波动光下棉花叶片比叶重对光合生产能力的调控
2019-07-11王婧璇雷长英张玉洁韩吉梅张亚黎
王婧璇,雷长英,张玉洁,韩吉梅,张亚黎
(石河子大学农学院/新疆生产建设兵团绿洲生态农业重点实验室,新疆石河子832003)
引言
光是植物进行光合作用的基础。在自然环境中,由于太阳高度角和云层的变化,植物叶片一直处在波动光的环境下[1]。尽管波动光只持续几秒钟到几分钟,但它占叶片接受总太阳光能的20%~80%,能被叶片有效吸收和利用[2-3]。植物通常通过调整叶片结构和功能来适应波动光环境[4-6]。
研究[7-13]表明,波动光降低了植物叶片光能利用效率。同时,Cao[14]研究表明波动光降低了叶片CO2同化速率从而抑制了马铃薯(SolanumtuberosumL.)植株的生长;研究[9,15-16]也表明波动光下,水稻(Rice)叶片光合速率的降低导致叶片面积、叶片厚度、叶片数目和生物量显著降低。然而,也有研究[8,17]发现一些植物叶片的光合能力未受到波动光的影响。
有研究指出,比叶重是调控植物光合能力的重要因素。Vialet-Chabrand[12]研究表明波动光未改变拟南芥(Arabidopsisthaliana)叶片单位面积的饱和净光合速率(Asat),同时降低了单位面积最大光合速率(Amax),但基于单叶质量Asat和Amax均显著增大。除此以外,并不是所有植物对波动光的响应都呈现上述变化,一些研究显示波动光下生长的番茄[13](Solanumlycopersiumcv.Cappricia)、拟南芥[18]和非洲菊[9](Gerberajamesonii)等植物的最大净光合速率显著增强。表明,波动光下不同植物类型的光合作用响应机制存在差异。
棉花具有喜光特性,然而未见针对波动光下棉花光合作用的研究工作。本文针对波动光下棉花光合生理机制的变化,研究波动光下棉花叶片结构及生理特征的变化,揭示波动光下棉花光合特征的适应机制,研究结果将丰富棉花光能利用理论,为进一步提高棉花的光能利用效率奠定理论基础。
1 材料和方法
1.1 植物材料和生长条件
在人工气候室内进行试验,供试材料为常规抗病中早熟陆地棉品种“欣抗4号”。在土壤∶蛭石∶珍珠岩体积之比为1∶1∶1的花盆中种植棉花,花盆直径18 cm,高约25 cm。人工气候室日间温度(27±2) ℃,夜间温度(25±2) ℃。光周期为日间12 h,夜间12 h,湿度范围为50%~75%。每周施用营养液1次(1∶100 dilution,20-5-10-2 N-P-K-Mg,霍格兰营养液),每2 d浇水1次,使植物保持良好的生长状态。
1.2 波动光处理
本试验设置3个光处理:恒定光强(对照组,CK),生长光强为1000 μmol·m-2·s-1;每2 h恒定光间隔5 min低光(低光的平均光照强度为80~100 μmol·m-2·s-1)(T1);每2 h恒定光间隔10 min低光(T2),CK 为恒定光处理,T1和T2为波动光处理。恒定光和波动光处理下的植物每日接收总光能相同(43.2 mol·m-2·d-1)。
1.3 测定项目及方法
1.3.1 气体交换测定
光响应曲线的测定:用LI-6800便携式光合测定系统和荧光叶室测定,空气流速为500 μmol·s-1, CO2浓度设定为400 μmol·mol-1。测定时光强由强到弱,依次设定光合有效量子通量密度(PPFD)为1000、900、700、500、400、300、200、150、100、50、0 μmol·m-2·s-1。测定时每1光强下停留1~3 min,待gs恒定时记录数据。每处理测定3~4个叶片。采用Photo assistant软件对测得的光响应曲线进行拟合,并计算其光补偿点(LCP)、光饱和点(LSP)、饱和净光合速率(Asat)和表观量子效益(AQY)。
1.3.2 比叶重的测定
用面积0.36 cm2的打孔器避开叶片主叶脉,随机打5个叶圆片,装入牛皮纸袋中,在80 ℃烘箱中烘干48 h,用电子天平进行称重,计算比叶重(LMA)。比叶重=叶圆片干重/圆片面积。每个处理设4~5个重复。
1.3.3 单位叶片质量A的计算
将生长4周后单位叶片面积上的净光合速率(A)转化为单位叶片质量上的叶片净光合速率(Amass):
1.3.4 叶面积和生物量的测定
棉花植株生长至8周,光处理35 d,每个处理选取长势均匀一致的植株4~5株,将每株全部的叶片取下并用叶面积仪LI-3100(USA)扫描叶片面积,并记录数值。每个处理选取4~5株。将棉株于105 ℃下杀青30 min,80 ℃下烘干至恒重后称重测定其单株生物量。
1.3.5 每日叶片碳预算A24;mmol·d-1):
A24=[(Aavg×12)-(R×12)]×(3600/1000)×LA
(1)
Aavg(μmol·m-2·s-1)是每个处理重复间在12 h光周期中测得的净叶光合速率的平均值。R(μmol·m-2·s-1)是叶片暗呼吸,暗适应下叶片的净光合速率。LA(m2)是叶片的面积。Aavg和R分别乘以光周期(12 h)和暗周期(12 h),从每秒的速率变化到每小时的速率(3600 s),从μmol到mmol (1000)。
1.4 统计分析
通过单因素方差分析,比较各处理间棉花生长参数的差异。利用Duncan法对不同光处理下棉花的比叶重、生物量进行多重比较(显著度为0.05)。所有的分析均在SPSS22.0软件(SPSS Statistics22.0,SPSS Inc.,USA)和Sigmaplot12.5软件中完成。
2 结果与分析
2.1 波动光下棉花叶片特征和光合物质积累的变化
由表1可以看出,与恒定光条件相比,在波动光条件下棉花生长受到了抑制,表现为叶片面积和干物质重量均显著降低。分析叶片的比叶重可以看出,不同处理条件下的比叶重存在显著差异,恒定光下的比叶重显著高于波动光处理,同时波动频率快的比叶重也高于波动频率慢的处理。进一步将比叶重分解为叶片厚度和密度可以看出,波动光对叶片厚度未见显著影响,但是波动光下叶片密度发生较大的变化。
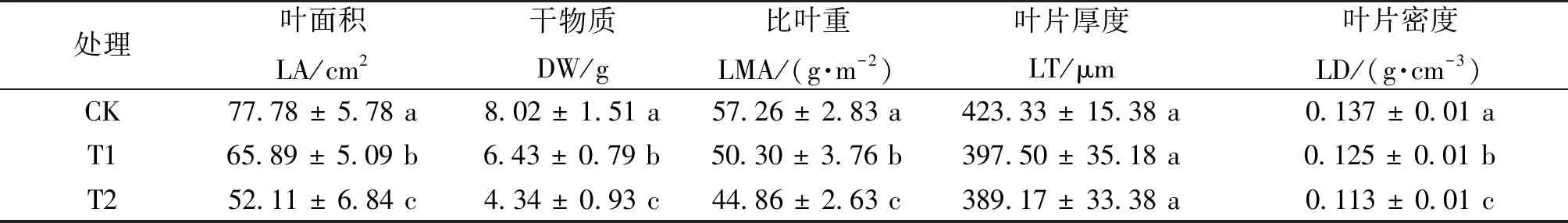
表1 不同处理下棉花叶片特性和生物量的变化Tab.1 Changes of leaves characteristics and biomass (DW) of cotton under different treatments(Mean±SD,n=3-4)
2.2 波动光下棉花气体交换参数的变化
在不同光处理下测定的棉花光响应曲线(图1 A)可以看出,当PPFD在200 μmol·m-2·s-1以下时,生长在这3种不同光处理下的棉花有着相似的单位叶面积净光合速率。而随着光强梯度的升高,3种光条件下的棉花对光强的响应逐渐产生差异。在PPFD达到400 μmol·m-2·s-1时,恒定光下的净光合速率显著高于波动光处理,在同一光强下,波动频率快的光合值也高于波动频率慢的处理。单位面积净光合速率在3个处理间存在显著差异,当其转换为单位叶片质量净光合速率时,波动光下的光合速率升高,但在不同光照处理下未见明显差异(图1B)。

图1 不同处理下棉花叶片的A-Q响应曲线及单位叶片质量净光合速率(Amass) (平均值±标准差,n=3-4)Fig.1 The A-Q response curve and photosynthesis per unit of leaf volume(Amass) in different treatments of cotton leaves(Mean±SD,n=3-4)
由表2可知,与CK相比,波动光处理下的叶片的Asat、LSP显著降低,LCP在T2处理下与恒定光处理之间存在明显差异,而在3种光条件下棉花叶片的AQY未呈现出显著差异。

表2 不同处理下棉花叶片光响应曲线拟合参数的变化Tab.2 Changes of light response curve fitting parameters in different treatments of cotton leaves(Mean±SD,n=3-4)
2.3 波动光下棉花生长的碳预算
通过计算植株叶片碳预算(A24,mmol·d-1),确定了整株植物的光能利用效率的差异。由表3可知,在波动光下生长的叶片Aavg比在恒定光下生长的叶片低29%和38%,而在3个光处理中,棉株的R几乎相同。根据Vialet-Chabrand等[12]人提出的计算公式计算棉花的每日碳预算,结果显示,波动光下的碳增益显著低于恒定光下的值,而这种差异是由较低的净光合速率以及由于较小的叶片(表1)导致。

表3 不同处理下植物叶片的每日碳预算Tab.3 Daily leaf carbon budget (A24,mmol·d-1) of plants growing under different treatments,as calculated by Eqn.1.
3 讨论
3.1 波动光下棉花光合物质生产能力降低
植株生物量的积累通常受到总叶片面积与比叶重的影响[33]。本研究表明,与恒定光相比,波动光减弱了棉花的光合物质生产能力,且波动频率越低对生物量的积累抑制越强(表1)。这与波动光下棉花光合作用降低导致的叶片面积的减少,比叶重的降低有关。波动光下生长的植物叶片明显变小变薄[9,12-13]。通过对各处理的碳固定进行估算可以看出,虽然各个处理接受的总光能是相同,但波动光下生长的光合能力和整体光能利用率均较低[10-12],导致棉株的碳固定损失较多,最终光合物质生产能力降低(表3)。
植物的生长与其碳固定能力密切相关;当总光能相同时,波动光下生长的植株碳同化能力通常低于恒定光照下生长的植株[14,19]。本研究表明,不论是在低光还是高光下,波动光下棉花的A、Asat均显著降低(图2),这说明波动光下棉花的光合能力确实受到了抑制。同时,有研究[6,18]表明,光合能力的大小也依赖于高和低光强度之间的转换频率,即较短的光波动引起的响应比较长的光波动小,低强度波动光引起的反应小于高强度波动光。在高光生长的植物中观察到的更高的Asat值,这通常与光合组分的数量有关,包括Rubisco,细胞色素f,H+-ATPase和反应中心[20]。在本研究中,与恒定光照相比,波动光下棉花净碳增益和地上总生物量较低,可能是由于较低的气孔导度(图2)和光合响应时间比光强变化率慢[9]。本研究中,在波动和恒定的光照下,植物叶片的最大表观量子产量没有发现差异,这表明当植物没有受到胁迫时,量子产量都是相似的[21]。
3.2 波动光下棉花叶片比叶重的调控
研究[22]表明,比叶重是与叶片的光合作用,生长和生活史策略相关的关键功能性状。比叶重是一个复合的生理结构参数,与叶片的很多解剖结构有关,因此对光合作用有潜在影响[23-24]。大量研究[25-27]发现植物在适应光环境的过程中主要通过调整叶片形态和结构来适应光照的变化,主要表现在比叶重的增大或减小。研究[28]认为,叶片比叶重的形成与每日光累积以及光照强度瞬时变化有关。与12 h/12 h的光/暗(方波光)条件相比,拟南芥的比叶重在波动的光线下降低[12],表明在波动光下有效光能积累量减少。本研究表明,波动光下棉花的比叶重与恒定光处理下的相比显著降低,说明在一定总光能范围内,光照供给形式的改变也会影响植物比叶重的形成。波动光下棉花光能利用效率降低,单位叶片面积上的光合速率下降,这与比叶重的研究结果基本一致,而波动光下的单位叶片质量净光合速率(Amass)与恒定光下的相比有明显的上升趋势[12,29-30]。这表明比叶重是构成波动光下棉花光合能力改变的重要因素。
叶片厚度和密度是决定植物比叶重大小的主要因素[31]。本研究发现,叶片厚度在不同光照处理下无明显差异,但叶片密度与比叶重呈现一样的变化规律,这说明本试验条件下比叶重主要受到叶片密度的调控,从而导致棉花的光合能力减弱[32-33]。有研究表明,高光条件下叶片密度的变化与叶脉密度有关[34];另外,可能还与栅栏组织细胞排列的紧密程度及植物非结构性碳水化合物(TNC)浓度的变化有关[33]。因此,叶片密度的大小也是影响植物光合作用的重要因素,但仍需进一步试验探究与叶片密度变化相关的指标。
综上所述,波动光下,叶片的光合适应过程本身是一个复杂而连续的现象。本研究表明,波动光抑制棉花的光合能力,导致其光合物质生产能力降低,同时降低叶片比叶重来调控波动光下棉花的光合能力。
