增温对冬小麦根系残体及秸秆分解特性的影响
2019-04-29王君陈书涛张婷婷刘仲旺殷梓絮王朝辉
王君 ,陈书涛 *,张婷婷 ,刘仲旺,殷梓絮,王朝辉
1. 南京信息工程大学/江苏省农业气象重点实验室,江苏 南京 210044;2. 南京信息工程大学应用气象学院,江苏 南京 210044
由于温室气体排放造成的全球变暖是当今倍受关注的全球性环境问题,据预测,到 21世纪末全球气温将上升 1.8-4.0 ℃(IPCC,2013)。全球变暖对生态系统碳循环具有潜在影响(Luo et al.,2008),农田生态系统是受人类活动强烈影响的系统,研究农田碳循环对气候变暖的响应规律和响应机制具有重要意义。在中国的农田生态系统中,秸秆还田是一种被广泛应用的保育土壤肥力的方法,作物残体分解不仅调节土壤碳转化和氮、磷营养的转运,也是向大气排放CO2的重要来源(Silver et al.,2001)。秸秆还田向土壤微生物提供了可供利用的碳源和营养,从而影响土壤 CO2排放(Thangarajan et al.,2013)。
已有研究较多关注于增温影响土壤呼吸、植物生长速率及碳氮含量等方面(An et al.,2005;Wan et al.,2007;Wu et al.,2011;Suseela et al.,2013;Cheng et al.,2017)。大多数研究表明,增温对土壤呼吸表现为正效应(Suseela et al.,2013;Xia et al.,2009),但在某些条件下,土壤呼吸也会对长期增温呈现出适应机制(Luo et al.,2001);增温提高了植物生长速率(Rustad et al.,2001;Wan et al.,2007;Wu et al.,2011);还有研究表明,增温处理下的植株氮含量提高,这会增加植物残体分解时归还到土壤中氮的量(An et al.,2005);但也有研究表明,增温通过降低总初级生产力和减弱生态系统呼吸进而造成净生态系统生产力下降(Li et al.,2017)。然而,以往关于长期昼夜增温处理的植物残体在下一阶段的分解系数的研究较为鲜见(Liu et al.,2017),特别是缺乏农田中作物残体经长期昼夜增温处理后在土壤中的分解系数及对土壤酶活性和可利用碳营养状况的研究(马欣等,2012;张旭博等,2011),而作物残体的这些分解特性对于了解农田土壤碳收支对气候变暖在多个生长季尺度上的综合响应规律是极为重要的(李小刚等,2002;王志明等,2003)。
作物残体经一定时间的增温处理后其自身性质(例如分解特性)可能会发生变化,而这种分解特性的变化可能与增温造成的作物本身碳氮含量的变化有关,同时作物残体性质的不同也可能会导致分解后土壤理化性质的变化。本研究采集经过一个生长季昼夜增温处理和对照条件下的冬小麦根系残体和秸秆样品,设计培养实验,研究不同处理下的根系残体和秸秆在土壤中的分解系数及残体分解后土壤中的酶活性、水溶性有机碳含量、pH等指标的变化情况,评估增温对冬小麦根系残体和秸秆分解特性的影响规律,以期为研究农田土壤碳收支对气候变暖的长期响应规律提供理论依据。
1 材料与方法
1.1 供试材料
于 2014年在南京信息工程大学农业气象试验站(32.16°N,118.86°E)设置了一个长期农田增温试验,种植方式为冬小麦(Triticum aestivum L.)-大豆(Glycine max L.)轮作。试验地属于北亚热带湿润气候,多年平均降水量和多年平均气温分别为1106 mm和15.4 ℃,年极端气温最高39.7 ℃,最低-13.1 ℃,年日照时数1902.5 h,无霜期237 d。试验地设置对照(不增温)和增温处理,采用红外辐射加热管对增温小区增温。增温小区中的增温装置为3个红外辐射加热管,加热管功率250 W,每天通过红外辐射加热管昼夜增温。本研究使用的有机物料为2017年5月23日收获的经一个生长季增温处理(即红外辐射加热管照射)和对照处理(即无红外辐射加热管照射,正常生长)的冬小麦根系残体和秸秆,该冬小麦种植于2016年11月6日,整个生长季增温处理比对照处理的温度高 2 ℃(Zhou et al.,2007;Suseela et al.,2012)。所收获的冬小麦根系残体和秸秆先磨碎,然后过2 mm筛,备用。供试根系残体及秸秆的碳氮含量如表1所示。在田间采集0-20 cm土壤,晾干,磨碎后过2 mm筛,备用。供试土壤有机碳含量19.4 g·kg-1,全氮含量 1.15 g·kg-1,容重 1.54 g·cm-1。
1.2 培养实验
通过培养实验研究经一个生长季增温处理后冬小麦根系残体和秸秆分解特性的变化情况。分别称量60 g过2 mm筛的土壤于490 mL广口培养瓶中,加水至25%的土壤质量含水量。将培养瓶置于25 ℃培养箱中预培养3 d。土壤预培养后,取增温处理的0.3、0.6、0.9、1.2 g根(标号为W-根)以及0.3、0.6、0.9、1.2 g秸秆(标号为W-秸秆),并取对照(不增温)处理的0.3、0.6、0.9、1.2 g根(标号为CK-根)以及0.3、0.6、0.9、1.2 g秸秆(标号为CK-秸秆),分别均匀平铺放入到装有60 g土的培养瓶中,每个处理设置3个重复。本研究参考了以往类似的培养实验的方法(Huang et al.,2004),将残体平铺撒到土壤上,由于培养瓶底面积为31.3 cm2,而最高的残体添加量仅为1.2 g,未观察到明显的残体堆积,而且在土壤水分适中的条件下,残体与秸秆的接触相对较大,也能进行较好地分解。同时设置不添加根系残体或秸秆,仅培养土壤的处理,标号为NR,该处理也是3个重复。培养实验所有处理的总数为 17个。整个培养实验过程中保持所有培养瓶的温度为25 ℃,含水量为25%。在培养试验期间为了使含水量保持为25%,每天采用称重法补水,即每天将装有土壤的培养瓶置于天平上称重,根据蒸发减少的水的质量,用滴管加入相应的水量。

表1 冬小麦残体的全碳和全氮含量Table 1 Carbon and nitrogen content of winter wheat residue
1.3 土壤CO2排放速率观测
在第 1、2、3、5、7、10、14、19、27 天采用红外CO2分析仪(Soilbox-FMS,M.C. Miller Inc.,USA)观测土壤 CO2排放速率。培养配上的瓶盖有2个铜制螺丝通气孔,通气孔通过两根乳胶管连接红外 CO2分析仪。瓶盖盖紧时,培养瓶内气体、红外CO2分析仪、乳胶管形成一个闭合回路,瓶内气体被泵入红外CO2分析仪内检测CO2浓度,检测频率为每秒1次,检测时长为3 min,由此可求出瓶内气体的变化速率,之后仪器可自动计算出单位土壤质量单位时间内的CO2排放量,即土壤CO2排放速率。
1.4 土壤pH、水溶性有机碳、酶活性测定
培养实验结束后,测定各个培养瓶中土壤的pH、水溶性有机碳(DOC)含量、脲酶活性、转化酶活性、过氧化氢酶活性。测定土壤pH的土水比为1∶2.5。以蒸馏水提取土壤DOC,再采用Mn(III)-焦磷酸比色法测定其含量(占新华等,2002;Giasson et al.,2014)。脲酶活性采用苯酚钠-次氯酸钠比色法进行测定(Gu et al.,2009),转化酶活性采用3, 5-二硝基水杨酸比色法测定(Gu et al.,2009),过氧化氢酶活性采用高锰酸钾滴定法进行测定(Stȩpniewska et al.,2009)。
1.5 数据分析
采用红外CO2分析仪测得的土壤CO2排放速率为单位质量土壤(g)在单位时间(d)内排放的CO2的质量(mg),采用内插法可根据每天的CO2排放速率计算不同处理下27 d内土壤累积CO2排放量。利用一元线性回归方法分析累积CO2排放量与秸秆添加量的关系,通过计算回归方程的斜率及标准误进而得到不同处理下根系残体及秸秆的分解系数,分解系数为单位质量(g)秸秆在单位质量(g)土壤中进行分解时所释放出的CO2量,以分解系数作为评估分解特性的指标之一。利用一元回归分析方法分析不同处理下累积CO2排放量与培养结束后土壤pH、DOC含量、酶活性的关系,以这些根系残体和秸秆分解后土壤理化性质的变化情况作为评估分解特性的参考。
2 结果
2.1 土壤累积CO2排放量及残体分解量
土壤累积CO2排放量随培养实验时间延长而逐渐增大(图1)。随冬小麦根系残体及秸秆添加量增大,处理27 d内土壤CO2累积排放量逐渐增大(图2)。不同残体添加量下的 CO2排放量存在显著(P<0.05)差异,添加根系残体和秸秆的土壤 CO2排放量无显著差异(P>0.05)。在相同的秸秆添加量条件下(如1.2 g),增温处理根系残体的土壤CO2排放量显著高于(P<0.05)对照根系残体的土壤CO2排放量;而对于相同的增温处理而言,添加根系残体的土壤CO2排放量与添加秸秆的土壤CO2排放量无显著差异(P>0.05)。土壤CO2排放量与残体添加量之间存在极显著的一元线性回归关系。线性方程的斜率即代表了不同残体的分解系数,表征了作物残体的分解特性。由图3可知,W-根的分解系数为(0.2699±0.0080)mg·g-1·g-1,高于 CK-根的分解系数(0.2407±0.0090)mg·g-1·g-1,对回归方程斜率(分解系数)进行t检验,结果表明,W-根和CK-根的分解系数达到显著差异水平(P<0.05);而 W-秸秆的分解系数为(0.2573±0.0030) mg·g-1·g-1,CK-秸秆的分解系数为(0.2587±0.0150) mg·g-1·g-1,t检验表明 W-秸秆和CK-秸秆的分解系数无显著差异(P>0.05)。此外,若将线性方程外推到秸秆添加量为0时,各处理的基础土壤CO2排放量外推值(即理论值)均小于观测得到的基础土壤CO2排放量,这表明秸秆添加对土壤原有有机碳的分解造成了负向的“激发效应”,即秸秆添加抑制了土壤自身含有的有机碳的分解。

图1 不同残体添加量下累积CO2排放量的时间变化Fig. 1 Temporal changes in cumulative CO2 emission under different addition amounts of residue

图2 不同残体添加量下的27 d累积CO2排放量Fig. 2 Cumulative CO2 emission during 27 days under different additive amounts of residue
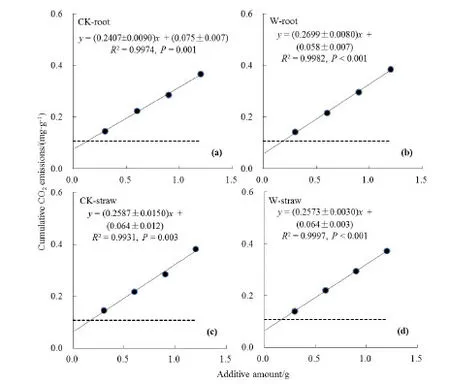
图3 土壤CO2排放量与残体添加量的线性回归关系Fig. 3 Linear regression relationship between soil CO2 emission and additive amount of residue
2.2 土壤CO2排放量与pH、DOC、酶活性的关系
2.2.1 土壤CO2排放量及残体分解系数与pH的关系

图4 土壤CO2排放量与土壤pH的关系Fig. 4 Relationship between soil CO2 emission and soil pH

图6 土壤CO2排放量与土壤DOC含量的关系Fig. 6 Relationship between soil CO2 emission and soil DOC content
回归分析表明,不同处理下土壤CO2排放量与土壤pH存在极显著(P<0.001)的自然对数回归关系(图4),前者随土壤pH的增大而降低,土壤pH可以解释 92.5%(R2=0.925)的土壤 CO2排放量的变异。随着残体添加量增加,土壤CO2排放量增大,培养后土壤pH相应降低,因而土壤pH的变化能解释土壤CO2排放量的变异。
计算4个残体添加水平的土壤CO2排放量平均值,同时计算4个残体添加水平对应的土壤pH值,然后将不同处理(CK-根、W-根、CK-秸秆、W-秸秆)的土壤CO2排放量与土壤pH进行回归分析,结果表明,两者间存在显著(P=0.010)的自然对数回归关系(图5a),土壤pH可解释94.7%的处理间土壤CO2排放量的变异,土壤CO2排放量随pH的增大而减小。不同处理下根系残体和秸秆的分解系数也与分解后的 pH存在一元线性回归关系(P=0.074)(图 5b)。
2.2.2 土壤CO2排放量与DOC的关系
如图6所示,不同处理下土壤CO2排放量与培养后的土壤DOC含量存在极显著(P<0.001)的自然对数回归关系,土壤CO2排放量随土壤DOC含量的增大而增大,土壤 DOC含量可以解释 87.3%(R2=0.873)的土壤CO2排放量的变异。一方面,随着残体添加量增加,培养后土壤中包含了更多的DOC,另一方面,秸秆添加量增加导致土壤 CO2排放量增大,综合这两方面的原因,土壤 DOC含量与土壤CO2排放量之间存在必然联系。
2.2.3 土壤CO2排放量与土壤脲酶、转化酶、过氧化氢酶活性的关系
与DOC类似,不同处理下土壤CO2排放量与培养后的土壤脲酶、转化酶、过氧化氢酶活性均存在极显著(P<0.001)的自然对数回归关系(图7),土壤脲酶、转化酶、过氧化氢酶活性分别可以解释75.7%(R2=0.757)、80.3%(R2=0.803)、92.7%(R2=0.927)的土壤CO2排放量的变异,其中,过氧化氢酶活性对土壤CO2排放量的解释性最好。

图5 4个残体添加水平的平均土壤CO2排放量和分解系数与平均土壤pH的关系Fig. 5 Relationship between averaged soil CO2 emission and averaged soil pH for four levels of residue addition and that between decomposition coefficient and averaged soil pH

图7 土壤CO2排放量与土壤脲酶、转化酶、过氧化氢酶活性的关系Fig. 7 Relationship between soil CO2 emission and soil urease, invertase and catalase activities
计算4个残体添加水平的土壤CO2排放量平均值,同时计算4个残体添加水平的土壤过氧化氢酶活性,然后将 4个不同处理(CK-根、W-根、CK-秸秆、W-秸秆)的土壤CO2排放量与土壤过氧化氢酶活性进行回归分析,结果表明,两者间存在显著(P=0.010)的线性回归关系(图8a),土壤CO2排放量随过氧化氢酶活性的增大而增大,土壤过氧化氢酶活性可解释99.0%的处理间土壤CO2排放量的变异。不同处理下根系残体和秸秆的分解系数也与分解后的平均过氧化氢酶活性存在一元线性回归关系(P=0.078)(图 8b)。
3 讨论
3.1 土壤 CO2排放及分解系数与残体添加量及类型的关系
本研究表明,土壤CO2排放量随冬小麦残体添加量的增大而增大,这与以往的研究一致(Thangarajan et al.,2013)。不同增温处理下的残体分解系数存在差异(图2),这与残体类型有关,增温处理后的根系残体具有更快的分解系数,而增温处理后的秸秆的分解系数与对照无显著差异(图3)。其可能的原因是田间增温导致土壤干旱(Liu et al.,2017),为获取水分,冬小麦根系更快地生长,根残体中的碳含量和氮含量也相应增加(表1),从而使增温处理后的根系残体在土壤中分解时可为微生物提供更多的碳氮营养,增温处理后根系残体全碳和全氮含量分别为(329.1±16.9) g·kg-1和(8.0±1.1) g·kg-1(表 1),显著高于对照,而增温处理后根系残体的碳氮比相对于对照而言更接近有机物料分解的最佳碳氮比 25∶1(Kumar et al.,2003),根系残体这些碳氮成分的变化促进了分解过程。
3.2 增温对残体分解特性的影响及机制
增温改变了根系残体和秸秆的分解特性,具体表现为其分解系数出现了差异,Meta分析表明,增温导致植物生长速率的加快,从而造成生物量的增加(Rustad et al.,2001;Dormann et al.,2002;Wu et al.,2011)。我们在其他田间试验中也观测到了增温导致冬小麦生物量的增加,生长速度的加快可能是导致增温条件下残体分解系数变化的原因之一。已有研究证明,温度较高地区的植物生长速度较高,生产力也较大,导致其分解速度也更快(Cornelissen et al.,1997;De Deyn et al.,2008)。

图8 4个残体添加水平的平均土壤CO2排放量和分解系数与平均土壤过氧化氢酶活性的关系Fig. 8 Relationship between averaged soil CO2 emission and averaged soil catalase activity for four levels of residue addition and that between decomposition coefficient and averaged soil catalase activity
增温与对照处理根系残体和秸秆的分解系数出现差异的机制可能既与作物的碳氮含量有关,也与分解过程中涉及的土壤理化性质的变化有关。其中,土壤pH可能是一个重要的指示指标,具体的过程为:不同增温处理下的根系残体和秸秆的分解速率产生了差异,而由于分解速率大的根系或秸秆导致土壤pH的下降也越明显,即相对更高的根系或秸秆分解量对应相对更低的土壤 pH(金婷,2017),因此本研究中不同增温处理下根系残体和秸秆的分解系数与土壤pH的负相关关系(图5b)在一定程度上说明了这种土壤pH对这些有机物料分解速率的指示作用。
除土壤pH外,培养后土壤其他理化性质的变化也可以在一定程度上解释不同增温处理和残体对土壤CO2排放量的影响规律。一方面,增温处理下分解速率大的根系残体和秸秆在分解过程中释放出更多的小分子含碳物质,导致土壤 DOC含量相应升高,另一方面,增温处理下分解速率大的根系残体和秸秆在分解过程中释放出更多的CO2,导致土壤CO2排放量的增大,因而土壤DOC含量的变化能较好地解释土壤CO2排放量的变异(图6),这与以往的研究结果类似(Huang et al.,2004;吴静等,2015)。土壤酶活性是表征土壤微生物活性和结构的重要指标(Bandick et al.,1999),生物大分子必须经胞外酶分解后才能进入微生物体内(Aerts,2006;Janssens et al.,2010),植物残体的分解是土壤微生物参与的一系列酶促反应(Cheng et al.,2012;黄继川等,2010;钱海燕等,2012;徐国伟等,2009),土壤酶活性一定程度上体现了有机底物和营养对微生物的满足程度(Caldwell,2005;Ling et al.,2010;吕国红等,2005;路文涛等,2011)。增温处理下分解速率大的根系残体和秸秆在分解过程中可为土壤中的微生物提供更多的营养,促使其释放出更多的酶并提高其酶活性,同时,根系残体和秸秆的分解会释放更多的CO2,虽然外来有机物料根系和秸秆会对土壤自身有机碳的分解具有一定的抑制效应(负向“激发效应”)(Kuzyakov,2002;Kuzyakov,2010),但根系和秸秆自身分解释放的CO2增量总体高于其对土壤有机碳负向“激发效应”的CO2减少量,因而导致土壤总 CO2排放量的增加,正是由于酶活性与土壤CO2排放量均受到不同增温处理下根系和秸秆添加的影响,其结果即表现为土壤CO2排放量与土壤的脲酶、转化酶、过氧化氢酶活性具有密切的相关性。
在进行本研究之前,已经测量得到田间增温和对照处理收获时的根残体总生物量分别为(149.4±16.7) g·m-2和(100.8±22.4) g·m-2,表明增温促进了根生物量的增加,增温造成根生物量增加的可能原因为:在水分较充足的条件下,增温使植物光合速率增大和生理活性增加(Wan et al.,2009;Dormann et al.,2002),且增温对光合的促进作用大于对呼吸的促进作用(Wan et al.,2009),有更多的光合产物会被分配到地下部分,从而导致根生物量增大。在本研究中,由于增温促使根残体的分解系数增大,分解加快。综合增温对田间根残体生物量的促进作用和对根分解速度的促进作用来看,增温导致的田间根残体生物量的增加效果可能会被根残体分解速度的增加作用所抵消。
增温对作物根系残体和秸秆分解量及分解系数的影响机制较为复杂,本研究仅对该问题进行了初步探讨,虽然本研究中测定的作物碳氮含量和土壤理化性质能部分解释其机制,但要更深刻地完全阐明相关机制今后还需要进一步研究。
4 结论
(1)添加残体后土壤的CO2排放量均随残体添加量的增加而呈线性增加。增温导致冬小麦根系残体的分解系数增大,但增温对秸秆的分解系数无显著影响。
(2)土壤CO2排放量与土壤pH,DOC含量,脲酶、转化酶、过氧化氢酶活性均存在显著的关系,土壤CO2排放量随着土壤pH的增大而线性减小,随着土壤 DOC含量的增大而呈对数形式增加,且随着土壤脲酶、转化酶、过氧化氢酶活性的增大而呈对数型增加。
(3)不同增温处理下根系和秸秆在土壤中的分解系数与土壤pH有关,分解系数随土壤pH的增大而减小,土壤pH是残体分解系数大小的重要指示因子。
