细胞自噬在个体发育和肿瘤发生中的作用
2019-05-16李国辉祁馨宇邱立鹏
李国辉,李 东,祁馨宇,邱立鹏,唐 琦
(江苏大学生命科学研究院,中国江苏镇江212013)
1963年比利时细胞生物学家Christian de Duve在细胞分级分离时首次发现溶酶体,并于1974年与Albert Claude及George E.Palade一起荣获诺贝尔生理或医学奖[1~2]。细胞自噬(autophagy)就是通过细胞中产生的自噬体与溶酶体膜间的融合,进而利用溶酶体中的蛋白酶将自噬体中包裹的蛋白质或细胞器进行降解,其不同于泛素-蛋白酶体系统的选择性降解机制。该开创性工作是在单细胞生物酵母中完成的,其发现者为日本细胞分子生物学家Yoshinori Ohsumi(大隅良典),2016年诺贝尔生理或医学奖获得者[3~4]。细胞自噬是广泛存在于从酵母、线虫、果蝇到高等脊椎动物细胞中的一种维稳机制,在进化上具有高度保守性,其利用溶酶体中的各种水解酶对胞内蛋白质聚集体或折叠错误的蛋白质、受损的细胞器以及入侵的病原体等进行降解并回收利用[5~7]。细胞自噬异常与疾病的发生发展紧密相关,如肿瘤细胞的恶性增殖、神经退行性疾病等[8~10]。
细胞自噬是一种区别于泛素-蛋白酶体系统的降解途径。根据细胞内含物进入溶酶体腔方式的不同,细胞自噬可分为巨自噬(macroautophagy)、分子伴侣介导的自噬(chaperone-mediated autophagy,CMA)和微自噬(microautophagy)3 种(图 1)[11~13]。巨自噬是目前研究最为深入的一种细胞自噬方式,该过程涉及到吞噬泡的形成、自噬体的产生以及自噬体和溶酶体间的膜融合,最后形成自噬溶酶体,将其包裹的蛋白质聚集体或发生错误折叠的蛋白质进行降解[12~13]。分子伴侣介导的自噬包括分子伴侣对底物的识别、结合,并作用于溶酶体相关膜蛋白2A(lysosome-associated membrane protein 2A,LAMP-2A),随后在LAMP-2A复合物的作用下,将靶蛋白转位到溶酶体内进行降解。微自噬就是通过溶酶体直接将胞内折叠错误的蛋白质包裹进来,然后在溶酶体中对其进行降解[12~13]。
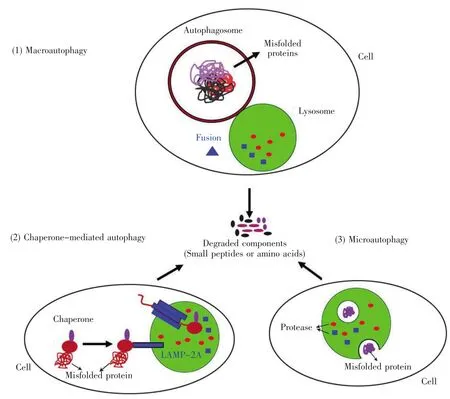
图1 三种类型的细胞自噬示意图(1)巨自噬能将错误折叠的蛋白质或蛋白质聚集体包裹起来形成自噬体,通过膜融合的方式介导溶酶体对其进行降解;(2)分子伴侣介导的自噬指的是含降解信号序列的蛋白质分子与热休克关联蛋白质分子发生结合,进而在胞质中招募分子伴侣并与溶酶体膜蛋白LAMP-2A结合,诱导其发生寡聚化,从而介导待降解蛋白质分子的转位和降解;(3)微自噬是指直接通过溶酶体对胞质中的一些蛋白质进行包裹并对其进行降解。Fig.1 Illustration of three types of autophagy(1)Macroautophagy is generally regarded as autophagy.In the process,double-membrane vesicles enclose some protein aggregates and misfolded proteins that are subsequently degraded after fusion with lysosome;(2)The chaperone-mediated autophagy is the degradation pathway for the target protein containing degradation signal.After proteins binding to LAMP-2A on lysosomes,they are translocated from cytosol into lysosome for degradation;(3)Microautophagy directly engulfs cytoplasmic components for degradation.
当遭受营养饥饿、组织缺氧、药物化疗和氧化应激等外界刺激时,细胞会被诱导产生自噬来进行自我保护,同时利用降解产生的氨基酸和腺嘌呤核苷三磷酸(ATP)来提供胞内蛋白质合成和细胞代谢所需的成分[14]。到目前为止,已在酵母和哺乳动物细胞中鉴定了40多个自噬相关基因(autophagy-related gene,Atg)。酵母中每个Atg基因的编码产物只有一种蛋白质形式,而哺乳动物细胞中一些Atg基因的编码产物却有多种异构体形式,这些蛋白质在自噬体的起始合成、蛋白质靶标的识别、自噬泡的延伸和自噬体与溶酶体膜间融合等过程中都发挥作用[15]。研究发现细胞自噬与人类疾病的发生发展密切相关。细胞自噬能降解胞内一些蛋白质聚集体,维持神经元的正常功能,一旦这些功能紊乱就容易引起老年痴呆、帕金森病、亨廷顿病等多种神经退行性疾病和人体自身免疫疾病的发生[16~18]。此外,细胞自噬与机体先天免疫和后天获得性免疫活性相关[19]。本文对细胞自噬形成的基本过程、分子调控机制及其在个体发育与肿瘤发生之间的关系进行综述,可为深入研究细胞自噬分子机制提供参考,并为人类疾病治疗提供新的靶点。
1 细胞自噬的发生与调控
细胞自噬活性不足或过度对机体的正常代谢都是有害的,因此,其生物活性在机体内受到严密监视和调控。目前,人们已在酵母、果蝇和哺乳动物细胞中对细胞自噬的诱导及调控进行了广泛深入的研究,其中TORC1和Ras/PKA等信号途径主要调控细胞自噬的活性水平。细胞自噬的动态过程可以人为地分为诱导、靶标识别、包裹、膜泡成核、膜泡延伸、Atg蛋白循环利用、与溶酶体膜融合、靶标发生降解和降解后组分的回收利用等几个主要阶段[20]。细胞自噬形成过程中最典型的一个生物学特征就是能够形成具有双层膜的运输泡,该囊泡在形成过程中受Atg蛋白的调控,依据这些调控蛋白质的差异可将其分为选择性的细胞自噬和非选择性的细胞自噬[21]。选择性的细胞自噬是通过自噬泡上受体蛋白质的特异性介导,将靶标包裹入囊泡中并对其进行降解;而非选择性的细胞自噬就是胞质中的靶标被随机性地包裹入自噬泡中并对其进行降解。
细胞自噬发生过程非常复杂,其囊泡不同于出芽方式产生的运输小泡,该过程牵涉到许多蛋白质的参与及调控,并在自噬泡组装位点(phagophore assembly site,PAS)形成自噬泡直至成熟。2013年,Hamasaki等[22]报道PAS介于内质网膜与线粒体膜的接触位点。现以巨自噬为例简要介绍自噬泡形成的过程[23~25]:在外部压力(如营养缺乏)条件下,哺乳动物雷帕霉素复合物1(mTORC1)失活,其下游的作用靶标ULK1/ULK2 (酵母中对应的是Atg1蛋白)复合物去阻遏,激活后的ULK1/ULK2复合物能作用于第Ⅲ类磷脂酰肌醇3激酶(PtdIns3K)复合物,该复合物包括Beclin 1(酵母中同源蛋白为Atg6)和VPS34(液泡分选蛋白34),当Beclin 1序列中Ser14发生磷酸化后,可促进PtdIns3K发生磷酸化产生PtdIns3P分子,并激活VPS34蛋白的生物活性,这一系列诱导都能被细胞自噬早期效应分子如Atg21和WIPI1/2 (酵母中为Atg18)等识别,并在PAS位点启动自噬泡的发生。
在酵母中,细胞自噬核心分子包括两个泛素样的蛋白质Atg8和Atg12,它们在Atg7激活之后,能介导自噬泡体积的变大。研究表明Atg8蛋白高度保守,酵母中只含有单个Atg8基因,其蛋白质产物C末端甘氨酸残基与脂质磷脂酰乙醇胺偶联,定位于酵母自噬泡表面,而N-端的螺旋结构能介导自噬体与溶酶体膜间融合,是自噬体形成非必需的,但能调控自噬体的颗粒大小;Atg12能招募并形成Atg12-Atg5-Atg16复合物,该复合物在Atg9蛋白的帮助下能正确定位到PAS 位点[26]。2017 年 Maeda 等[27]报道,Atg8 蛋白在细胞静止状态下也能够降解一定量的脂滴,但是当Atg8基因突变或缺失时则不会有此功能,表明Atg8蛋白具有不依赖细胞自噬途径来维持细胞中脂滴动态的活性。在哺乳动物细胞中,Atg8同源蛋白统称为微管蛋白1轻链3(light chain 3,LC3),分为两个亚家族,由6种蛋白质组成,其中LC3B(light chain 3B)蛋白研究最为广泛,与自噬体的组装和成熟有关,可决定自噬体中被包裹进来的蛋白质的特异性,同时可调控自噬体的大小。LC3B蛋白通常被用作细胞自噬活性的一个监测指标,为细胞自噬研究提供了一个有效的标记[13,28~29]。
2 细胞自噬在免疫系统方面的作用
细胞自噬在胞内病原体的去除、胞内分泌途径调控、淋巴细胞的发育和促炎症信号等方面都起着重要的作用。机体可通过直接吞噬胞内病原体,或者利用膜蛋白受体如Toll样受体(Toll-like receptor,TLR),介导参与胞内病原体的吞噬,产生的囊膜小泡与溶酶体融合形成自噬溶酶体,从而对胞内病原体进行清除或者降解[30~31]。
吞噬过程中产生的囊泡与分泌途径中的膜泡具有相似的结构和功能,因此,参与细胞自噬过程中的一些蛋白质可同时在吞噬作用和蛋白质分泌途径中起作用。2012年Castillo等[32]报道,Atg5基因缺陷小鼠的巨噬细胞能高水平表达白介素IL-1α,从而导致过度的炎症反应。抗原呈递细胞中抑制细胞自噬会促进胞内IL-1β和IL-23分泌因子水平的升高;而脂多糖刺激会抑制胞内IL-1β 因子的分泌水平[33~34]。2016 年 Lee等[35]报道,细胞自噬调控线粒体中活性氧的水平,在巨噬细胞炎症因子的分泌中起重要作用,胞内活性氧水平异常与多种疾病及肿瘤发生相关。
细胞自噬激活抗原呈递细胞,可将外来抗原有效地呈递在巨噬细胞、树突状细胞和B细胞的表面,从而影响淋巴细胞的发育。后天获得性免疫途径主要通过T淋巴细胞和B淋巴细胞发挥免疫作用,它们在发育过程中需要特定的激活,否则会导致人体自身免疫疾病的发生。T淋巴激活信号主要是通过一个主要组织相容性复合体(major histocompatibility complex,MHC)分子进行抗原呈递。该复合体分为MHCⅠ和MHCⅡ两类,其中MHCⅠ主要存在于有核的细胞中,而MHCⅡ存在于巨噬细胞、树突状细胞和B淋巴细胞中。胞内或胞外的一些蛋白质需要通过泛素降解途径或者细胞自噬进行降解,产生的肽段抗原才可呈递在细胞表面[36]。在巨噬细胞感染过程中,细胞自噬不仅能提高MHCⅠ分子呈递病毒抗原的能力,而且也能促进MHCⅡ呈递病毒抗原[37]。细胞自噬也是树突状细胞表面分子MHCⅡ与T细胞表面分子CD4结合所必需的;若树突状细胞中缺乏细胞自噬活性,则会降低MHCⅡ呈递外来抗原的能力[38~39]。另外,细胞自噬也是维持浆细胞(也叫效应B细胞)动态稳定和长期体液免疫所必需的,细胞自噬缺陷或过度活跃都会对浆细胞的发育和功能产生影响,从而导致人体自身免疫疾病的产生[40]。
近年研究表明,细胞自噬与NF-κB信号通路激活相关。2012年Paul等[41]报道,细胞自噬衔接蛋白Bcl10可调控T细胞受体(TCR)介导的NF-κB信号通路激活(一种高度保守的胞内降解途径)。2016年Zhong等[42]报道,胞内损伤线粒体的清除需要依赖细胞自噬衔接蛋白p62/SQSTM1,该蛋白质可调控NLRP3-炎性体的激活;若p62/SQSTM1发生缺失,则会导致炎性体和胞内IL-1β表达水平的增加。
3 细胞自噬可维持胞内蛋白质/细胞器动态平衡
胞内蛋白质合成与降解需要维持在一个动态平衡范围内,其与细胞自噬和泛素-蛋白酶体系统两条降解途径直接相关:泛素-蛋白酶体系统主要负责降解生命周期短的蛋白质[43];细胞自噬则可批量降解胞内生命周期长的胞质蛋白质和细胞器。对于3种不同类型的细胞自噬,它们的区别在于其囊膜是特异性还是非特异性地包裹胞中的内含物。细胞自噬功能异常或调控紊乱,容易导致人体各种疾病的发生[44~46]。
细胞自噬在功能正常的生理状态下充当细胞管家,对胞内错误折叠的蛋白质、氧化损坏的蛋白质以及功能异常的细胞器等进行严密监视,及时清除蛋白质聚集体和功能异常的细胞器,这不仅可避免细胞过早死亡,也有助于细胞寿命的延长,提高机体的各项生理机能[47]。在哺乳动物中,细胞自噬能降解胞内形成的一些蛋白质聚集体,如神经元细胞中的突触蛋白、突变的亨廷顿蛋白和tau蛋白,以及一些入侵的病原体如伤寒杆菌和结核杆菌等,从而降低神经退行性疾病和传染性疾病的发生概率;同时,细胞自噬也能降解胞内功能异常的线粒体、过氧化物酶体、脂滴、核糖体和部分细胞核[48]。细胞在正常的生理状态下,其自噬活性维持在一个正常的基础水平,将胞内一些功能异常的蛋白质和细胞器去除,控制着胞内质控系统;而当细胞出现炎症时,自噬活性急剧上调,从而维持胞内一个动态平衡[49]。随着人和酵母中自噬基因的鉴定,科研人员发现它们不仅直接参与细胞自噬体的形成,而且与组织动态变化、抗衰老、细胞分化、发育、先天免疫、后天获得性免疫、代谢疾病、心脏疾病、中风和造血功能缺陷等都有直接关联[50~53]。由此可见,机体各种器官的退化和衰老与细胞自噬功能降低呈正相关,目前尽管对细胞自噬抗衰老的分子机制仍不甚清楚,但对细胞自噬途径中相关靶分子进行调控或许是延长寿命的一种有效方式。
当机体处在紫外照射、营养饥饿和组织缺氧等逆境条件下时,细胞可以通过降解并回收利用自身蛋白质、脂类和碳水化合物等大分子物质,来提供机体生存所需要的基本成分和能量,是机体在环境不利的情况下做出的一种正常的反应[54~55]。当DNA发生损伤得不到及时修补,不断累积在细胞中时,会导致细胞老化、死亡甚至癌变,而细胞自身可以通过DNA修补、细胞凋亡和细胞自噬等分子机制来抑制该级联反应[56~57]。蛋白质的翻译后修饰(如磷酸化、泛素化、乙酰化和脂质化)可快速诱导细胞自噬发生,如:肿瘤抑制因子p53的靶标蛋白DAPk(death-associated protein kinase),其能对Bcl-2序列上119位丝氨酸以及蛋白激酶D(protein kinase D,PKD)进行磷酸化,从而导致Vps34-PI3K复合物磷酸化,进而启动细胞自噬的发生[58],直接将损伤的DNA进行降解,有效管控DNA损伤诱导的不利反应,维持细胞中DNA和蛋白质的代谢平衡。
4 细胞自噬影响个体发育
在一些模型生物中,人们发现细胞自噬缺陷会影响个体发育,如:酵母不能形成孢子、线虫体细胞中P颗粒不能发生降解以及果蝇幼虫致死和果蝇蛹变态发育成昆虫受阻等[59]。在哺乳动物中,细胞自噬在心脏、骨骼及神经系统发育和细胞分化中起着重要作用,如:红细胞来源于骨髓造血前体细胞,细胞自噬可对幼红细胞中的线粒体、核糖体和其他细胞器进行降解,从而产生成熟的红细胞,若自噬相关蛋白质功能异常,将会阻碍正常红细胞的产生,从而产生贫血症和其他血液疾病[60]。
在哺乳动物细胞中,细胞自噬对于胚胎发育、新生儿存活与器官发育都是非常重要的。哺乳动物受精后,细胞自噬与泛素-蛋白酶体系统协同降解精子中的线粒体,从而有助于线粒体DNA的异质性;受精卵发育到2个细胞之后,细胞自噬被高度激活,可降解母系来源的mRNA和蛋白质,这有助于合子基因组的活化[61~62]。Atg5基因缺失的小鼠胚胎在发育过程中不会超过8细胞阶段;细胞自噬对新生小鼠发育有重要影响,这主要是来源于小鼠母系的胎盘营养供应出现中断,使新生小鼠面临严重的饥饿挑战,而细胞自噬在乳汁喂养这个营养过渡期起重要的作用,神经功能缺陷和细胞自噬缺陷都会导致乳汁喂养失败[63]。另外,细胞自噬还有利于去除老旧组织,在细胞命运决定时与细胞凋亡协同发挥作用。
在果蝇幼虫变态发育过程中,其唾液腺的去除高度依赖细胞自噬介导的细胞死亡和细胞凋亡,然而中肠组织的降解仅依赖细胞自噬[64];在果蝇卵巢发育过程中,需要半胱天冬酶来诱导细胞发生自噬,而果蝇营养细胞的凋亡需要通过细胞自噬降解其凋亡抑制蛋白[65]。在大多数机体中,细胞自噬是一种重要的自我保护反应。为了说明细胞自噬是细胞死亡的主要原因,细胞自噬导致的细胞死亡也被称为Ⅱ型细胞死亡,这意味着化学或遗传上抑制细胞自噬的活性,细胞死亡也会受到抑制。细胞自噬能降解胞内的铁蛋白,导致活性氧在细胞内积累,从而引起细胞死亡。这种细胞防御机制也叫异体吞噬,能够选择性地识别胞内微生物包括细菌、病毒和原生生物,从而把它们传送到细胞自噬泡中进行降解[66]。
5 细胞自噬抑制肿瘤发生
细胞自噬是一种正常的生理生化过程,能清除机体中的一些异常细胞和胞内的异常蛋白质,对维持胞内组分的动态平衡和保障机体生存起重要作用。细胞自噬异常与人体许多疾病的发生发展密切相关,其中细胞自噬与抗肿瘤增殖活性的研究较为深入。
Yu等[67]报道,异喹啉类生物碱小檗碱(berberine)能上调原发性肝癌细胞中Beclin-1和LC3蛋白的表达,诱导细胞凋亡和细胞自噬来杀死肝癌细胞,从而抑制肿瘤细胞的迁移和发展。另外,通过对抗细胞凋亡的细胞系进行生物碱处理,结果发现在处理后的细胞中,细胞自噬流和细胞自噬体数量显著增多,同时胞内AMPK-mTOR途径也被激活,进而杀死这些细胞[68]。粉防己碱(tetrandrine)是一种草本植物根部的生物碱提取物,同时也是一种广谱有效的细胞自噬激动剂。Wang等[69]报道,低剂量的粉防己碱不会影响细胞的活力,但会诱导非小细胞肺癌A549细胞系、人前列腺癌PC3细胞系、乳腺癌MCF-7和MDA-MB-231细胞系、神经胶质瘤U87细胞系和宫颈癌HeLa细胞系中细胞自噬的发生,从而引发细胞死亡。另外,一些生物碱如苦参碱、槐果碱、槐定碱、氧化苦参碱、苦豆碱和金雀花碱都具有抗肿瘤生物活性,它们能够诱导一些肿瘤细胞如人白血病细胞系K562、U937和HL-60以及癌细胞系EC109、A549和HeLa发生细胞自噬,抑制肿瘤细胞的增殖,而且这种抗肿瘤效果与生物碱浓度呈正相关[70]。这些研究表明:临床上可以通过生物碱或其类似物对肿瘤进行治疗,诱导胞内发生细胞自噬来杀死肿瘤细胞,达到根除肿瘤的治疗效果。目前,已在人体多种肿瘤中发现细胞自噬异常的现象,其功能异常有利于肿瘤细胞的恶性增殖和迁移,说明细胞自噬在维持正常细胞生命活动和杀死异常细胞方面具有重要的作用。由此可见,将来有望在临床上通过激活细胞自噬途径,为肿瘤患者提供可行、有效的治疗方案。除此之外,细胞自噬还与神经退行性疾病、肝脏疾病、心力衰竭、糖尿病、肥胖症和自身免疫性疾病等多种疾病和炎症密切相关[12,71]。
6 小结与展望
细胞自噬是胞内的一种降解途径,与胞内物质代谢平衡、细胞存活和机体发育等生物过程都密切相关。细胞自噬功能异常会导致多种人类疾病的发生,已有报道显示许多重大疾病都与细胞自噬功能异常相关。尽管细胞自噬在过去20年里取得了巨大的进展,揭示了细胞自噬主要是通过溶酶体途径对功能异常的蛋白质和损伤的细胞器进行降解,然而我们对细胞自噬的认识仍停留在初级阶段,如自噬体膜的来源以及胞内来源的膜组分如何融入到自噬体膜中等科学问题仍不清楚,这些都有待于我们进一步去阐明。此外,在细胞自噬发生过程中,许多与细胞自噬相关的蛋白质都发生了翻译后修饰,然而这些修饰方式如何调控细胞自噬的活性也不甚清楚,或许通过揭示细胞自噬相关蛋白质的空间结构及其相互之间的作用,有助于阐明这些科学问题,从而为深入揭示细胞自噬在人体疾病中的发生和调控机制提供理论依据。
