SIRT5 基因在苏姜猪组织中的表达分布研究
2019-06-18王利红曹玉娇
王利红,张 伟,2,曹玉娇
(1.江苏农牧科技职业学院,江苏泰州 225300;2.江苏姜曲海种猪场,江苏泰州 225300)
沉默蛋白(Sir2-related Enzymes,Sirtuin)或者沉默信息调节因子2(Silence Information Regulator 2,Sir2)最初是在酿酒酵母中发现的一类高度保守的烟酰胺腺嘌呤二核苷酸(NAD)依赖的组蛋白去乙酰化酶(HDAC)。此后研究发现,从古细菌到人类的生命活动,沉默蛋白都发挥着重要作用。哺乳动物有7 种Sirtuin同源基因SIRT1-SIRT7,它们具有不同的亚细胞定位和功能[1]。这些蛋白能够结合转录因子、信号转导因子、细胞骨架蛋白等多种因子,通过去乙酰化反应或ADP(Adenosine Diphosphate)核糖转移反应参与能量代谢、基因沉默、DNA 损伤修复、细胞周期调控和细胞分化等功能过程,是生物学功能十分广泛并且作用较为重要的酶类[2]。SIRT5 因具有更大的赖氨酸酰基结合口袋的结构特点,使其除了具有较弱的去乙酰化酶活性外,还有很强的去琥珀酰化、去丙二酰化和去戊二酰基化酶活性,通过不同的酶活性,参与调控葡萄糖、氨基酸、脂质等多种物质代谢过程,并发挥抗氧化、调控细胞和线粒体凋亡和炎性反应等生物学功能[3-5]。截至目前,有关猪SIRT5 基因在组织中表达的研究较少,且尚未见在苏姜猪组织中表达的研究报道。本研究以苏姜猪为研究对象,进行SIRT5 基因组织表达和定位检测分析,旨在从分子遗传方面深入了解SIRT5基因在猪组织中的表达特征,进而为优质猪种的选育提供理论参考。
1 材料方法
1.1 组织样的采集 选取体重在85 kg 左右的苏姜猪6头(公、母各半)。屠宰后立即采集猪的心脏、肝脏、脾脏、肺脏、肾脏、大肠、小肠、腹脂肪、背最长肌、腿肌、胃、卵巢(母猪)、子宫(母猪)、输卵管(母猪)等组织,每个组织3 个重复,分装在 1.5 mL 的Eppendorf 管(EP 管)中,放入液氮进行保存和运输。在实验室中将内脏组织存储在-80℃条件下,用于组织总 RNA 和蛋白质的提取。同时采集各组织样本置于10%中性福尔马林溶液中固定,用于后续免疫组织化学检测。
1.2 实时荧光定量PCR 根据GenBank 提供的猪SIRT5 基因序列使用Primer Premier 5 设计引物(表1),由上海英骏生物技术有限公司合成。根据说明书,使用TRIzol(TaKaRa)提取所有受试猪各组织样品中的总RNA。取1 000 ng 总RNA,按照SuperScriptTMIII First-Strand Synthesis SuperMix for qRT-PCR 试剂盒说明操作,Real-Time PCR 扩增反应体系为20 μL:上、下游引物(10 μmol/L) 各0.5 μL,cDNA 1.0 μL,Power SYBR®Green Master Mix 10.0 μL;反应条件为95℃,1 min;40 个 循 环(95 ℃,15 s,63 ℃,25 s), 反 转录成cDNA 于-70℃保存备用。

表1 实时荧光定量PCR 引物
1.3 酶联免疫分析(ELISA) 利用ELISA 对苏姜猪各组织中的蛋白水平进行测定:①取适量组织块,于预冷的PBS(0.02 mol/L,pH 为7.0~7.2)中清洗去除血液,称重1.0 g 组织用于后续实验(组织块较大需先剪碎后再匀浆)。②首先将组织块移入10 mL EP 管中,加入5 mL 预冷PBS 及10 µL PMSF 蛋白酶抑制剂后,使用电动匀浆机10 000 r/min 间歇匀浆 3 min,每次1 min,该过程在冰上进行。得到的匀浆液再利用超声破碎30 s(冰浴)。③将制备好的匀浆液于离心机中13 000 r/min 离心5 min,取上清用于ELISA 检测。④蛋白提取液中SIRT5 的蛋白水平使用SIRT5 酶联免疫分析ELISA 试剂盒测定,即在酶标包被板上的待测样品孔中先加稀释液40 µL,然后再加待测样品10 µL,之后在每孔中加入酶标试剂50 µL,封板膜封板后置37℃温育30 min,加洗涤液静置30 s 后弃去,重复洗涤5 次,拍干,每孔加入显色剂A 和B 各50 µL 混匀,37℃避光显色10 min,最后每孔加终止液50 µL,终止反应后以450 nm 波长测定样品吸光度。
1.4 免疫组织化学检测 取10%中性福尔马林溶液中固定的各组织脏器,经常规石蜡包埋,5 μm 厚切片。石蜡切片经脱蜡和水化处理后,用Tris-EDTA(pH=9)作为抗原修复液,微波加热至98℃并保持10 min,自然冷却至室温后,TBS 洗涤3 次,用3% H2O2封闭内源性过氧化物酶。TBS 洗涤3 次后,以5% BSA 于37℃封闭30 min,弃掉封闭液,使用兔抗SIRT5 抗体(CST,1∶250)作为一抗4℃孵育过夜。使用HRP 标记的羊抗兔IgG(南京生兴生物技术有限公司,sn134,1∶100)作为二抗识别特异性一抗,DAB 显色液显色。以苏木精复染细胞核后,梯度酒精脱水,中性树胶封片,光学显微镜下观察染色结果。
1.5 统计分析 SIRT5 mRNA 转录水平数据以心脏组织中表达量为标准,并以2-△△Ct计算。使用SPSS 19.0 单因素方差分析(ANOVA)及独立样本t 检验,进行差异显著性分析;用双变量Pearson 相关分析,P<0.05 表示差异或相关显著。
2 结 果
2.1 苏姜猪各组织中SIRT5 mRNA 转录和蛋白表达水平通过RT-PCR 及ELISA 方法,在苏姜猪心脏、肝脏、脾脏、肺脏、肾脏、大肠、小肠、腹脂肪、背最长肌、腿肌、胃、卵巢(母猪)、子宫(母猪)、输卵管(母猪)等组织中均检测到SIRT5 的表达(图1、图2)。
由图1 可看出,苏姜猪公猪SIRT5 mRNA 转录水平在心脏和肝脏组织中有丰富的表达,显著高于其他组织(P<0.05);其次肺脏、大肠、小肠和胃组织的SIRT5 mRNA 转录水平高于脾脏、肾脏、腹脂肪、背最长肌和腿肌组织(P<0.05)。从各组织中SIRT5 蛋白表达水平看,胃和大肠组织中表达量相对最高,均高于脾脏、肾脏、腹脂肪、背最长肌、腿肌组织(P<0.05),小肠和肺脏组织中表达量相近,与其他组织中表达量差异不显著,腹部脂肪中表达量相对最低,低于心脏、大肠和胃组织(P<0.05)。
由图2 可看出,苏姜猪母猪的小肠组织中SIRT5 mRNA 表达量相对最高,显著高于其他组织(P<0.05);其次在心脏、肺脏、肾脏、大肠、胃、卵巢、输卵管组织中也有较丰富的表达量,但与其他剩余组织间差异不显著。从各组织中SIRT5 蛋白表达结果看,心脏、肺脏、肾脏、大肠、小肠、子宫和输卵管组织中的表达量高于肝脏、脾脏、腹脂肪、背最长肌、腿肌、胃和卵巢组织(P<0.05)。
由表2 可看出,苏姜猪公猪各组织SIRT5 mRNA转录水平与蛋白表达水平间未达到线性显著相关(P>0.05),而母猪各组织SIRT5 mRNA 转录水平与蛋白表达水平间则呈线性极显著相关性(P<0.01)。

图1 SIRT5 在苏姜猪公猪不同组织脏器中的mRNA 转录和蛋白表达水平

图2 SIRT5 在苏姜猪母猪不同组织脏器中的mRNA 转录和蛋白表达水平
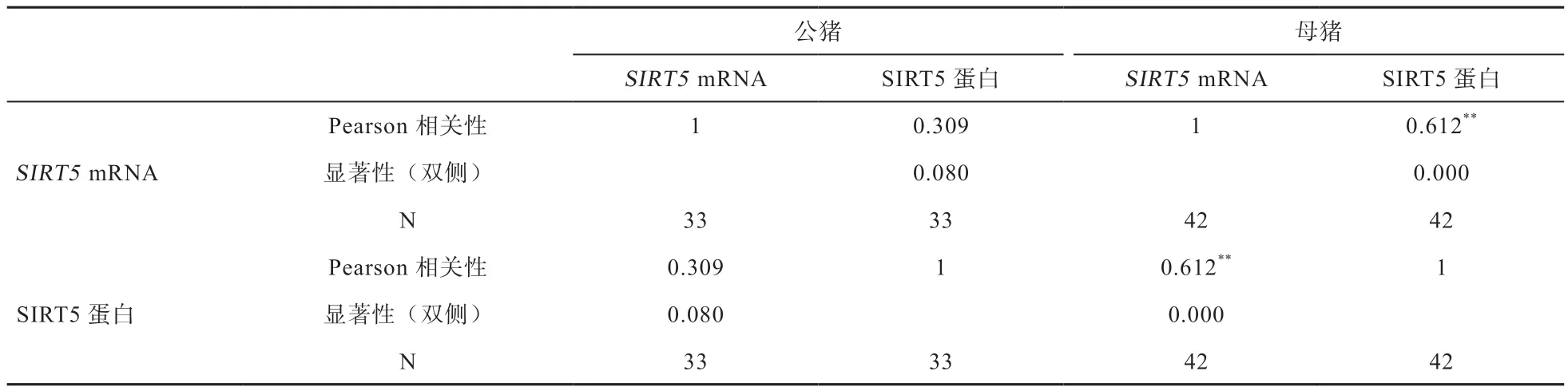
表2 苏姜猪SIRT5 mRNA 转录与蛋白表达相关性分析
2.2 SIRT5 在苏姜猪组织中的表达定位分布 通过免疫组织化学方法,在苏姜猪的心脏、肝脏、脾、肺、肾脏、胃、大肠、小肠、脂肪、背最长肌、腿肌、卵巢、输卵管、子宫等组织中均检测到SIRT5 免疫阳性细胞,且免疫阳性颗粒主要定位于各组织构成细胞的细胞质中(图3)。由图3 可看出,SIRT5 蛋白免疫阳性颗粒在苏姜猪多种脏器的组织细胞质中表达,且在肌肉细胞(如心脏的心肌细胞、背最长肌和腿肌的肌细胞、胃和肠的肌层细胞、子宫和输卵管的肌层细胞)有丰富的表达。此外,在肝脏的肝细胞、脾脏的红髓细胞、肺脏的肺泡壁上皮层和支气管黏膜层的细胞、肾脏皮质迷路的近曲小管内皮细胞、大肠和小肠肠腺和固有层细胞、脂肪组织的脂肪细胞、胃组织的固有腺层细胞、卵巢卵泡颗粒细胞以及子宫和输卵管组织的黏膜层细胞中也有丰富的免疫阳性颗粒。


图3 SIRT5 在苏姜猪多种组织内的免疫组化检测
3 讨 论
SIRT5 作为Sirtuins 家族中的一员,具有去乙酰化酶、去琥珀酰化酶、去丙二酰基化酶及去戊二酰基化酶活性,主要参与调控葡萄糖、脂质、活性氧(ROS)、ATP 等物质和能量代谢过程[6]。在动物组织中SIRT5表达具有广泛性,且在动物的心脏、肝脏、肌肉、脑以及生殖器官组织中呈高表达[3]。本研究通过RT-PCR、ELISA 和免疫组织化学方法在苏姜猪公猪11 个组织和母猪14 个组织中均检测到SIRT5 mRNA 转录和蛋白表达,表明苏姜猪组织中SIRT5 表达同样具有广泛性;通过相关性分析发现,苏姜猪母猪组织中SIRT5 mRNA相对表达量与蛋白表达量间呈线性极显著相关,表明该基因在苏姜猪母猪组织中的转录和翻译水平具有一致性,但公猪mRNA 相对表达量与蛋白表达量间则未呈显著线性相关,表明公、母苏姜猪不同组织细胞中SIRT5 调控机制存在差异。
本研究结果显示,苏姜猪公猪SIRT5 mRNA 在心脏和肝脏组织中表达量相对最高,显著高于其他组织,脾脏中表达量相对最低。这一结果与Jin 等[7]报道的通城猪公猪的SIRT5 mRNA 组织表达谱结果相似。然而,本实验中SIRT5 mRNA 在胃组织中的相对表达量则与通城猪公猪的结果不同,从ELISA 检测结果看,苏姜猪公猪胃组织中SIRT5 蛋白表达量也较高。对比苏姜猪公猪SIRT5 mRNA 和蛋白表达水平,脾脏和腹脂肪组织中表达量相对均最低,其次是背最长肌、腿肌和肾脏组织,而表达量差异相对较大的为心脏、胃、肝脏、肺脏、大肠和小肠组织。
在动物机体物质和能量代谢过程中SIRT5 发挥着重要的作用,SIRT5 缺失会导致动物机体几种代谢途径紊乱,且已有研究表明,SIRT5 可以作为秦川牛肉品质性状的候选基因,用于加速优质肉牛品种的选育[3-5,8-9]。本研究中,公、母苏姜猪脾脏、腹部脂肪、背最长肌和腿肌组织中SIRT5mRNA 表达量均相对较低,心脏、胃、肺脏和肝脏组织中有一定丰度的表达量,而在肾脏、大肠和小肠组织中表达量则母猪相对较高、公猪相对较低;母猪组织中的SIRT5 蛋白表达量总体相对低于公猪,这一点与母猪的生长势较公猪弱(如苏姜猪成年公猪平均体重204.68 kg,母猪体重168.71 kg)的特点相符。除生长势外,公、母猪在屠宰与肉品质方面也存在差异,如阉公猪的腹脂重显著低于母猪,而阉公猪肉的饱和脂肪酸含量显著高于母猪,且其肉色、贮存损失、熟肉率上均有一定优势,母猪则在大理石纹评分和失水率上稍有优势[10-11]。这种生长或肉品质性状差异是否与SIRT5基因表达有关,尚待后续进一步研究。
通过免疫组织化学方法,在所测定的苏姜猪组织中均检测到SIRT5 免疫阳性颗粒。肝脏是动物机体内重要的代谢器官。Nakagawa 等[12]研究表明,SIRT5 缺失的小鼠在长期饥饿后磷酸合成酶1(CPS1)的乙酰化程度升高,氨浓度增加,显示SIRT5 与肝脏中氨的代谢相关。本研究在苏姜猪肝细胞中检测到了丰富的SIRT5免疫阳性颗粒。在消化系统中,SIRT5 免疫阳性颗粒主要定位于苏姜猪胃组织的胃底腺层细胞和大肠、小肠的肠腺细胞,表明SIRT5 与腺细胞的分泌功能有关。机体最大的免疫器官——脾脏含有大量淋巴细胞和巨噬细胞,是机体细胞免疫和体液免疫的中心。通过细胞定位发现,SIRT5 免疫阳性颗粒主要分布于苏姜猪脾脏的红髓细胞。肌肉是动物机体能量代谢的主要组织。本研究在苏姜猪心脏、背最长肌和腿肌的肌细胞中均检测到SIRT5 免疫阳性颗粒,表明SIRT5 在苏姜猪能量代谢过程中具有重要作用。
氨基甲酰磷酸合成酶1(CPS1)是SIRT5 的一个重要底物,是尿素循环过程中重要的限速酶。SIRT5 不仅可以直接去乙酰化作用CPS1,增强其活性,而且在能量限制条件下还可以间接提高CPS1 的活性,限制尿素循环起始步骤,完成氨的解毒,在禁食或高蛋白质饮食期间,SIRT5 基因敲除小鼠的血液中有较高水平的氨,正是因CPS1 去乙酰程度降低,其活性降低所至[8,12]。当SIRT5 过表达时,CPS1 的戊二酰化水平降低,其酶活性升高[12-13]。通过细胞定位检测,SIRT5 免疫阳性颗粒主要在苏姜猪肾脏的皮质迷路近曲小管的内皮细胞中分布。在生殖系统中,SIRT5 免疫阳性颗粒主要分布于苏姜猪输卵管和子宫的黏膜层细胞中,表明SIRT5 与生殖道分泌功能有关,但其表达量相对不高。在卵巢组织中,SIRT5 免疫阳性颗粒主要分布于卵泡的颗粒层细胞,表明SIRT5 与颗粒细胞的功能有关,进而间接决定着卵母细胞的生长成熟;在卵巢黄体中未检测到SIRT5免疫阳性颗粒,表明该基因不参与黄体功能的行使。
4 结 论
SIRT5 在苏姜猪组织中具有表达广泛性,但仅母猪组织中SIRT5 转录与翻译水平呈极显著线性相关;在组织脏器中,在物质和能量代谢旺盛的组织(如胃、肠、肝脏、肌肉和肾脏等)中有相对较高的表达量,但母猪SIRT5 蛋白表达量总体低于公猪;SIRT5 免疫阳性颗粒主要定位于肝细胞、脾脏的红髓细胞、肺泡上皮细胞、近曲小管内皮细胞、胃和肠组织腺细胞、卵泡颗粒细胞、黏膜层细胞以及多种脏器的肌细胞中。
