相关蛋白表达的影响 正常H9C2经AGEs处理后,Western blot检测蛋白表达,发现Beclin1的表达显著上调,P62的表达显著下调,LC3Ⅱ的表达上调,使LC3 Ⅱ/LC3 Ⅰ的比率显著上升,提示细胞自噬水平升高(图3A,P<0.01)。白桦脂醇与AGEs共同作用于H9C2细胞后,Beclin1表达显著下调,P62表达显著上调,而LC3Ⅱ的下调和LC3Ⅰ的上调,使LC3Ⅱ/LC3Ⅰ显著下降(图3A,P<0.01)。向上述各组细胞中加入巴佛洛霉素A1,检测结果显示,AGEs组与H9C2组相比,LC3 Ⅱ/LC3 Ⅰ显著上升;不同浓度白桦脂醇处理细胞后,LC3 Ⅱ/LC3 Ⅰ显著降低(图3C,P<0.01),且上述作用均具有白桦脂醇剂量依赖效应。
2.4白桦脂醇抑制AGEs作用下H9C2心肌细胞自噬体的形成 免疫荧光染色结果显示,AGEs处理H9C2心肌细胞后,LC3光点呈现团状,光点数目显著上升,表明有大量自噬体形成(图4,P<0.01)。白桦脂醇与AGEs共同处理H9C2细胞后,LC3光点随着白桦脂醇浓度的增加而显著减少(图4,P<0.01)。实验结果表明,白桦脂醇抑制AGEs作用下H9C2心肌细胞自噬体的形成。
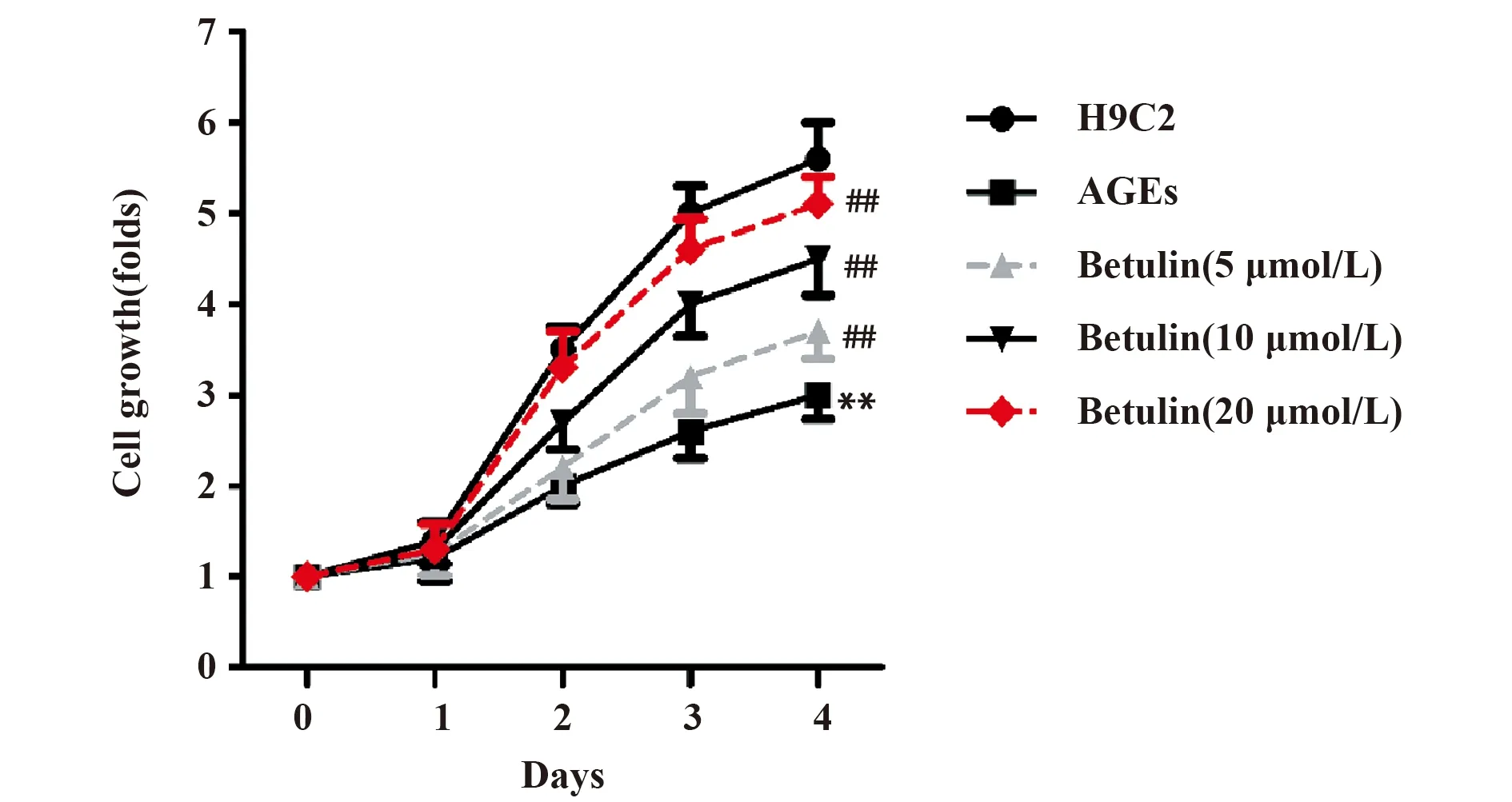
图1 白桦脂醇对心肌细胞细胞活性的影响Fig.1 Effect of Betulin on activity of myocardial cellsNote: Compared with the H9C2 group,**.P<0.01;compared with the AGEs group,##.P<0.01.
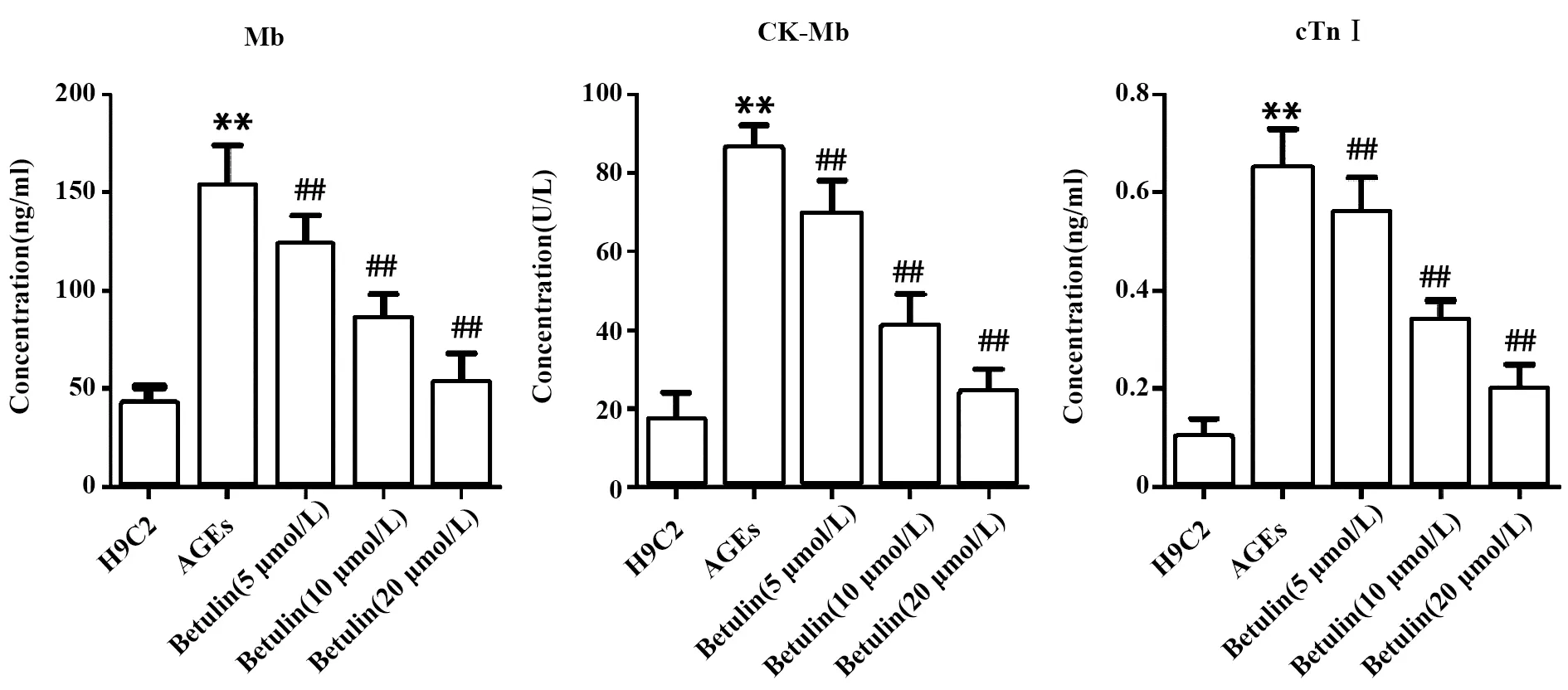
图2 白桦脂醇抑制AGEs对H9C2心肌细胞损伤的诱导作用Fig.2 Betulin inhibits H9C2 myocardial cell injury induced by AGEsNote: Compared with the H9C2 group,**.P<0.01;compared with the AGEs group,##.P<0.01.
2.5白桦脂醇抑制AGEs作用下H9C2细胞PI3K和AKT的活化 AGEs处理H9C2细胞后,PI3K和AKT蛋白总量未见明显变化,p-PI3K 和p-AKT则显著上升,使p-PI3K/PI3K和p-AKT/AKT显著提高(图5,P<0.01)。白桦脂醇与AGEs共同作用于H9C2心肌细胞后,PI3K和AKT蛋白总量没有明显变化,p-PI3K和p-AKT的含量随着白桦脂醇浓度的增加而降低,使p-PI3K和p-AKT在细胞中占比显著下降(图5,P<0.01)。实验结果表明,白桦脂醇降低PI3K和AKT的磷酸化水平,在起始阶段抑制AGEs诱导的H9C2细胞自噬。
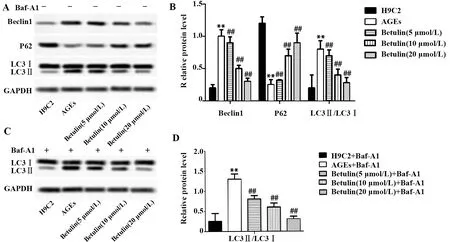
图3 白桦脂醇对自噬相关蛋白表达的影响Fig.3 Effect of Betulin on expression of autophagy related proteinsNote: A and B.Effects of betulin on Beclin1,P62 and LC3Ⅱ/LC3Ⅰ without the addition of Bafilomycin A1;C and D.Eeffects of betulin on LC3Ⅱ/LC3Ⅰ in the presence of Bafilomycin(200 nmol/L).Compared with the H9C2 group,**.P<0.01;compared with the AGEs group,##.P<0.01.


图4 免疫荧光检测LC3的表达Fig.4 Expression of LC3 was determined by immunofluorescenceNote: Compared with the H9C2 group,**.P<0.01;compared with the AGEs group,##.P<0.01.
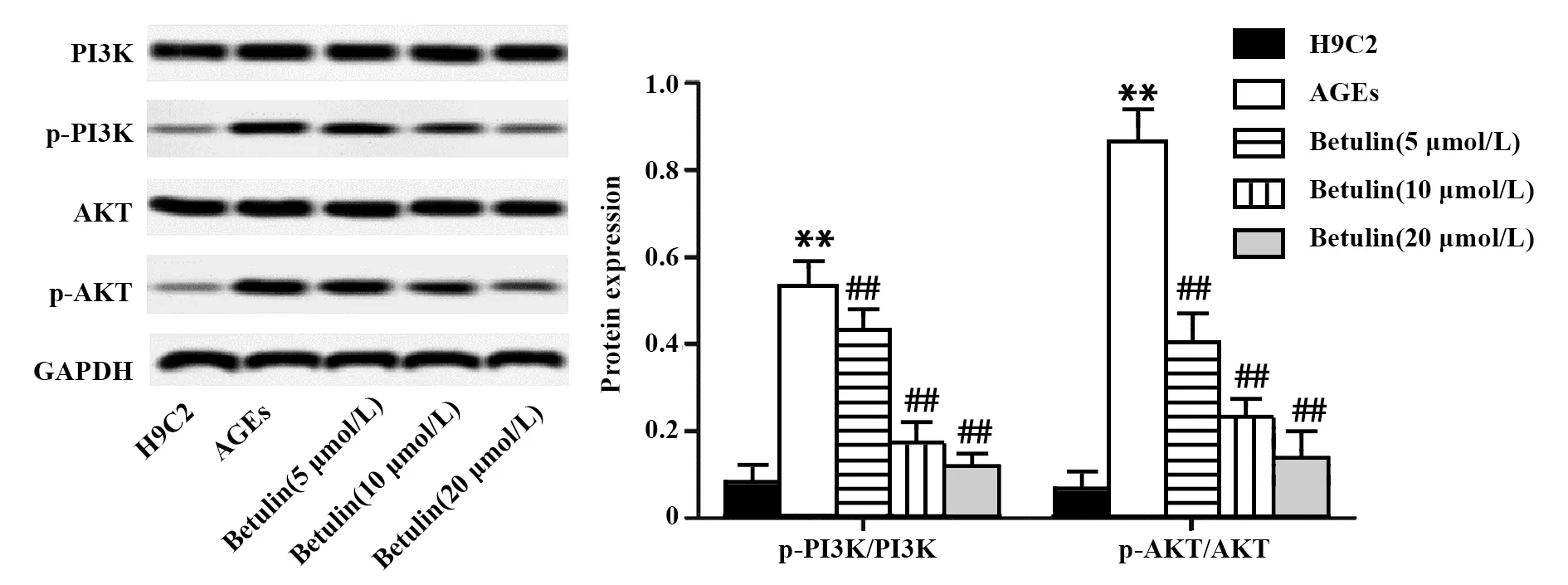
图5 白桦脂醇对AGEs作用下H9C2细胞PI3K,AKT表达及其磷酸化水平的影响Fig.5 Effect of Betulin on expression and phosphorylat-ion level of PI3K and AKT in AGEs treated H9C2 cellsNote: Compared with the H9C2 group,**.P<0.01;compared with the AGEs group,##.P<0.01.
3 讨论
细胞自噬的是一种溶酶体降解途径,对生存、分化、发育和机体稳态至关重要[12]。研究发现,自噬在许多疾病如感染、神经退行性疾病、癌症中起到保护作用[3,13]。当机体细胞自噬水平发生异常时,会诱发多种疾病,如细胞自噬减弱与阿尔兹海默症的发生密切相关[14,15]。生长因子、细胞内氨基酸水平、葡萄糖水平、DNA损伤、氧自由基等均可引起细胞自噬水平的变化。白桦脂醇是一种具有多种生物活性的天然提取物。大量研究显示,许多天然药物能促进细胞的增殖速率,如Lai等[16]发现,原花青素能抑制氧化应激反应,促进Th17和Treg细胞增殖,从而治疗银屑病。本文研究结果显示,白桦脂醇能提高AGEs作用下H9C2心肌细胞的活性。
当AGEs在体内含量异常升高时,会损伤血管内皮,促进白细胞黏附和血小板数量增加,促进动脉粥样硬化、糖尿病血管并发症,严重损害心血管系统。当心肌受到损伤时,心肌酶谱系列酶的浓度会有不同程度的升高,因而心肌酶谱常用来检测心肌损伤的情况,对心肌梗死的诊断具有参考价值。Mb、CK-MB和cTnⅠ是常见的心肌损伤标记物。许多研究证实,白桦脂醇能降低心肌组织中CK的表达。如Xia等[17]的研究结果显示,白桦脂醇能降低乳酸脱氢酶(Lactate dehydrogenase,LDH)和肌酸激酶(Creatinekinase,CK)的活性,对心肌缺血再灌注大鼠具有保护作用。与前面研究结果一致,本研究结果显示,白桦脂醇能降低AGEs作用下H9C2心肌细胞Mb、CK-MB和cTnⅠ的浓度,缓解AGEs诱导的H9C2心肌细胞的损伤。
细胞自噬是一个多基因参与的高度保守的复杂过程。Beclin1、P62和LC3是自噬发生过程中的3个重要蛋白。Beclin1是酵母自噬相关基因Atg6的同源基因,在自噬的起始阶段通过招募其他自噬相关基因形成复合物,介导其定位于自噬泡,调控自噬的起始[18,19]。LC3作为自噬标记物,在自噬体膜延伸阶段,LC3Ⅰ经过酶切转化为LC3Ⅱ,LC3Ⅱ与磷脂酰乙醇胺结合后,定位于自噬体膜上[20]。P62与LC3形成复合体,随着自噬的发生被降解,因此细胞内LC3Ⅱ/LC3Ⅰ的比值和P62的含量作为自噬发生的标志物[21]。Lin等[22]研究发现,白桦脂醇在小鼠体内具有镇痛、降低炎性反应的作用,同时抑制自噬水平。本研究结果显示,白桦脂醇添加到AGEs处理的H9C2心肌细胞后,Beclin1表达下调,P62含量上升,LC3Ⅱ/LC3Ⅰ下降,提示白桦脂醇可能会抑制AGEs诱导的H9C2心肌细胞自噬。
细胞自噬发生时,先形成一个杯状的前自噬体,吞噬聚集的蛋白和受损的细胞器,随后闭合成完整的自噬体,自噬体与溶酶体融合后,内容物在溶酶体作用下消化。正常生理状态下,LC3在胞质中呈现弥散的状态。当细胞发生自噬时,LC3Ⅱ与磷脂酰乙醇胺结合定位于自噬体膜[23]。通过免疫荧光染色,根据荧光颗粒的聚集程度和分布,不仅反映出细胞中LC3蛋白的表达情况,还能判断细胞自噬发生的状态。免疫荧光检测LC3实验结果显示,白桦脂醇能抑制AGEs诱导的H9C2心肌细胞自噬体的形成。
PI3K是一种胞内磷脂酰肌醇激酶,通过激活AKT从而参与细胞增殖、分化、凋亡等多种细胞的功能调节[24,25]。大量研究证实,白桦脂醇对PI3K/AKT信号通路具有调节作用。如Jin等[26]发现,从黄莲根中提取的白桦脂醇通过调节PI3K/AKT信号通路,能够抑制甲基黄嘌呤诱导的黑色素的生成。Majeed等[27]发现,白桦脂醇通过抑制AKT信号通路从而抑制癌细胞的扩散。Ci等[28]研究表明,白桦脂醇通过调节AKT依赖的信号通路从而抑制内毒素诱导的炎性反应。本文研究结果表明,白桦脂醇可能抑制AGEs对H9C2心肌细胞PI3K和AKT的磷酸化激活作用。
综上所述,白桦脂醇能促进H9C2细胞增殖,下调Beclin1的表达,抑制AGEs诱导的H9C2细胞自噬,缓解心肌细胞的损伤,其作用机制可能与调节PI3K/AKT通路的活化有关。下一步研究将探讨白桦脂醇在大鼠心肌梗死模型中对细胞自噬及氧化应激反应的影响。
