Unconjugated bilirubin alleviates experimental ulcerative colitis by regulating intestinal barrier function and immune inflammation
2019-05-08JiaDongZhengYanHeHengYuanYuYuanLiLiuYiXuanGeXueTingLiXueLiYanWangMengRuGuoYiLinQuXiaoFaQinMingShanJiangXiuHongWang
Jia-Dong Zheng, Yan He, Heng-Yuan Yu, Yuan-Li Liu, Yi-Xuan Ge, Xue-Ting Li, Xue Li, Yan Wang,Meng-Ru Guo, Yi-Lin Qu, Xiao-Fa Qin, Ming-Shan Jiang, Xiu-Hong Wang
Abstrac t BACKGROUND Unconjugated bilirubin (UCB) is generally considered toxic but has gained recent prominence for its anti-inflammatory properties. However, the effects of it on the interaction between intestinal flora and organisms and how it influences immune responses remain unresolved.AIM To investigate the role of UCB in intestinal barrier function and immune inflammation in mice with dextran-sulfate-sodium-induced colitis.METHODS Acute colitis was induced by 3% (w/v) dextran sulfate sodium salt in drinking water for 6 d followed by untreated water for 2 d. Concurrently, mice with colitis were administered 0.2 m L UCB (400 μmol/L) by intra-gastric gavage for 7 d.Disease activity index (DAI) was monitored daily. Mice were sacrificed at the end of the experiment. The length of the colon and weight of the spleen were recorded. Serum level of D-lactate, intestinal digestive proteases activity, and changes to the gut flora were analyzed. In addition, colonic specimens were analyzed by histology and for expression of inflammatory markers and proteins.Committee of Harbin Medical University.RESULTS Mice treated with UCB had significantly relieved severity of colitis, including lower DAI, longer colon length, and lower spleen weight (colon length: 4.92 ±0.09 cm vs 3.9 ± 0.15 cm; spleen weight: 0.33 ± 0.04 vs 0.74 ± 0.04, P < 0.001). UCB administration inactivated digestive proteases (chymotrypsin: 18.70 ± 0.69 U/g vs 44.81 ± 8.60 U/g; trypsin: 1.52 ± 0.23 U/g vs 9.05 ± 1.77 U/g, P < 0.01), increased expression of tight junction (0.99 ± 0.05 vs 0.57 ± 0.03, P < 0.001), decreased serum level of D-lactate (31.76 ± 3.37 μmol/L vs 54.25 ± 1.45 μmol/L, P < 0.001), and lowered histopathological score (4 ± 0.57 vs 7 ± 0.57, P < 0.001) and activity of myeloperoxidase (46.79 ± 2.57 U/g vs 110.32 ± 19.19 U/g, P < 0.001). UCB also regulated the intestinal microbiota, inhibited expression of tumor necrosis factor(TNF) α and interleukin 1β (TNF-α: 52.61 ± 7.81 pg/mg vs 105.04 ± 11.92 pg/mg,interleukin 1β: 13.43 ± 1.68 vs 32.41 ± 4.62 pg/mg, P < 0.001), decreased expression of Toll-like receptor 4 (0.61 ± 0.09 vs 1.07 ± 0.03, P < 0.001) and myeloid differentiation primary response gene 88 (0.73 ± 0.08 vs 1.01 ± 0.07, P <0.05), and increased expression of TNF-receptor-associated factor 6 (0.79 ± 0.02 vs 0.43 ± 0.09 P < 0.05) and inhibitor of kappa B α (0.93 ± 0.07 vs 0.72 ± 0.07, P < 0.05)in the colon.CONCLUSION UCB can protect intestinal barrier function, regulate normal intestinal homeostasis, and suppress inflammation via the Toll-like receptor 4/ nuclear factor-κB signaling pathway.
Key words: Ulcerative colitis; Unconjugated bilirubin; Intestinal barrier; Intestinal homeostasis; Digestive proteases; Inflammation
INTRODUCTION
Inflammatory bow els d iseases (IBD), w hich includ es ulcerative colitis (UC) and Crohn's d isease (CD), are associated w ith chronic, relap sing inflammation of the intestinal tract. In the early 1900s, regions such as North America, Europ e, and Oceania had millions of individ uals w ith IBD. The prevalence of IBD is highest in the western w orld, w hich was estimated to affect up to 0.5% of the general population in 2015 and equates to approximately 2.2 million Americans living w ith IBD in 2025[1,2].How ever, new ly ind ustrialized countries have a low p revalence of IBD, but the incid ence has been stead ily rising. A representative stud y from the Asia-Pacific Crohn's and Colitis Epidemiology Study show ed that IBD w as established in Asia,with an average incidence of 1.4 per 100000 people in 2011, and the incid ence of UC was tw o-fold higher than that of CD in Asia and the incidence of IBD in China was 3.3 per 100000[3]. IBD affects millions of people around the world, producing a substantial burden on healthcare systems.
Evidence from studies on etiology and pathogenesis suggests that IBD results from a dysregulated intestinal immune response driven by complex interplay betw een the host and intraluminal microbiota[4]. Intestinal epithelial cells recognize conserved signature molecules in the gut commensal bacteria, called p athogen-associated molecular patterns, by pattern recognition receptors. Toll-like recep tors (TLRs) are one of the major typ es of pattern recognition receptor families and have a p ivotal effect on maintaining homeostasis of the gut microbiota[5]. During inflammation, TLR4 as an innate immune receptor is stimulated by recognition of gut pathogen-associated molecular patterns. TLR4 und ergoes a conformational change and then recruits the signaling adaptors myeloid d ifferentiation primary response gene 88 (My D88) and tumor necrosis factor (TNF) receptor-associated factor (TRAF) 6, thereby activating dow nstream transcription of nuclear factor-κB (NF-κB) target genes and expression of some pro-inflammatory cytokines such as interleukin 1β (IL-1β), TNF-α, or IL-6[6].How ever, anti-inflammatory and immuno-regulatory cytokines are related to the negative regulation of nitric oxid e (NO), w hich is positively correlated w ith the severity of the disease. In IBD patients, NO production was positively correlated w ith increased levels of pro-inflammatory cytokines[7]. The role of cytokines and NO level are fundamental to regulate inflammation in IBD. Meanwhile, TLR4 was significantly more upregulated in IBD patients than in controls, especially in UC patients[8,9]. The probiotic cocktail (Lactobacillus acidophilus, Lactobacillus plantarum, Bifidobacterium lactis,and Bifidobacterium breve) ameliorated clinical symptoms and histological scores,d ecreased NO level, and red uced TLR4, ind ucible NO synthase, and NF-κB expression[10]. An important mechanism of UC might be that abnormal inflammation by intestinal d ysbacteriosis causes d ysregulation of TLR that med iates innate immunity. The TLR4/NF-κB signaling p athw ay is an essential key p oint in the development of UC.
Unconjugated bilirubin (UCB) is generated during the physiological breakdow n of heme by heme oxygenase 1. Serum levels of bilirubin increase w ith the accelerated release of heme from hemoglobin or w ith d iminished hep atic conjugating activity.UCB is a potent antioxidant even at low concentrations[11,12]. It was first discovered in 1934 that bilirubin w as exerts anti-inflammatory activity in patients w ith rheumatoid arthritis w ho exp erienced remission of symp toms after d evelop ing jaund ice second ary to liver d isease[13]. Studies have show n that bile acid s play a role in the regulation of gut injury, immunity, and inflammation[14,15]; however, there is a lack of studies on the physiological function of bilirubin that enters the intestine together w ith bile. Recent investigations hav e d emonstrated that UCB sup p resses inflammatory responses in animal mod els of autoimmune encep halomyelitis and lung inflammation in asthma[16-18]. Nevertheless, the mechanisms on inflammation and role of UCB in the interaction betw een intestinal flora and immunity are not w ell understood. The aim of the present study w as to investigate the effects of UCB on intestinal barrier function and inflammation on dextran sodium sulfate (DSS)-induced colitis in mice.
MATERIALS AND METHODS
Animals
Male C57BL/6 mice aged 8-12 wk (w eight approximately 25 g) were purchased from the Experimental Animal Center of the Second Affiliated Hospital of Harbin Medical University and w ere acclimatized for 1 wk before experiments were performed. They w ere reared in the Animal Laboratory Centre of Harbin Med ical University und er specific pathogen-free conditions (temperature 24-25 °C, humidity 70%-75%, w ith a 12 h light/d ark lighting regimen) and w ere fed a stand ard d iet of pellets and water a d libitum. The study w as approved by the Institutional Animal Care and Use Committee of Harbin Medical University.
Chemicals and reagents
UCB, N-benzoyl-L-tyrosine ethyl ester, and N-α-benzoyl-L-arginine 4-nitroanilide hydrochloride were purchased from Sigma-Aldrich (St. Louis, MO, United States).DSS (36-50 k Da) was obtained from MP Biomedical (Solon, OH, United States).Enzyme linked immunosorbent assay (ELISA) kits for D-lactate, TNF-α, IL-1β, and myeloperoxidase (MPO) were from Beijing Propbs Biotechnology (Beijing, China).The antibodies used in this study were anti-TLR4 (19811-1-AP; Proteintech, Rosemont,United States), anti-My D88 (4283; Cell Signaling Technology, Danvers, MA, United States), anti-TRAF6 (ab33915; Abcam, Cambridge, MA, United States), anti-inhibitor of NF-κB alpha (IκBα) (4814; Cell Signaling Technology), and anti-occludin (ab167161;Abcam). Anti-glyceraldehyde 3-phosphate dehydrogenase (GAPDH), goat anti-rabbit immunoglobulin G and goat anti-mouse immunoglobulin G w ere purchased from ZSGB-BIO Co. Ltd. (Beijing, China). All other reagents used were of analytical grade.
Induction of colitis and pharmacological treatment
Colitis was induced by oral administration of DSS as described previously, with some modifications[19]. The animals w ere randomly divided into five groups with five mice in each group: Control group (Control), DSS group (DSS), DSS plus UCB group (DSS+ UCB), and UCB group (UCB). Colitis w as induced by ad ministering 3% DSS in d rinking w ater for 6 d, and then d rinking w ater w ithout DSS for another 2 d for recovery. From the first day, UCB was administered daily via gavage at 0.2 mL for 7 d(UCB w as d issolved in 0.4% d imethyl sulfoxid e at concentrations up to 400 μM).During the stud y, weight, physical condition, stool consistency, and the presence of occult blood in feces w ere examined and d ocumented d aily. All animals w ere sacrificed after 8 d by intrap eritoneal injection of an overd ose of chloral hyd rate.Blood specimens were collected and serum samples w ere prepared by centrifugation at 4000 g for 15 min at 4 °C and stored at -80 °C. The entire colon, spleen and total feces of mice were carefully removed, measured and w eighted, then stored at -80 °C for further analysis.
Disease activity index
Disease activity index (DAI) was calculated for each animal by measuring the body weight, stool consistency, and fecal blood. Each score was calculated based on the criteria listed in Table 1[20].
Histology analysis for scoring colonic damage
Colonic tissues fixed in 4% (w/v) paraformaldehyde were paraffin-embedded and sliced into 5 μm sections, followed by staining with hematoxylin and eosin for light microscopic examination to assess colon injury and inflammation. Colonic damage was graded in a blinded manner as described as Table 2[21].
Analysis of intestinal permeability
D-Lactate concentration in serum was measured by ELISA. Results are expressed as μmol D-lactate/mL serum.
Determination of fecal digestive proteases activity
Trypsin and chymotrypsin (amidase) activities were measured using N-α-benzoyl-Larginine 4-nitroanilid e hyd rochlorid e or N-benzoyl-L-tyrosine ethyl ester as the substrate, respectively, as described previously[22,23].
Colon RNA extraction and quantitative reverse transcription polymerase chain reaction
Total RNA was isolated from colon tissues using the UNlQ-10 Column Trizol Total RNA Isolation Kit (Sangon Biotech, Shanghai, China). After treating with DNase I(TaKaRa, Tokyo, Japan), RNA was transcribed into cDNA using Prime Script TM RT regent Kit (TaKaRa) using an Eppendorf Mastercycler personal Thermo cycler. All primers were obtained from Sangon. The primers were used as followed: forward: 5'-GAGCACCTTCTTTTCCTTCATCTT-3' and revers: 5'-TCACACACCAGCAGGTT
ATCATC-3' for IL-1β, forward: 5'-CATCTTCTCAAAATTCGAGTGACAA -3' and reverse: 5'-TGGGAGTAGACAAGGTACAACCC-3' for TNF-α and forw ard: 5'-CATGGCCTTCCGTGTTCCTA -3' and revers: 5'-GCGGCACGTCAGATCCA-3' for GAPDH. Quantitative real-time polymerase chain reaction (qRT-PCR) was performed in a volume of 20 μL with FastStart Universal SYBR Green Master (Roche, Basel,Sw itzerland). All samp les w ere analyzed in trip licate, and the results w ere normalized to the expression of GAPDH. Results were expressed as 2-ΔΔCt.
Fecal bacteria DNA extraction and qRT-PCR
Genomic DNA from feces w as extracted by Stool Genomic DNA Extraction Kit(Tiangen Biotech, Beijing, China) and used for quantitative analysis of fecal bacteria Firmicutes, Bacteroid etes, Actinomycetes, and Proteobacteria. All primers w ere obtained from Sangon. The p rimers w ere used as follow ed: forw ard: 5'-GCTGCTAATACCGCATGATATGTC-3' and reverse: 5'-CAGACGCGAGTCCAT
CTCAGA-3' for Firmicutes, forward: 5'-GAGAGGAAGGTCCCCCAC-3' and reverse: 5'-CGCTACTTGGCTGGTTCAG-3' for Bacteriod etes, forw ard: 5'-TGTAGCGGTGGA
ATGCGC-3' and r ev er se: 5'-AATTAAGCCACATGCTCCGCT-3' for Actinomycetes, forward: 5'-TAGGCTTGACATTGATAGAATC-3' and reverse: 5'-CTTACGAAGGCA

Table 1 Criteria for scoring disease activity index
GTCTCCTTA-3' for Proteobacteria and forward: 5'-GCAACGAGCGCAACCC-3'and reverse: 5'-ACGGGCGGTGTGTAC-3' for 16S rDNA. qRT-PCR in a volume of 20 μL was performed using FastStart Universal SYBR Green Master. All samples were analyzed in triplicate, and the results were normalized to the expression of 16S rDNA.The results were expressed as 2-ΔΔCt.
Cytokine levels measured by ELISA and Western blotting
Segments of colon w ere homogenized using rad ioimmunoprecicpitation assay buffer and protein inhibitor cocktail (1:10) (PhosSTOP ESAYpack; Roche). The homogenates were kept on ice for 30 min and centrifuged at 12000 g for 5 min at 4 °C. The protein concentration w as determined using the bicinchoninic acid assay Protein Assay Kit(Beyotime, Shanghai, China). For ELISA, the sup ernatants w ere collected and subjected to IL-1β, TNF-α, and MPO assays. Results of IL-1β and TNF-α w ere exp ressed as pg/mg and MPO as U/g. For w estern blot analysis, proteins w ere electroblotted onto a polyvinylid ene d ifluoride membrane following sep aration on 10% sodium dodecyl sulfate polyacrylamide gel electrophoresis. The immunoblot was then incubated w ith primary antibodies against occludin, TLR4, My D88, TRAF6, and IκBα, or GAPDH. The chemiluminescence signals were analyzed using Quantity One(version 4.5.2; Bio-Rad Laboratories, Hercules, CA, United States) and Image J software.
Statistical analysis
Results w ere exp ressed as mean ± stand ard error of the mean (SEM). Differences betw een group s w ere d etermined using one-w ay analysis of variance follow ed by Tamhane multiple comparisons post-hoc tests using SPSS version 19.0 (IBM, Armonk,NY, United States). Grap hs w ere analyzed using Grap hp ad Prism version 5.0(Graphpad Software, La Jolla, CA, United States). Statistical significance was denoted as P < 0.05.
RESULTS
UCB relieved clinical severity of DSS-induced colitis
Compared w ith the DSS group, the DSS + UCB group exhibited less w eight loss,diarrhea, and intestinal bleeding, as reflected by the significantly lower DAI scores (P< 0.001, Figure 1A and B). The tw o control groups that received regular d rinking w ater or UCB alone did not d evelop signs of colitis throughout the experiment (all DAI score < 1 from 1 d to 8 d, Figure 1B). Furthermore, UCB treatment significantly reduced colon shortening and increased spleen weight (P < 0.001, Figure 1C and D).
UCB inactivated digestive protease activity of DSS-induced colitis
Digestive proteases are believed to play a crucial role in the d estruction of the intestinal barrier[24,25], and UCB has previously been demonstrated to inactivate digestive proteases activity[26]. Therefore, we tested the effects of UCB on trypsin and chymotrypsin activity in the feces of mice with DSS-induced colitis. Compared with the DSS group, the DSS + UCB group showed remarkably lower activities of fecal digestive proteases trypsin and chymotrypsin, and levels in the UCB alone group were similar to the control group (P < 0.01, Figure 2A and B).
UCB ameliorated colon tissue injury in DSS-induced colitis
DSS-induced colon tissue injury was demonstrated by epithelial destruction, intense inflammatory infiltration, and crypt distortion (Figure 3A) as well as increased histological scores for inflammation and crypt damage (Figure 3B). DSS with UCB treatment reduced neutrophil infiltration and crypt damage in the colon, leading to a decrease in colonic MPO activity (P < 0.001, Figure 3C), which is an inflammatorymarker for colitis. The epithelial destruction, histological score, and MPO activity of the UCB alone group were similar to those in the control group.
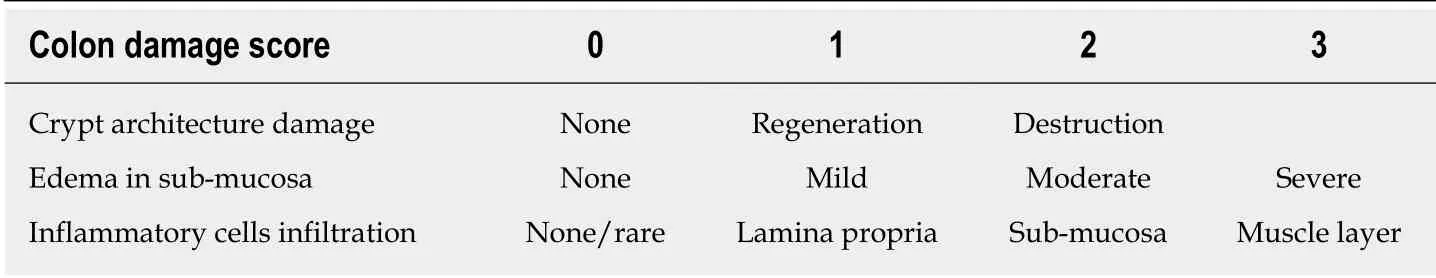
Table 2 Criteria for histology analysis for scoring colonic damage
UCB protected intestinal barrier function of DSS-induced colitis
Compared with the control group, expression of the tight junction protein occludin in the colon of the DSS group w as decreased significantly (P < 0.001, Figure 4A), and serum level of D-lactate w as significantly elevated (P < 0.001, Figure 4B). How ever, in the DSS + UCB group, exp ression of occlud in w as increased in the colon w ith a decrease in serum levels of D-lactate (P < 0.001, Figure 4A and B). Results in the UCB alone group were similar to those in the control group (Figure 4).
UCB regulated fecal microbiota composition of DSS-induced colitis mice
We assessed the change in fecal microbiota in mice at the phylum level. DSS induced an overall decrease in feces microbiota. Compared with the controls, Firmicutes and Actinomycetes were dominant in the intestinal flora (P < 0.05), while Bacteroidetes (P< 0.01) and Proteobacteria (P < 0.05) w ere less. The intestinal flora of UCB treated mice recovered approximately. How ever, Firmicutes (P < 0.01) w ere decreased and Actinomycetes (P < 0.001) w ere increased comp ared w ith the DSS group.Bacteroidetes in the UCB alone group were also higher than in the control group, but the difference w as not significant (Figure 5).
UCB reduced production of proinflammatory cytokines and inhibited TLR4/MyD88/TRAF6/NF-κB signaling in DSS-induced colitis mice
The mRNA and protein levels of the pro-inflammatory cytokines, TNF-α and IL-1β, in colon tissues of mice were determined. DSS-induced acute colitis w as accompanied by a significant increase in TNF-α and IL-1β levels (P < 0.001), and treatment with UCB restored these cytokines to normal levels (P < 0.001, Figure 6A, B). TLR4 is a key immune recep tor that p lays an imp ortant role in the regulation of colonic inflammation[27]. Compared with the control group, expression of TLR4 (P < 0.01) and My D88 (P < 0.05) was increased in the DSS group, and expression of TRAF6 (P < 0.01)and IκBα (P < 0.05) w as decreased. In comparison, expression of these proteins w as restored to near normal in the UCB-treated group. The results in the UCB only group were similar to those in the control group (Figure 7).
DISCUSSION
Previous observation on human evolution has shown that bilirubin-predominant species are often carnivores or omnivores, while biliverdin-predominant species are often herbivores[28], suggesting that bilirubin may have an important yet unknown effect on the human body. Significant increases in concentration of serum bilirubin in neonates (> 20 mg/d L, approximately 340 μM) can trigger nerve damage[29]; however,it is a powerful antioxidant and toxic in adults[11,30,31]. UC patients have reduced total serum bilirubin levels[32]. Numerous studies have suggested that UCB plays an important potential protective role in vascular endothelial function[33]; ameliorates allergic lung inflammation in a mouse model of asthma[18]; prevents murine colitis by inhibiting leukocyte infiltration and suppressing upregulation of inducible NO synthase[34]; or scavenges various reactive oxygen species[12].
Our previous w ork confirmed that UCB ameliorates the inf lammation in trinitrobenzenesulfonic acid-induced colitis; an animal model of CD[35]. In this study,we further established the UC mouse model that has a different extent of ulceration and tissue edema. Our data suggest two potential mechanisms by which UCB exerts its protective effect on DSS-induced colitis: (1) Inactivation of digestive proteases to protect intestinal mucosal barrier and prevent intestinal flora translocation to the colon lead ing to an abnormal inter action; and (2) sup p ression of the TLR4/My D88/TRAF6/NF-κB signaling pathway to inhibit immune inflammation and regulate intestinal microflora homeostasis.
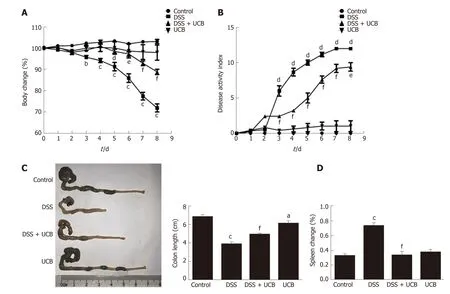
Figure 1 UCB significantly relieved severity of dextran sulfate sodium induced colitis. A: Body weight changes, data plotted as percentage of basal body weight; B: Disease activity index; C: Changes in colon length; D: Relative spleen weight. Data are expressed as means ± SEM (n = 5). a P < 0.05, b P < 0.01, and c P <0.001 vs control group, d P < 0.05, e P < 0.01, and f P < 0.001 vs dextran sodium sulfate group. UCB: Unconjugated bilirubin; DSS: Dextran sodium sulfate; SEM:Standard error of the mean.
The intestinal barrier is a complex multilayer system, consisting of an external anatomical barrier and an inner functional immunological barrier. The interplay of these tw o barriers maintains normal intestinal function and a stable intestinal environment[36]. We d etected that the intestinal tract of UC mice contained elevated active d igestive p roteases, w hich can d estroy the intestinal tissue structure and increase intestinal permeability. This provides favorable cond itions for intestinal pathogens or toxins of microbiota invading the body. The large number of digestive proteases as vital factors contributes to the pathogenesis and development of IBD due to d estruction of the intestinal barrier. UCB can be a sp ecific inhibitor to inactivate d igestive p roteases to p rotect the p hysical barrier function that is the first line of defense against invasion of intestinal pathogens.
Another p ivotal factor, intestinal microbes, p articip ates in many imp ortant physiological processes, including nutritional absorption, substance metabolism, and immunity[37,38]. In ord er to investigate how UCB may sp ecifically contribute to intestinal microbiota during DSS colitis, w e tested the composition and abundance at the p hylum level (Bacteroid etes, Firmicutes, Actinomycetes and Proteobacteria).Exposure to DSS reduced the diversity and abundance of intestinal flora. The ratio of Bacteroid etes to Firmicutes w as d ecreased and that of Proteobacteria to Actinomycetes w as increased. UCB treatment can reverse these phenomena; however,it d ramatically red uces the content of Firmicutes and makes Bacteroid etes and Actinomycetes dominant in the flora. Actinomycetes have alw ays produced many antibiotics[39]; therefore, w e suspect that UCB can promote Actinomycetes to produce some secondary products to protect mice against colitis. Our study show s that UCB is beneficial to maintain the stability of intestinal flora, but its specific role need s to be examined through further research that our group has and w ill continue to be concerned.
TLRs are the recognition receptors of the innate immune system and are located on the surface of various immune cells and play an imp ortant role in d efense against infection and regulation of immune responses. TLR4 expression is usually low under normal cond itions, w hile in IBD, the intestinal f lora lose tolerance and sp ecif ic recognition of lipopolysaccharid e, then prod uce excessive signal transduction, and f inally activ ate TLR4 and cascad e ind uce transcrip tion of NF-κB and p roinf lammatory cytokines[40,41]. Our results demonstrate that UCB decreases expression of TLR4 and My D88, and increases expression of TRAF6 and IκBα in the colon. In ad d ition, UCB red uces levels of pro-inflammatory cytokines, TNF-α and IL-1β,suggesting it has an anti-inflammatory role in DSS-induced colitis.
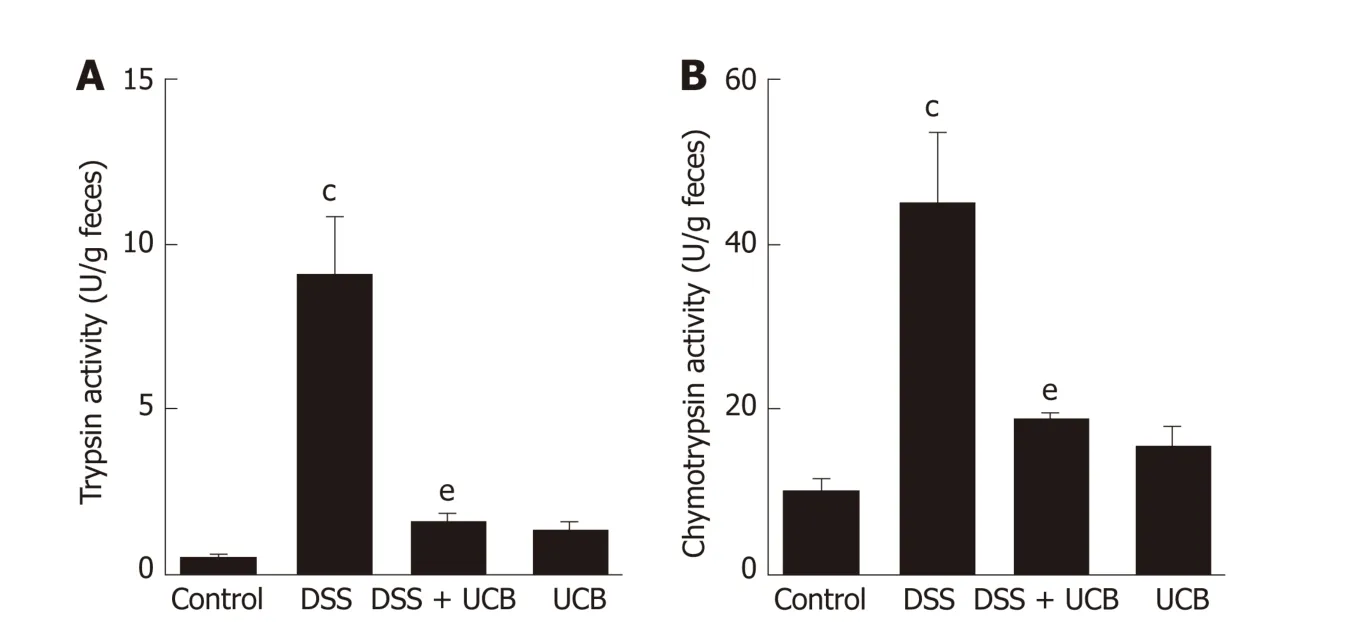
Figure 2 UCB inactivated digestive protease activity in DSS treated mice. A: Trypsin activity in feces; B:Chymotrypsin activity in feces. Data are expressed as means ± SEM (n = 5). c P < 0.001 vs control group and e P <0.01 vs dextran sodium sulfate group. UCB: Unconjugated bilirubin; DSS: Dextran sodium sulfate; SEM: Standard error of the mean.
UCB can suppress intestinal positive immune response and inflammation through the TLR4/NF-κB signaling pathw ay. Therefore, this app ears to be an ad d itional mechanism of UCB to regulate the relationship betw een intestinal flora and host immunity. UCB can imp rove intestinal inflammation, regulate homeostasis of bacterial flora, and maintain intestinal immune function; it is a w ay to resolve the serious problem of ad verse effects and d rug resistance of IBD. Perhaps low dose of UCB or in combination w ith other p robiotics can d evelop healthcare to prevent clinical diseases. It is a novel therapeutic approach, not only for UC but also for other gastrointestinal diseases.
This study had some limitations. First, w e found that UCB affected the intestinal flora but the change in genera was unclear. Second, we only induced acute UC, w hich may have different mechanisms from chronic UC. Third, this stud y shows that UCB protects intestinal barrier function and sup presses immunity and inflammation in DSS-induced colitis, but we did not study how UCB regulates intestinal immune cells and other potential pleiotropic roles of UCB. These issues will be studied in our future research.
In conclusion, our find ings not only shed light on how UCB ameliorates UC but also offer a clue about the physiological function of UCB. This provides a noteworthy example of how UCB can serve as a “double-edged sw ord” in the human body. This is believed to be the first study to show that UCB ameliorates DSS-induced colitis via inactivation of d igestive proteases to protect the intestinal barrier and regulate the intestinal flora and TLR4/NF-κB signaling p athw ay to sup p ress intestinal inflammation.
ARTICLE HIGHLITHTS
Research background
Clinical treatment of ulcerative colitis consists of drugs that are both expensive and have side effects. Unconjugated bilirubin (UCB) has gained recent prominence for its anti-inflammatory and antioxid ant properties. How UCB influences UC remains unresolved.
Research motivation
Patients with UC require lifelong treatment, and drugs for UC are linked to many adverse effects. Therefore, there is an urgent need to develop effective and safe drugs for UC.
Research objectives
To investigate the significance of UCB in intestinal barrier function and immune inflammation of mice with dextran sodium sulfate (DSS)-induced colitis.
Research methods
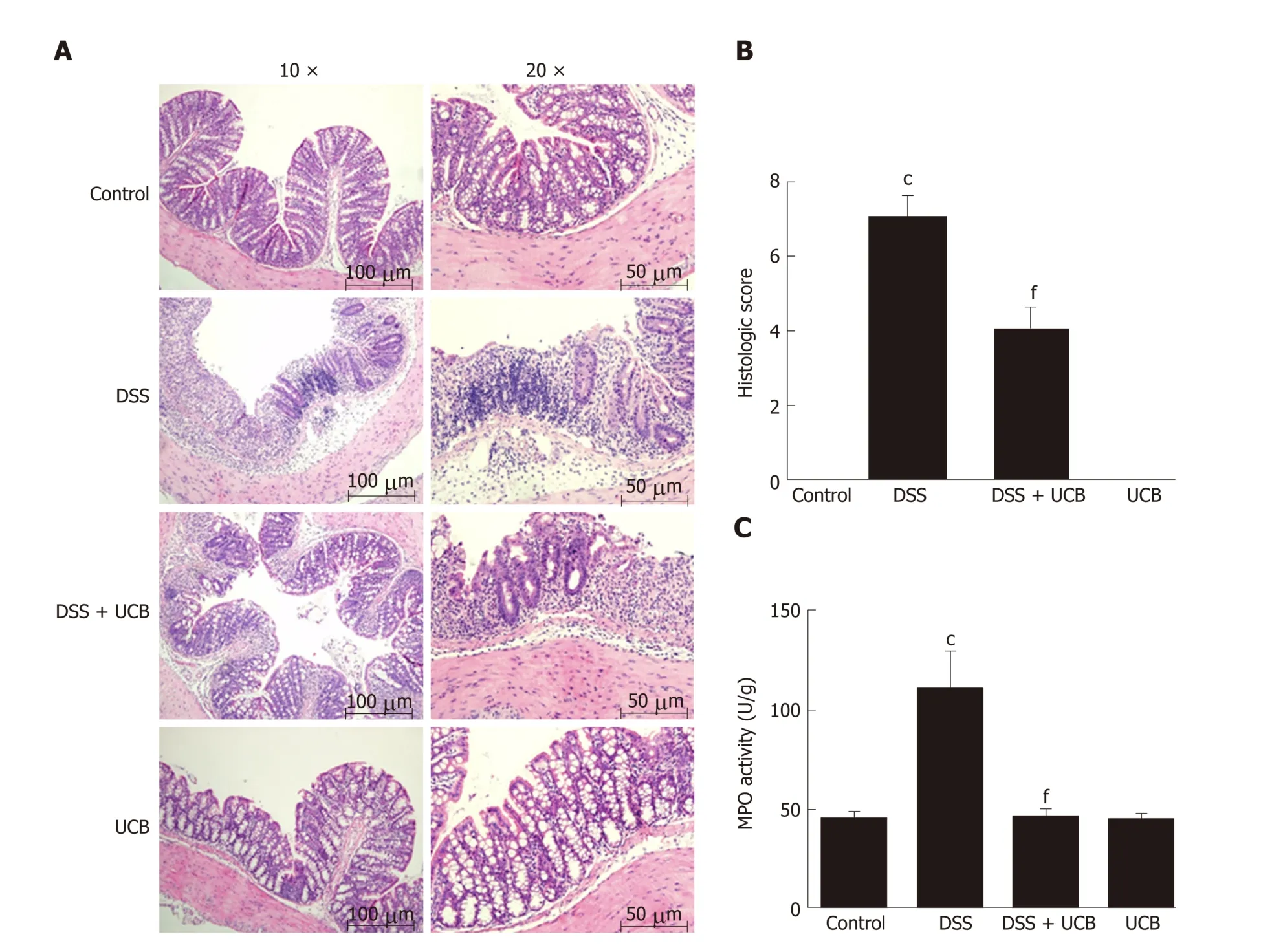
Figure 3 UCB ameliorated colon tissue injury in DSS induced colitis mice. A: Light microscopic assessment of hematoxylin and eosin stained sections; B:Histological scores (images acquired at 10 × using Olympus BX60; scale bar =100 μM; images acquired at 20 × using Olympus BX60, scale bar = 50 μM, n = 3); C:MPO activity of colon tissues (n = 5). Data are expressed as means ± SEM. c P < 0.001 vs control group, and f P < 0.001 vs DSS. UCB: Unconjugated bilirubin; DSS:Dextran sodium sulfate; MPO: Myeloperoxidase; SEM: Standard error of the mean.
UC was induced by 3% (w/v) DSS in drinking water for 6 d followed by untreated water for 2 d. Concurrently, colitis mice were administered 0.2 mL UCB (400 μM) by intra-gastric gavage for 7 d. Disease activity index (DAI) was monitored daily. The length of the colon and weight of the spleen were recorded. Serum level of D-lactic acid, intestinal digestive proteases activity, and changes in gut flora were analyzed. In add ition, colonic specimens were analyzed by histology and for expression of inflammatory markers and proteins.
Research results
UCB significantly relieved the severity of colitis, including lower DAI, longer colon length, and smaller spleen weight (P < 0.001). UCB inactivated digestive proteases (P< 0.01), increased expression of tight junction protein occludin (P < 0.001), decreased serum level of D-lactate (P < 0.001), and lowered histopathological score and activity of myeloperoxidase compared with those in colitis mice (P < 0.001). UCB also regulated the intestinal microbiota, inhibited expression of tumor necrosis factor(TNF)-α and interleukin-1β (P < 0.001), decreased expression of Toll-like receptor(TLR) 4 (P < 0.001) and myeloid differentiation primary response gene 88 (P < 0.05),and increased expression of TNF-receptor-associated factor 6 (P < 0.05) and IκBα (P <0.05) in the colon.
Research conclusions
UCB has a beneficial regulatory effect on intestinal barrier function and regulates normal intestinal homeostasis, and can suppress inflammation via the TLR4/NF-κB signaling pathway. This provides a theoretical basis for use of UCB as a clinical drug.
Research perspectives
UCB plays a pivotal role in intestinal innate immunity and inflammation. Thus, the find ings of this stud y ind icate a novel potential mechanism by w hich UCB can treat UC. More studies are needed to investigate the effect of UCB on chronic UC or colon cancer.
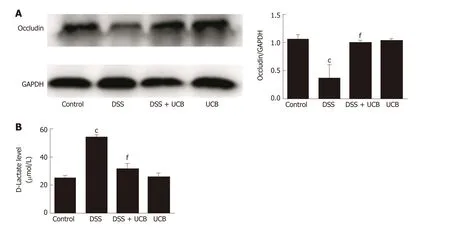
Figure 4 UCB increased expression of tight junction proteins in the colon and decreased intestinal permeability. A: Protein expression of tight junction occludin (n = 3); B: D-Lactate level in serum (n = 5). Data are expressed as means ± SEM. c P < 0.001 vs control group, and f P < 0.001 vs DSS group. UCB:Unconjugated bilirubin; DSS: Dextran sodium sulfate; SEM: Standard error of the mean.
ACKNOWLEDGMENTS
We acknow led ge Jin-An Zhou, Rong-Yan Li, and Yu Song for their excellent laboratory assistance.
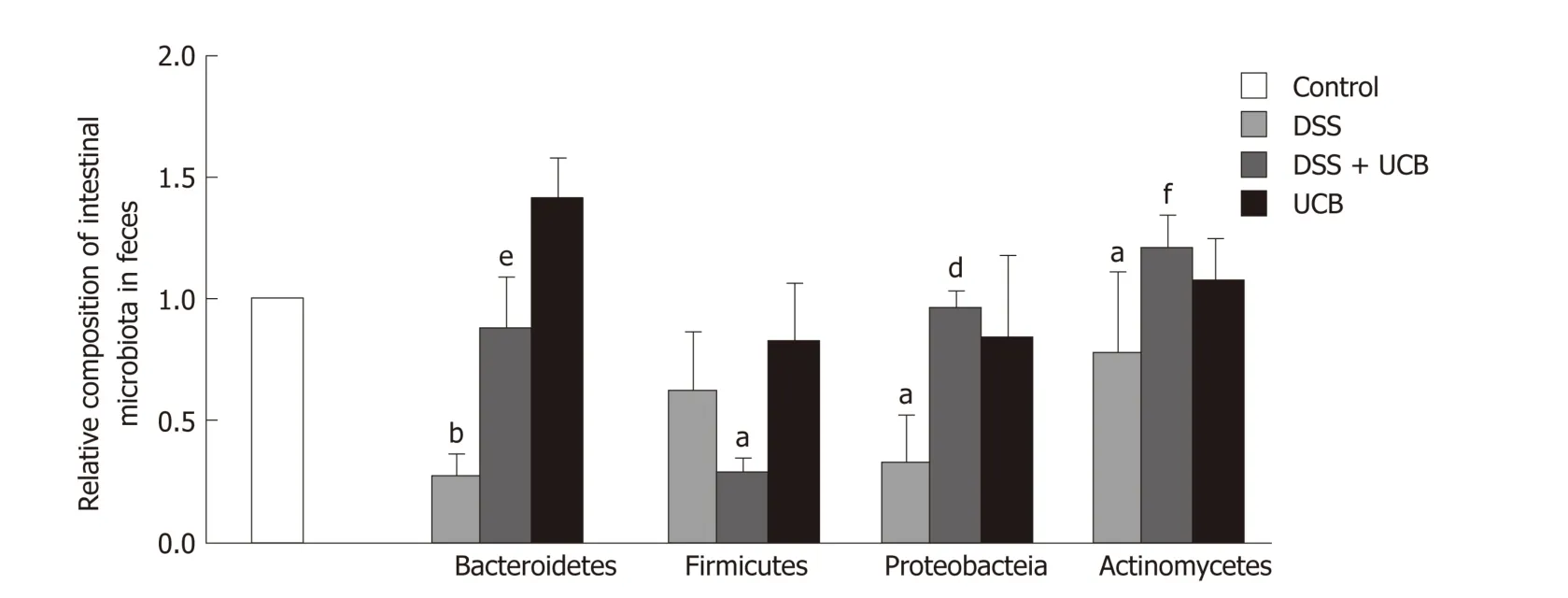
Figure 5 UCB maintained the steady state of intestinal flora of DSS induced colitis mice. Data are expressed as means ± SEM (n = 5). a P < 0.05 and b P < 0.01 vs control group; e P < 0.01, and f P < 0.001 vs dextran sodium sulfate group. UCB: Unconjugated bilirubin; DSS: Dextran sodium sulfate; SEM: Standard error of the mean.
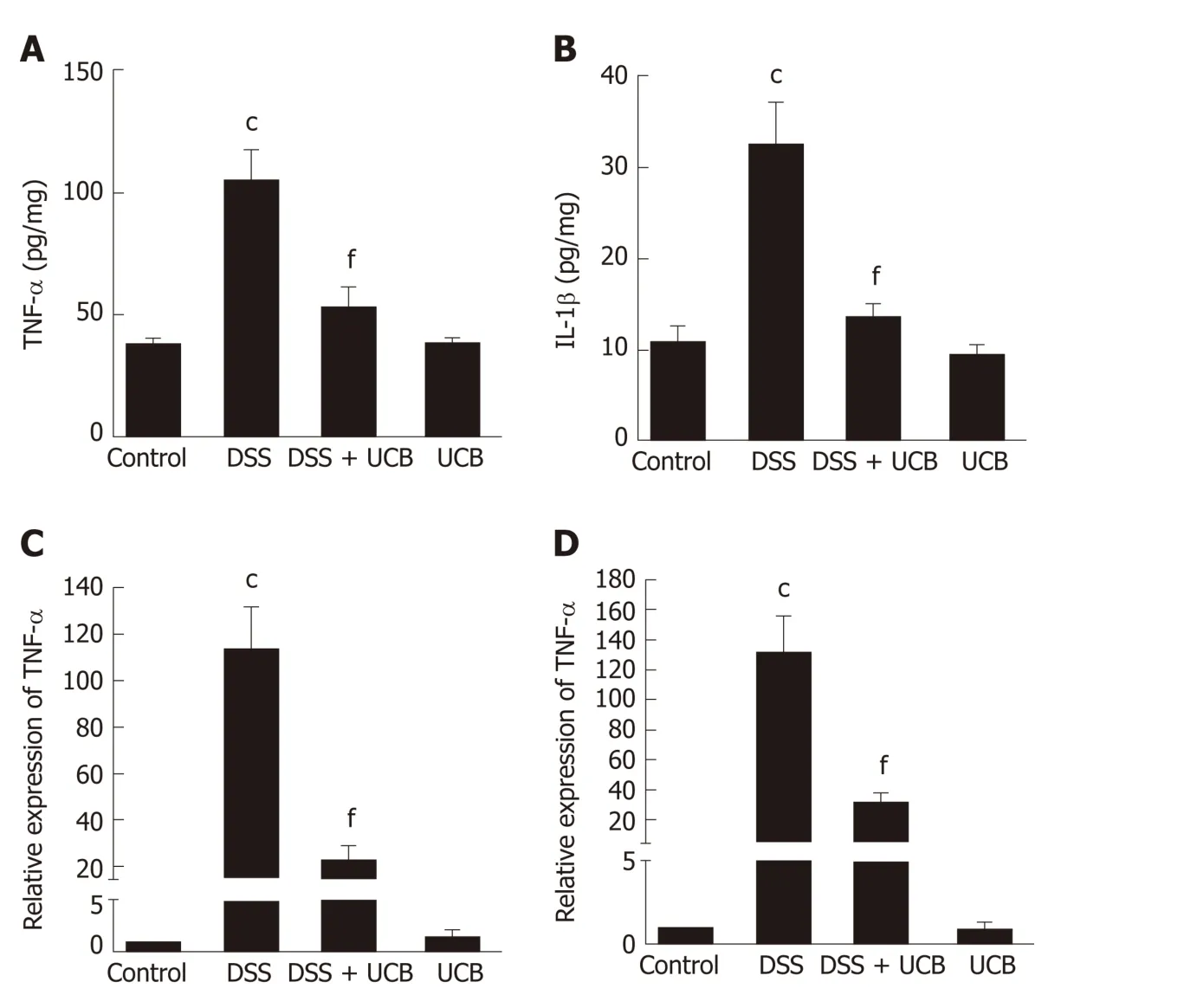
Figure 6 UCB reduced production of proinflammatory cytokines in DSS induced colitis mice. A and B: Protein expression of TNF-α and IL-1β; C and D: mRNA transcript expression of TNF-α and IL-1β. Data are expressed as means ± SEM (n = 5). c P < 0.001 vs control group and f P < 0.001 vs DSS. UCB: Unconjugated bilirubin; DSS: Dextran sodium sulfate; TNF-α: Tumor necrosis factor α; IL-1β: Interleukin 1β; SEM: Standard error of the mean.
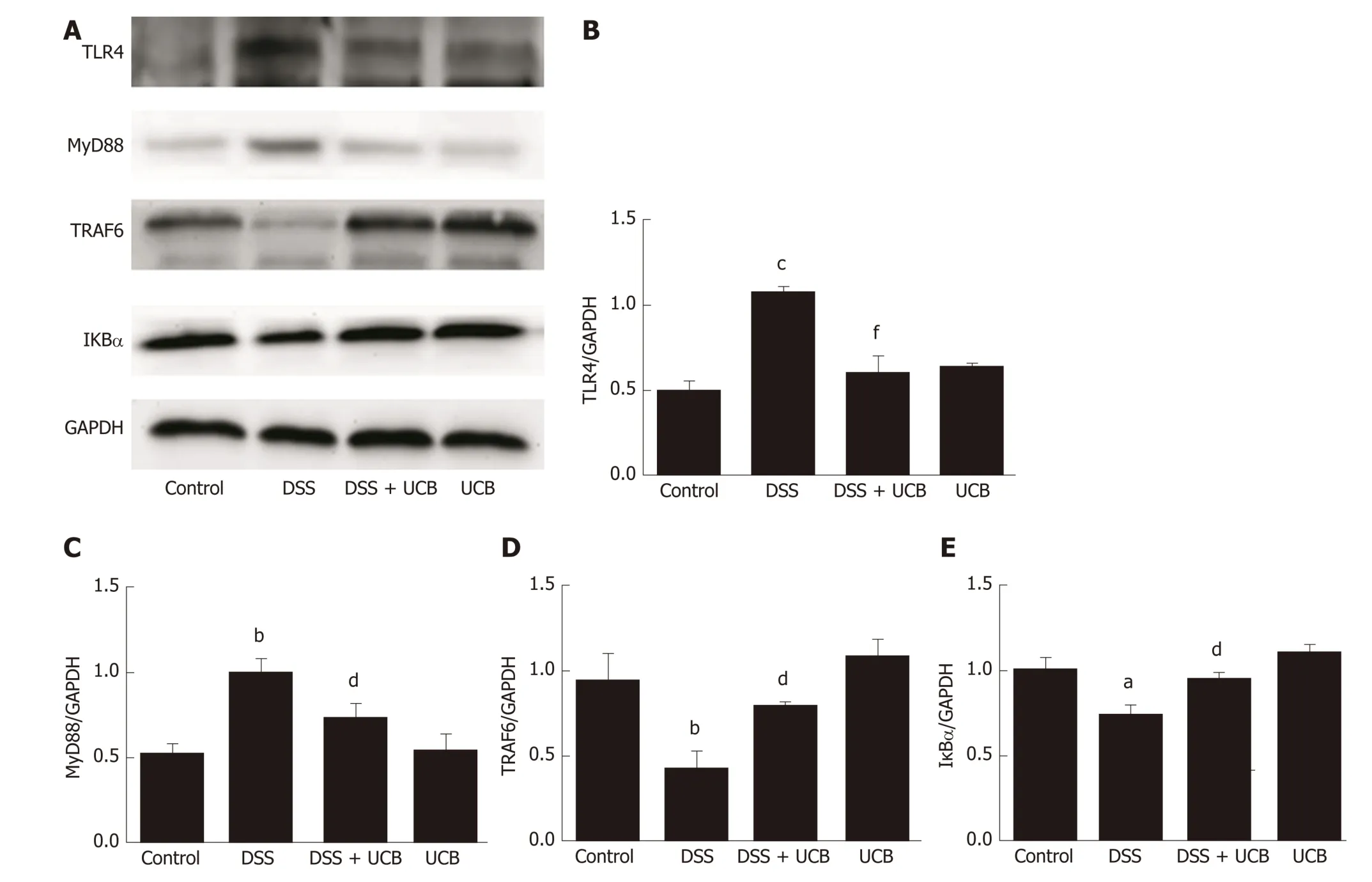
Figure 7 UCB inhibited the TLR4/MyD88/TRAF6/nuclear factor-κB signaling in DSS induced colitis mice. A: Total protein from colon samples was extracted,and TLR4, MyD88, TRAF6, and IκBα protein expression was measured by western blotting; B-E: Quantification of TLR4, MyD88, TRAF6, and IκBα protein expression was performed by densitometric analysis of the blots. Data are expressed as means ± SEM (n = 3). a P < 0.05 and c P < 0.001 vs control group; d P < 0.05 and f P <0.001 vs dextran sodium sulfate group. UCB: Unconjugated bilirubin; DSS: Dextran sodium sulfate; TLR4: Toll-like receptor 4; MyD88: Myeloid differentiation primary response gene 88; TRAF6: Tumor necrosis factor receptor-associated factor 6; IκBα: Anti-inhibitor of nuclear factor-κB alpha.
杂志排行

World Journal of Gastroenterology的其它文章
- Repurposing drugs to target nonalcoholic steatohepatitis
- Central role of Yes-associated protein and WW-domain-containing transcriptional co-activator with PDZ-binding motif in pancreatic cancer development
- Considerations of elderly factors to manage the complication of liver cirrhosis in elderly patients
- Lysyl oxidase and hypoxia-inducible factor 1α: biomarkers of gastric cancer
- Predictive and prognostic implications of 4E-BP1, Beclin-1, and LC3 for cetuximab treatment combined with chemotherapy in advanced colorectal cancer with wild-type KRAS: Analysis from real-world data
- Extract of Cycas revoluta Thunb. enhances the inhibitory effect of 5-f luorouracil on gastric cancer cells through the AKT-mTOR pathway