半滑舌鳎黏蛋白MUC5AC基因的克隆与结构预测
2019-07-17谭静胡秀彩吕爱军孙敬锋刘小雪廉正义
谭静,胡秀彩,吕爱军,孙敬锋,刘小雪,廉正义
半滑舌鳎黏蛋白基因的克隆与结构预测
谭静,胡秀彩,吕爱军通信作者,孙敬锋,刘小雪,廉正义
(天津农学院 水产学院 天津市水产生态及养殖重点实验室,天津 300384)
黏蛋白作为鱼类黏液的主要成分,在抵御病原感染过程中发挥重要作用。以半滑舌鳎()黏蛋白为研究对象,采用RT-PCR方法对其皮肤黏蛋白MUC5AC(即CsMUC5AC)进行基因克隆及结构预测分析。结果获得基因片段的长度为2 771 bp,推测编码917个氨基酸,预测所含5个抗原表位分布在314-321、507-516、578-589、748-762和811-826 aa,3个结构域主要为C8(164-238,624-698 aa)、VWD(1-128,430-588 aa)和VWC(303-365,768-836 aa)。二级结构预测C8区主要由α-螺旋构成、VWD区以β折叠为主,三级结构蛋白亚基主要由VWD-C8-VWC组成。构建系统发育树分析显示,其与大菱鲆、攀鲈、罗非鱼黏蛋白MUC5AC亲缘关系较近,与分泌型黏蛋白家族自然聚为一支。以上结果为研究半滑舌鳎黏蛋白基因功能提供科学参考。
半滑舌鳎;黏蛋白;基因克隆;结构预测
黏蛋白()作为机体黏膜免疫应答的主要组分之一,备受关注[1-3]。对人类和哺乳动物研究发现,黏蛋白与肠炎[1]、支气管炎[2]及多种癌症[3]等疾病发生密切相关。在水产养殖中,鱼类易受水体病原微生物感染,主要通过皮肤、鳃和胃肠道等黏膜组织侵入,这些组织中含有黏液细胞,分泌一种保护性黏液,其中黏蛋白作为黏液的主要成分,在抵御病原体感染过程中发挥着重要作用[4]。研究表明,鱼类皮肤黏液中含有多种活性物质,包括黏蛋白、多糖、免疫球蛋白及酶类等,对鱼类的许多生理、病理功能有着重要影响[5-6]。迄今为止,在人类和高等脊椎动物中,已发现二十多种黏蛋白,其被分为分泌型黏蛋白和膜结合黏蛋白两种类型[7],其中,分泌型黏蛋白一般由上皮杯状细胞和黏膜下腺细胞分泌形成[8]。目前认为鱼类体表黏液主要由分泌型黏蛋白组成,含有多个结构域包括VWD (Von willebrand D)、VWC (Von willebrand C)、C8 (Cysteine 8)、TIL(Trypsin inhibitor-like domain)、PTS(Proline- threonine-serine)等[9]。近几年,有对斑马鱼()[10]、团头鲂()[11]、黄颡鱼()[12]等多种鱼类黏蛋白基因克隆及表达的相关研究,但目前对半滑舌鳎黏蛋白基因尚未见文献报道。
半滑舌鳎是我国主要的海水经济鱼类,随着养殖规模日益扩大,病害问题也日趋严重[13]。半滑舌鳎作为一种底栖鱼类,其体表黏液不仅是物理屏障,而且能防御水体病原侵入,同时在渗透压调节等生理方面也起一定作用[14]。已有对鲤()、黄颡鱼基因克隆及特异性表达的研究,认为其在皮肤黏膜免疫应答过程中具有重要功能[12,15]。因此,本研究以半滑舌鳎为对象,对其皮肤黏蛋白基因克隆,对其基因编码序列进行蛋白结构预测分析,为今后深入研究基因功能提供科学参考。
1 材料与方法
1.1 材料
1.1.1 试验鱼
健康半滑舌鳎购自天津市海发珍品实业发展有限公司,体重(110±10)g,取其皮肤组织立即投入液氮冷冻,后转入-80 ℃冰箱保存,用于总RNA提取。
1.1.2 试剂
RNA提取试剂盒(TRIzol Reagent)、逆转录试剂盒、DNA Marker、PCR Buffer、pMD18-T载体等试剂购自TaKaRa公司。PCR Master Mix体系、UNIQ-10柱式胶回收试剂盒购自上海捷瑞公司。无水乙醇、氯仿、异丙醇等为分析纯产品。
1.2 方法
1.2.1 引物设计
根据半滑舌鳎基因编码序列(登录号:XM_025055866.1),用Primer 5.0软件设计3对特异性引物(表1),引物由苏州金唯智生物有限公司合成。

表1 半滑舌鳎CsMUC5AC基因引物序列的设计
1.2.2 皮肤总RNA提取和cDNA第一链合成
参照Neuwirth等[16]的方法,用TRIzol Reagent抽提RNA。总RNA质量通过琼脂糖凝胶电泳进行检测,其浓度及260/280值通过Nanodrop ND-2000分光光度计测定。然后以提取的总RNA为模板,用TaKaRa反转录试剂盒(PrimeScript TM II 1st Strand cDNA Synthesis Kit)合成第一链cDNA,合成的cDNA -20 ℃保存,备用。
1.2.3基因克隆
以RNA反转录cDNA为模板,3对特异性引物用于PCR扩增,PCR反应条件为:95 ℃预变性2 min;95 ℃ 40 s,55 ℃ 45 s,72 ℃ 90 s,30个循环;72 ℃延伸5 min。产物用1%琼脂糖检测,切胶回收,连接pMD18-T载体,转化到DH5α感受态细胞中,孵育、涂板,培养过夜后挑取单克隆,菌液PCR检测选取阳性单克隆后送公司测序。
1.2.4基因序列与结构预测
参照梁昱等[17]描述方法进行,采用ORF程序查找序列的开放阅读框,DNAStar软件进行蛋白抗原性、亲水性等理化特性分析,利用ExPASy Protseale、SMART等软件进行蛋白质结构域和三级结构预测,ClustalW及MEGA 6.0进行序列比对, Neighbour-Joining方法构建系统进化树。
2 结果
2.1 半滑舌鳎CsMUC5AC基因克隆
以半滑舌鳎皮肤组织cDNA作为模板,采用RT-PCR方法扩增基因得到3个目的片段,将其分别亚克隆至pMD18-T载体分别测序为893、1 305、715 bp,拼接获得片段长度为2 771 bp,推测编码917个氨基酸(图1),预测分子量为99.30 kD、理论等电点(PI)为4.86,蛋白不稳定系数为37.82、亲水性平均系数(GRAVY)为-0.234。DNAStar预测CsMUC5AC为亲水性蛋白,含有5个主要抗原表位(314-321、507-516、578-589、748-762、811-826 aa)。
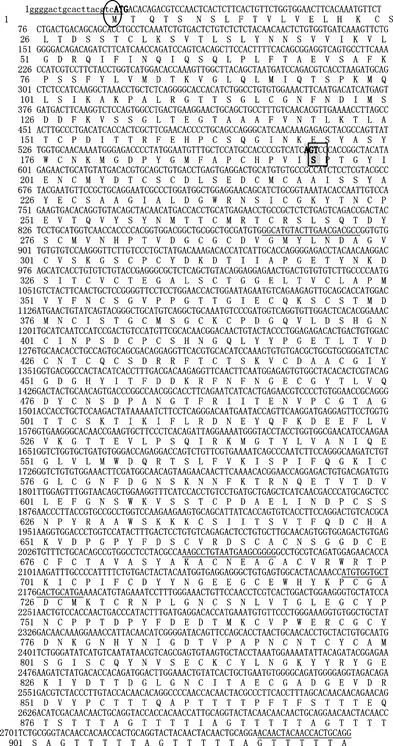
图1 半滑舌鳎CsMUC5AC基因的引物及编码氨基酸序列
注:A为CsMUC5AC基因的引物序列,B为CsMUC5AC基因的编码氨基酸序列,小写字母为UTR序列,椭圆为起始密码子,下划线为引物序列,方框为糖基化位点进一步对CsMUC5AC蛋白修饰位点预测表明,CsMUC5AC有1个糖基化位点(S189),当阈值≥0.9时共有13个磷酸化位点,其中6个丝氨酸(Ser)
磷酸化位点(S19/90/96/265/331/627),2个苏氨酸(Thr)磷酸化位点(T693/859),5个酪氨酸(Tyr)磷酸化位点(Y166/193/220/269/871)。此外,阈值≤0.5时,Ser磷酸化位点主要分布在200-400 aa和600-680 aa之间,Thr磷酸化位点主要分布在850-917 aa之间,Tyr磷酸化位点在740-820 aa之间显著分布。
2.2 MUC5AC多序列比对与系统发育树分析
对MUC5AC蛋白序列比对显示,含3个结构域,主要为VWD (pfam00094,1-128 aa,430-588 aa)、C8(pfam08742,164-238aa,624-698)和VWC(pfam00093,303-365,768-836)(图2A)。

图2 CsMUC5AC结构域及不同物种MUC5AC氨基酸序列多重比对分析
注:A为CsMUC5AC蛋白结构域,B为不同物种MUC5AC氨基酸序列多重比对;黑色为氨基酸同源性100%,深灰色为氨基酸同源性75%以上;实粗线方框指VWD结构域,虚线方框指C8结构域,实细线方框指VWC结构域;*为半胱氨酸
其中半滑舌鳎CsMUC5AC与大菱鲆(,AWP02052.1)同源性最高,为68.52%,其次为攀鲈(,XP_026234507.1),为67.60%,与罗非鱼(,XP_019210361.2)和雪鲷(,XP_024655160.1)的同源性分别为66.32%和64.69%。其与人(,AGR44427.1)、小鼠(,XP_006508562.1)、鸡(,XP_025006868.1)和非洲爪蟾(,XP_018113011.1)同源性分别为50.75%、51.68%、51.80%和51.22%,MUC5AC蛋白序列比对结果见图2B。
进一步选择人、鼠、大菱鲆、攀鲈、罗非鱼等18个黏蛋白基因序列,采用MEGA 6.0软件构建N-J系统进化树,结果显示半滑舌鳎CsMUC5AC与大菱鲆亲缘关系较近,自然聚为一支,属于分泌型黏蛋白家族,并且与膜结合型黏蛋白亲缘关系相对较远(图3)。

图3 邻接法构建不同物种黏蛋白的系统进化树
2.3 CsMUC5AC蛋白结构预测
对CsMUC5AC蛋白进行二级结构预测,表明以β-折叠为主,其中C8区主要由α-螺旋构成,VWD、VWC区主要由β折叠构成(图4A)。三级结构有2个蛋白亚基,主要由VWD-C8-VWC结构域构成,且与二级结构预测基本一致(图4 B)。
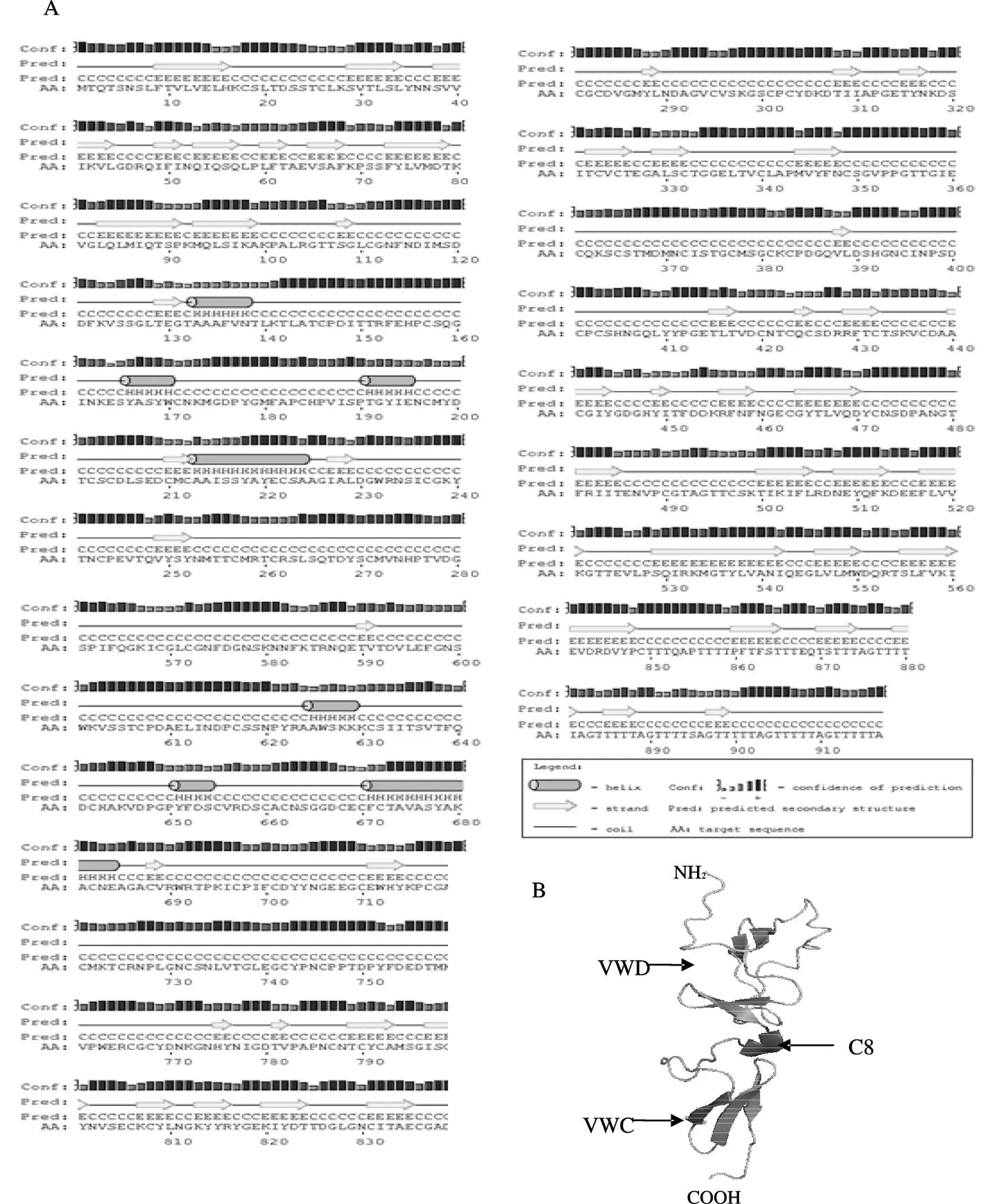
图4 CsMUC5AC蛋白的结构预测
注:A为CsMUC5AC蛋白二级结构预测,圆柱为α-螺旋,箭头为β折叠;B为CsMUC5AC蛋白的三级结构预测
3 讨论
鱼类皮肤及黏液是先天免疫反应的第一道防线,其中黏蛋白不仅抑制细胞表面与病原结合,而且刺激细胞吞噬作用[18]。研究表明,黏蛋白家族主要有分泌型黏蛋白(MUC2、MUC5AC、MUC5B、MUC6和MUC19)和膜结合型黏蛋白(MUC1、MUC3、MUC4、MUC12、MUC13、MUC14、MUC15、MUC16、MUC17和MUC18)[19]。对人类和哺乳动物研究表明,黏蛋白与多种疾病发生密切相关,主要分泌呼吸道黏蛋白为MUC5AC和MUC5B[1-3]。Pérez-Sánchez等[20]报道分泌型黏蛋白为动物上皮细胞生长、修复和存活提供防御作用,且在免疫应答中发生表达变化。Mejías-Luque等[21]对幽门螺杆菌感染小鼠胃肠黏膜MUC2、MUC4和MUC5AC等黏蛋白表达变化研究,提示MUC5AC是胃癌发生蛋白标志物之一。Matsukita等[22]发现MUC5AC黏蛋白表达水平与人乳腺癌有关。Imai等[23]发现MUC5AC在结直肠癌患者中显著性上调表达。黏蛋白作为鱼类机体黏膜免疫应答主要组分之一,对其基因克隆及功能研究备受关注[11,15,24]。Marel等[15]对鲤、基因克隆表达研究,表明在皮肤中显著性上调表达,而在鳃和肠道中显著性差异表达。薛春雨等[11]对团头鲂基因克隆及表达分析,表明捕捞应激后在皮肤和鳃中含量差异表达。Duan等[24]对南美白对虾()、和基因表达进行研究,发现外界压力胁迫下分泌型黏蛋白基因表达变化显著。目前,未有对半滑舌鳎黏蛋白基因克隆表达研究的报道。本研究对半滑舌鳎皮肤黏蛋白CsMUC5AC基因克隆,并对其编码蛋白质进行序列和结构预测,为深入研究基因功能打下基础。
对分泌型黏液蛋白结构分析显示,基因蛋白具有特定的C8、VWD结构域序列,且富含PTS重复序列[11]。本研究获得CsMUC5AC黏蛋白基因推测编码917个氨基酸,预测含有保守结构域VWD(pfam00094)、C8 (pfam08742)和VWC(pfam00093),这与鲤、团头鲂等文献报道基本一致[11,15]。研究表明,黏蛋白VWD、C8和VWC结构域相对保守,且与黏液蛋白基因类型及功能密切相关[9]。同时,用黏蛋白氨基酸序列构建系统发育进化树,分析显示半滑舌鳎CsMUC5AC属于分泌型黏蛋白家族,且与大菱鲆、鲈等鱼类黏蛋白亲缘关系较近。鱼类分泌型黏蛋白基因功能有待进一步研究。
本研究对半滑舌鳎皮肤黏蛋白基因克隆与结构预测等生物信息学分析,结果表明CsMUC5AC属于分泌型黏蛋白,主要由VWD-C8- VWC模式构成蛋白亚基,并对其糖基化、磷酸化位点及主要抗原表位分析,为深入研究鱼类黏蛋白基因功能奠定基础,对鱼类疾病免疫预防研究具有重要意义。
[1] Lee H S,Lee H K,Kim H S,et al. MUC1,MUC2,MUC5AC,and MUC6 expressions in gastric carcinomas[J]. Cancer,2015,92(6):1427-1434.
[2] Bonser L R,Zlock L,Finkbeiner W,et al. Epithelial tethering of MUC5AC-rich mucus impairs mucociliary transport in asthma[J]. Journal of Clinical Investigation,2016,126(6):2367-2371.
[3] Betge J,Schneider N I,Harbaum L,et al. MUC1,MUC2,MUC5AC,and MUC6 in colorectal cancer:expression profiles and clinical significance[J]. Virchows Archiv,2016,469(3):255-265.
[4] 吕爱军,胡秀彩,孙敬锋,等. 鱼类皮肤免疫应答及蛋白质组学[J]. 水产科学,2016,35(3):302-307.
[5] 杨桂文,安利国. 鱼类黏液细胞研究进展[J]. 水产学报,1999(4):403-408.
[6] Welsh K G,Rousseau K,Fisher G,et al. MUC5AC and a glycosylated variant of MUC5B alter mucin composition in children with acute asthma[J]. Chest,2017,152(4):771-779.
[7] McGuckin M A,Lindén S K,Sutton P,et al. Mucin dynamics and enteric pathogens[J]. Nature Reviews Microbiology,2011,9(4):265-278.
[8] Hovenberg H W,Davies J R,Carlstedt I. Different mucins are produced by the surface epithelium and the submucosa in human trachea:identification of MUC5AC as a major mucin from the goblet cells[J]. Biochemical Journal,1996,318(1):319-324.
[9] Allen A,Hutton D A,Pearson J P,et al. Mucus glycoprotein structure,gel formation and gastrointestinal mucus function[J]. Ciba Foundation Symposium,1984,109(12):137.
[10] Lang T,Klasson S,Larsson E,et al. Searching the evolutionary origin of epithelial mucus protein components- mucins and FCGBP[J]. Molecular Biology & Evolution,2016,33(8):1921-1936.
[11] 薛春雨,董晶晶,习丙文,等. 团头鲂黏蛋白基因克隆及表达分析[J]. 动物学杂志,2014,49(6):886-896.
[12] Cao X J, Wang W M. Histology and mucin histochemistry of the digestive tract of yellow catfish,[J]. Anatomia Histologia Embryologia,2010,38(4):254-261.
[13] Han Z R,Sun J F,Lü A J,et al. Isolation,identification and characterization offrom reared tongue sole,Günther[J]. Aquaculture,2017,468:356-362.
[14] Sveen L R,Grammes F T,Ytteborg E,et al. Genome-wide analysis of Atlantic salmon () mucin genes and their role as biomarkers[J]. Plos One,2017,12(12):e0189103.
[15] Marel M V D,Adamek M,Gonzalez S F,et al. Molecular
cloning and expression of two β-defensin and two mucin genes in common carp (L.) and their up-regulation after β-glucan feeding[J]. Fish & Shellfish Immunology,2012,32(3):494-501.
[16] Neuwirth L S,Phillips G R,Abdeslem E I . Perinatal Pb2+exposure alters the expression of genes related to the neurodevelopmental GABA-shift in postnatal rats[J]. Journal of Biomedical Science,2018,25(1):45.
[17] 梁昱,刘燕霏,孙晓莉,等. 犬链球菌M蛋白基因的克隆及生物信息学分析[J]. 天津农学院学报,2018, 25(1):44-48.
[18] Benhamed S,Guardiola F A,Mars M,et al. Pathogen bacteria adhesion to skin mucus of fishes[J]. Veterinary Microbiology,2014,171(1-2):1-12.
[19] Hjelmeland K,Christie M,Raa J. Skin mucus protease from rainbow trout,Richardson,and its biological significance[J]. Journal of Fish Biology,1983,23(1):10.
[20] Pérez-Sánchez J,Itziar E,Redondo M J,et al. Mucins as diagnostic and prognostic biomarkers in a fish-parasite model:transcriptional and functional analysis[J]. Plos One,2013,8(6):e65457.
[21] Mejías-Luque R,Lindén S K,Garrido M,et al. Inflammation modulates the expression of the intestinal mucins MUC2 and MUC4 in gastric tumors[J]. Oncogene,2010,29(12):1753-1762.
[22] Matsukita S,Nomoto M,Kitajima S,et al. Expression of mucins(MUC1,MUC2,MUC5AC and MUC6)in mucinous carcinoma of the breast:comparison with invasive ductal carcinoma[J]. Histopathology,2010,42(1):26-36.
[23] Imai Y,Yamagishi H,Fukuda K,et al. Differential mucin phenotypes and their significance in a variation of colorectal carcinoma[J]. World J Gastroenterol,2013,19(25):3957-3968.
[24] Duan Y,Liu Q,Wang Y,et al. Impairment of the intestine barrier function in,,exposed to ammonia and nitrite stress[J]. Fish & Shellfish Immunology,2018,78:279-288.
Cloning and sequence analysis of mucingene from the half-smooth tongue sole,
TAN Jing, HU Xiu-cai, LÜ Ai-junCorresponding Author, SUN Jing-feng, LIU Xiao-xue, LIAN Zheng-yi
(Tianjin Key Lab of Aqua-Ecology and Aquaculture, College of Fisheries, Tianjin Agricultural University, Tianjin 300384, China)
As the main component of fish mucus, mucin plays an important role in protecting from pathogen infection. The MUC5AC gene of, namely CsMUC5AC gene was cloned by RT-PCR method and protein structure was predicted by bioinformatics analysis. Results showed that the length of thegene fragment was 2 771 bp, encoding 917 amino acids(aa), and 5 antigenic epitopes were predicted to be mainly located within 314-321, 507-516, 578-589, 748-762 and 811-826 aa sites. The 3 domains including C8(164-238, 624-698 aa), VWD(1-128, 430-588 aa)and VWC(303-365, 768-836 aa)were observed. Analysis of secondary structure showed that the C8 motif was α-helix, the VWD domain was β-sheet, and the tertiary structure was two subunits profiles(VWD-C8-VWC). The phylogenetic tree analysis showed that CsMUC5AC was closely related tothat of,and, and naturally clustered into one group with the secretory mucins. These results provide a scientific reference for the further study ofgene function in tongue sole.
; MUC5AC; gene cloning; structure prediction
S917.4
A
1008-5394(2019)02-0051-06
10.19640/j.cnki.jtau.2019.02.012
2019-01-27
天津市自然科学基金重点项目(18JCYBJC29900,16JCZDJC33500);天津市水产生态及养殖重点实验室开放基金项目(TJAE201803);天津市水产产业技术体系创新团队(ITTFRS2017009);天津农学院综投平台项目(J01009030638)
谭静(1995-),女,硕士在读,主要从事水产动物微生物与免疫学的研究。E-mail:1109849082@qq.com。
吕爱军(1973-),男,教授,博士,主要从事水产动物微生物与免疫学的研究。E-mail:lajand@126.com。
责任编辑:张爱婷
