水稻叶绿素荧光特性对CO2浓度升高的代际响应研究
2019-04-29冯芳范佩佩刘超孙文娟于凌飞曹琰梅陈书涛胡正华
冯芳,范佩佩,刘超,孙文娟,于凌飞,曹琰梅,陈书涛,胡正华*
1. 南京信息工程大学气象灾害预报预警与评估协同创新中心/江苏省农业气象重点实验室,江苏 南京 210044;2. 中国科学院植物研究所/植被与环境变化国家重点实验室,北京 100093
随着工农业的发展,尤其是工业革命以来,煤炭、石油等化石燃料的大量使用,大气CO2浓度不断增加。工业革命前大气中的 CO2浓度为 280µmol·mol-1,而目前大气 CO2浓度已经增长到 409µmol·mol-1(https://www.CO2.earth/daily-CO2),21世纪末甚至达到 700 µmol·mol-1(Kump,2002;IPCC,2013)。
CO2是植物进行光合作用最重要的原料,CO2浓度升高会对植物的光合作用产生一定的影响(林伟宏,1998;Reddy et al.,2010)。植物叶绿素荧光可以反映光合能力的变化,特别是光系统Ⅱ(PSⅡ)的生理变化。当环境因子变化时,植物的叶绿素荧光变化在一定程度上反映环境因子对植物的影响(Maxwell et al.,2000)。通过对不同环境因子的快速叶绿素荧光诱导动力学曲线的分析,可以了解不同环境因子对光合机构的影响,尤其是光系统Ⅱ(PSⅡ)的影响和光合机构对环境的适应机制(李鹏民等,2005)。Ibaraki et al.(2005)研究表明,大气CO2浓度升高,低温条件下CO2浓度升高对水稻老叶的最大光化学效率Fv/Fm无影响,而高温条件下 CO2浓度升高对水稻老叶的 Fv/Fm有抑制作用。Hao et al.(2013)研究表明,大气CO2浓度升高对板蓝根(Isatis indigotica Fort.)叶片光系统Ⅱ的最大光化学效率Fv/Fm没有显著影响。王晨光等(2015)发现大气CO2浓度升高对大豆(Glycine max L.)叶片PSⅡ最大光化学效率、PSⅡ实际光化学量子产量和非化学淬灭系没有显著影响。张仟雨等(2016)则认为CO2浓度升高对大豆叶绿素荧光参数的影响因生育期的不同有所差异,大豆的最大光化学效率(Fv/Fm)在开花期无显著变化,在鼓粒期明显降低。王佩玲等(2011)研究表明 CO2浓度升高对冬小麦(Triticum aestivum L.)光合速率、叶绿素含量和光系统Ⅱ(PSⅡ)的光合电子传递和潜在活性具有一定抑制作用。CO2浓度对植物叶绿素荧光特性影响的研究大多是建立在 CO2浓度倍增或高CO2浓度下,且在连续几年试验中 CO2浓度保持不变(李清明等,2011;刘汉峰等,2012;石元豹等,2015;韩燕青等,2017)。叶绿素荧光技术是以植物的叶绿素荧光为探针,对植物本身没有损伤,荧光信号包含了光合作用过程的许多相关信息,因此被视为研究植物光合作用与环境因子关系的重要指标(Genty et al.,1989;Bolhar-Nordenkampf et al.,1989;Sayed,2003),对于探讨植物光合生理对大气 CO2浓度升高的响应有重要意义。目前关于CO2浓度升高对水稻叶绿素荧光特性的影响研究主要集中在一季水稻,而针对下一代水稻受CO2浓度升高影响,即对水稻叶绿素荧光代际影响的研究鲜有报道。
本试验以水稻(Oryza sativa L.)为研究对象,由开顶式气室(Open Top Chambers,OTC)组成的自动调控系统,设置不同 CO2浓度水平,使用上一季受到和没有受到 CO2浓度升高处理的粳稻种子育秧,对比分析两组种子的粳稻叶绿素荧光动力学参数对 CO2浓度升高的响应特征,研究常规稻的叶绿素荧光特性对 CO2浓度升高是否有遗传响应。研究结果对于探讨不同 CO2浓度升高条件下水稻的光合适应机制具有重要意义,可为提高农业应对气候变化能力提供基础性资料。
1 材料与方法
1.1 研究区域概况
试验于 2017年在南京信息工程大学农业气象与生态实验站(32.16ºN,118.86ºE)进行,该站位于江苏省南京市浦口区盘城镇落桥村,海拔约 22 m,属于亚热带季风气候,年平均气温为15.6 ℃,年平均降水量约为 1100 mm,平均日照时数大于1900 h,无霜期为237 d。土壤类型属于潴育型水稻土,灰马肝土属,耕作层土壤质地为壤质黏土,黏粒含量为 26.1%,0-20 cm 土壤容重为 1.57 g·cm-3,pH(H2O)值为 6.3,有机碳、全氮含量分别为 11.95 g·kg-1、1.19 g·kg-1,有效磷和速效钾分别为 6.89 mg·kg-1、62.8 mg·kg-1。田间轮作的作物为水稻和冬小麦,手工种植和收割,水稻收割留茬约10 cm,收获的秸秆不还田。
1.2 供试材料
供试水稻品种为粳稻9108,为常规粳稻,全生育期为149-153 d。有两组种子,一组是2016年水稻生长季种植在开顶式气室(OTC)内,并经CO2浓度升高处理(CO2浓度比自然环境高 40µmol·mol-1)收获的种子(SI),另一组是没有经过CO2浓度升高处理收获的种子(SII)。
两组种子均于2017年5月26日播种,6月20日移栽入OTC,移栽密度为30穴/m2。两组种子的秧苗在每个OTC内的移栽面积均为1.7 m2。全生育期的氮肥施用量为 17.6 g·m-2,复合肥(N∶P2O5∶K2O=15%∶15%∶15%)和尿素(有效成分占46.7%)配合施用,氮肥施用的比例为基肥∶分蘖肥∶穗肥=4∶3∶3。其他田间管理措施与当地大田生产一致,水稻主要生育期见表1。

表1 水稻主要生育期Table 1 Main growth stages of rice
1.3 试验设计
2017年水稻生长季试验设置3种CO2浓度水平: CK(以背景大气CO2浓度为对照)、CK+80µmol·mol-1(T1,在 CK 的 CO2浓度基础上增加80 µmol·mol-1)、CK+200 µmol·mol-1(T2,在 CK的 CO2浓度基础上增加 200 µmol·mol-1),每种处理有 4个重复。CO2浓度升高处理从水稻移栽至成熟。
农田CO2浓度自动调控平台由12个OTC、CO2传感器、供气系统和控制系统组成。OTC为正八边形菱柱体,框架由铝合金和高透光性的普通玻璃组成,高3 m,对边直径3.75 m,底面积约为10 m2。为聚拢气体并使气体散失速度放缓,顶部开口向内倾斜45°。在每个OTC内,距地面1.8 m处沿壁悬挂一圈输气软管,软管有均匀小孔,向 OTC内输送CO2,并通过鼓风机与风扇,促进OTC内的CO2均匀分布。OTC内CO2浓度采用自动控制系统进行快速反馈调节,通过接收气室内CO2传感器(型号GMM222,Vaisala Inc.,Helsinki,Finland)信号,根据实验设定浓度范围阈值,每天24 h连续供气,实时向气室补充CO2气体,使得OTC内CO2气体浓度达到目标浓度。OTC内安装有气温和相对湿度测定系统。
1.4 测定项目及方法
1.4.1 叶绿素荧光测定
使用 Handy-PEA便携式植物效率分析仪(Han-satech,UK)测定叶片叶绿素荧光特性。选择晴朗天气,分别在拔节期(8月10日)、扬花期(9月12日)、灌浆期(9月19日)、乳熟期(9月26日)、蜡熟期(10月6日)、完熟期(10月19日)进行测定。观测从9:00开始,每个OTC内任选5株长势均匀的SI和SII植株,用暗适应夹夹住剑叶进行暗反应20 min后,测定叶绿素基础荧光(Fo)与最大荧光(Fm)。根据Srivastava et al.(1997)的方法计算以下荧光参数:可变荧光(Fv)、PSⅡ最大光化学效率(Fv/Fm)、单位反应中心吸收的光能(ABS/RC)、单位反应中心捕获用于还原电子受体QA的能量(TRo/RC)、单位反应中心捕获用于电子传递的能量(ETo/RC)、单位反应中心以热能形式耗散的能量(DIo/RC)和单位面积光合机构含有的反应中心的数量(RC/CSo)。
1.4.2 产量测定
水稻收获时,分别收割不同处理下两组种子的水稻,脱粒后于 30 ℃下烘干称重,计算单面积产量即籽粒产量。
1.5 数据处理
运用Microsoft Excel 2010进行数据预处理;运用 SPSS 19.0统计软件对荧光参数进行ANOVA分析,采用LSD法对不同CO2浓度处理的叶绿素荧光参数进行差异显著性检验;运用 OriginPro 2016软件进行绘图。
2 结果与分析
2.1 叶绿素荧光诱导动力学参数的响应
2.1.1 基础荧光
图1所示为不同CO2浓度升高对水稻Fo的影响,由图可知,T1和 T2处理下,SI和 SII水稻的Fo在各生育期无显著差异。与CK相比,除拔节期和灌浆期外,T1和 T2处理使得 SI、SII水稻的Fo下降。其中,T1处理使乳熟期、蜡熟期和完熟期SI的Fo分别下降了 7.7%(P=0.000)、8.6%(P=0.004)、4.8%(P=0.001),灌浆期上升了5.7%(P=0.000);扬花期、乳熟期、蜡熟期和完熟期SII的Fo分别下降了17.7%(P=0.016)、5.2%(P=0.005)、8.0%(P=0.033)、4.2%(P=0.03),灌浆期上升了5.9%(P=0.002)。
与CK相比,T2处理下,扬花期、乳熟期、蜡熟期和完熟期SI的Fo分别下降了12.5%(P=0.033)、6.6%(P=0.000)、7.1%(P=0.014)、3.5%(P=0.015),灌浆期上升 5.9%(P=0.000);扬花期、乳熟期、蜡熟期SII的Fo分别下降了18.0%(P=0.015)、6.9%(P=0.000)、9.3%(P=0.014),在拔节期和灌浆期分别上升了66.4%(P=0.003)、3.0%(P=0.037)。

图1 不同CO2浓度水平下水稻Fo变化Fig. 1 Variation of Fo in rice under different CO2 concentration levels
2.1.2 最大荧光
Fm是 PSⅡ反应中心完全关闭时的荧光量,也表示为暗反应后的最大荧光强度(李鹏民等,2005)。由图 2可知,T1和 T2水平下,SI和 SII水稻的Fm在各生育期没有明显差异。与CK相比,乳熟期、蜡熟期和完熟期 T1和 T2处理使 SI、SII水稻的Fm上升。

图2 不同CO2浓度水平下水稻Fm变化Fig. 2 Variation of Fm in rice under different CO2 concentration levels
与CK相比,T1处理下,在乳熟期、蜡熟期和完熟期,SI的 Fm分别上升了 9.1%(P=0.000)、10.1%(P=0.001)、5.9%(P=0.011),SII的 Fm分别上升了7.1%(P=0.005)、11.0%(P=0.001)、6.5%(P=0.012)。
T2处理下,乳熟期和蜡熟期SI的Fm分别上升了11.7%(P=0.000)、12.0%(P=0.000);乳熟期、蜡熟期和完熟期 SII的 Fm分别上升了 9.4%(P=0.000)、10.6%(P=0.001)、5.6%(P=0.03)。2.1.3 可变荧光
Fv反映PSⅡ最初的电子受体QA的氧化还原状况(张其德等,2001)。由图3可知,T1和T2处理下,SI和SII水稻的Fv在各生育期均无显著差异。与CK相比,除拔节期和灌浆期外,T1和T2处理使得SI、SII水稻的Fv上升。

图3 不同CO2浓度水平下水稻Fv变化Fig. 3 Variation of Fv in rice under different CO2 concentration levels
与CK相比,T1处理使乳熟期、蜡熟期和完熟期SI的Fv分别上升了14.2%(P=0.000)、16.2%(P=0.001)、9.0%(P=0.002);扬花期、乳熟期、蜡熟期和完熟期 SII的 Fv分别上升了 10.8%(P=0.005)、10.8%(P=0.003)、17.7%(P=0.001)、9.7%(P=0.005)。
T2处理下,在乳熟期和蜡熟期,SI的Fv分别上升了17.2%(P=0.000)、18.2%(P=0.000);乳熟期、蜡熟期和完熟期SII的Fv分别上升了14.3%(P=0.000)、17.6%(P=0.001)、7.9%(P=0.02),拔节期下降了48.0%(P=0.007)。
2.2 PSⅡ最大光化学效率的响应
图4所示为不同CO2浓度升高对水稻Fv/Fm的影响,T1和T2处理下,SI和SII水稻的Fv/Fm在各个生育期均不存在显著差异。与CK相比,除拔节期和灌浆期外,T1和T2处理使SI、SII水稻的Fv/Fm上升。其中,T1处理使乳熟期、蜡熟期和完熟期SI的 Fv/Fm分别上升了 4.9%(P=0.000)、6.2%(P=0.001)、3.0%(P=0.000),灌浆期下降了1.4%(P=0.001);扬花期、乳熟期、蜡熟期和完熟期SII的 Fv/Fm分别上升了 7.1%(P=0.009)、3.7%(P=0.002)、6.5%(P=0.005)、3.1%(P=0.001)。
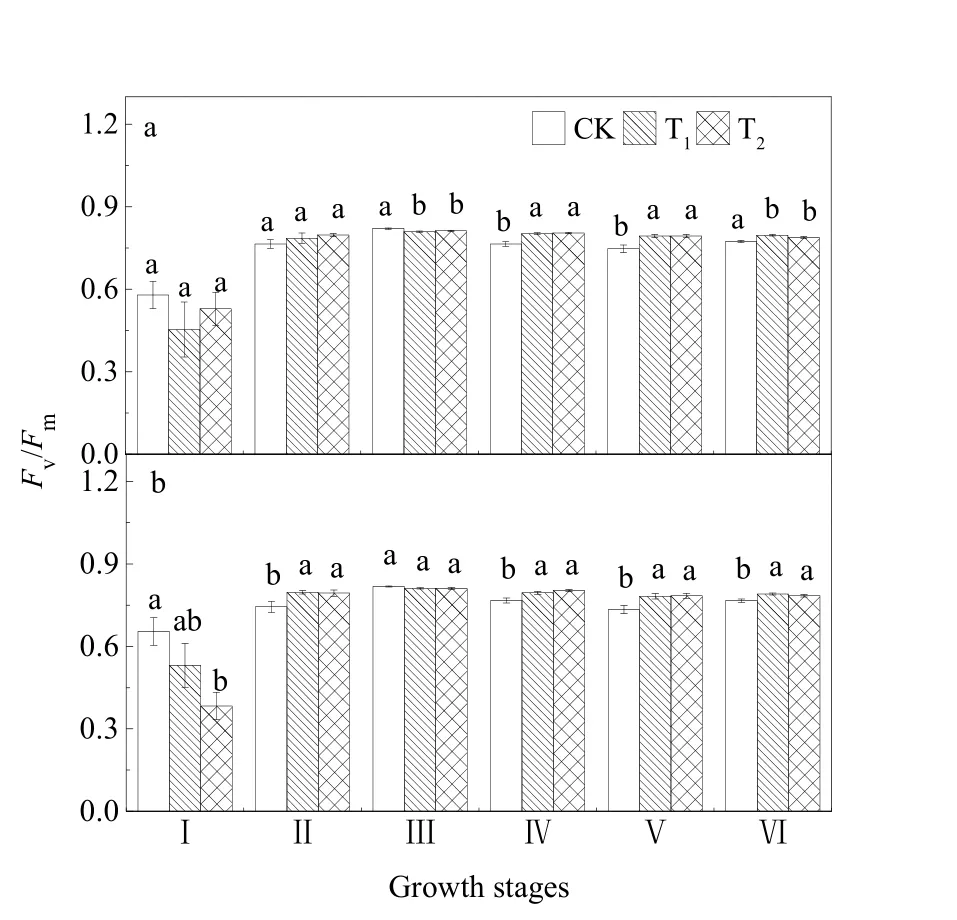
图4 不同CO2浓度水平下水稻Fv/Fm变化Fig. 4 Variation of Fv/Fm in rice under different CO2 concentration levels
与CK相比,T2处理下,乳熟期、蜡熟期和完熟期SI的Fv/Fm分别上升了5.3%(P=0.000)、6.2%(P=0.001)、1.9%(P=0.005);扬花期、乳熟期、蜡熟期和完熟期 SII的 Fv/Fm分别上升了 6.7%(P=0.013)、4.8%(P=0.000)、6.8%(P=0.003)、2.4%(P=0.012),拔节期下降了 41.4%(P=0.003)。
2.3 叶片PSⅡ反应中心比活参数和反应中心数量的响应
2.3.1 ABS/RC
由图5a和5b可知,与CK相比,T1处理下,乳熟期、蜡熟期和完熟期SI的ABS/RC极显著降低(P<0.01);扬花期、乳熟期、蜡熟期和完熟期SII的 ABS/RC极显著降低(P<0.01)。T2处理使乳熟期、蜡熟期和完熟期SI的ABS/RC极显著降低(P<0.01);扬花期、乳熟期、蜡熟期和完熟期SII的ABS/RC极显著降低(P<0.01)。
2.3.2 DIo/RC
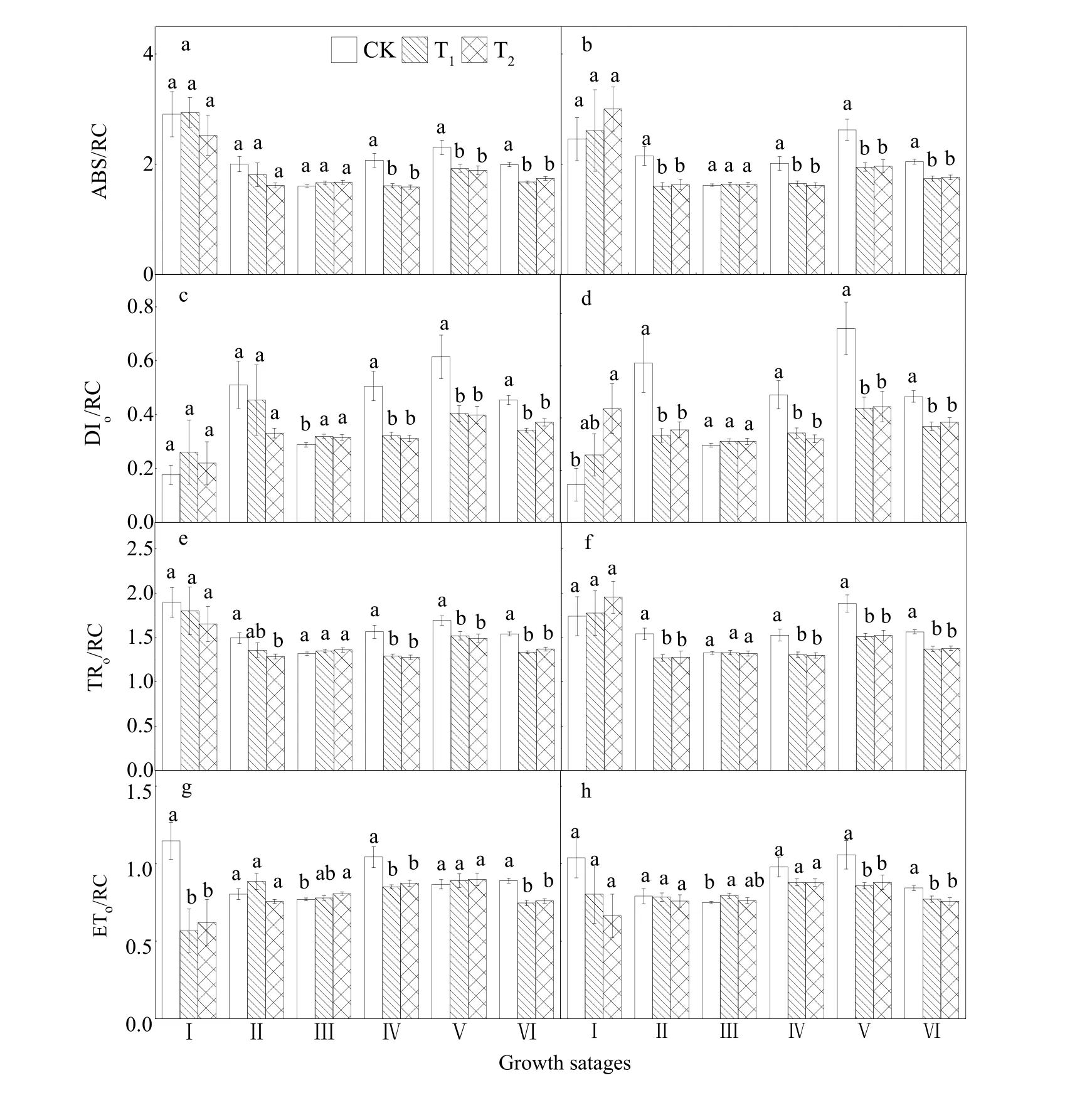
图5 不同CO2浓度水平下水稻PSⅡ反应中心比活参数的变化Fig. 5 Variation of activity parameters of PSⅡ reaction centers in rice under different CO2 concentration levels
由图5c和5d可知,与CK相比,T1处理下,乳熟期、蜡熟期和完熟期SI的DIo/RC极显著下降(P<0.01),灌浆期显著上升(P<0.05);扬花期、乳熟期、蜡熟期和完熟期SII的DIo/RC极显著下降(P<0.01)。T2处理使乳熟期、蜡熟期和完熟期 SI的DIo/RC极显著下降(P<0.01),灌浆期显著上升(P<0.05);扬花期、乳熟期、蜡熟期和完熟期 SII的DIo/RC显著下降(P<0.05)。
2.3.3 TRo/RC
由图5e和5f可知,与CK相比,T1处理下,乳熟期、蜡熟期和完熟期 SI的 TRo/RC显著下降(P<0.05);扬花期、乳熟期、蜡熟期和完熟期SII的TRo/RC显著下降(P<0.05)。T2处理使扬花期、乳熟期、蜡熟期和完熟期两类粳稻的TRo/RC显著下降(P<0.05)。
2.3.4 ETo/RC
由图5g和5h可知,与CK相比,T1处理使拔节期、乳熟期、完熟期 SI的 ETo/RC极显著下降(P<0.01);蜡熟期和完熟期SII的ETo/RC显著下降(P<0.05),灌浆期显著升高(P<0.05)。T2处理下,拔节期、乳熟期和完熟期SI的ETo/RC极显著下降(P<0.01),灌浆期显著升高(P<0.05);蜡熟期和完熟期SII的ETo/RC显著降低(P<0.05)。
在 T1和 T2处理下,SI与 SII水稻之间的ABS/RC、DIo/RC、TRo/RC、ETo/RC没有显著差异。2.3.5 RC/CSo
由表2可知,T1和T2水平下,SI和SII水稻的RC/CSo在各个生育期无显著差异。与CK相比,T1处理使乳熟期、蜡熟期和完熟期SI的RC/CSo分别上升了15.0%(P=0.000)、9.2%(P=0.012)、13.2%(P=0.000);扬花期、乳熟期、蜡熟期和完熟期SII的 RC/CSo分别上升了 10.5%(P=0.017)、12.7%(P=0.001)、19.0%(P=0.000)、13.3%(P=0.000)。

表2 不同CO2浓度水平下水稻的RC/CSo值Table 2 RC/CSo of rice under different CO2 concentration levels
T2处理下,在乳熟期、蜡熟期和完熟期,SI的 RC/CSo分别上升了 18.1%(P=0.000)、13.1%(P=0.000)、10.6%(P=0.000);SII的RC/CSo分别上升了12.9%(P=0.001)、18.4%(P=0.000)、13.7%(P=0.000)。
2.4 产量及其与叶绿素荧光参数的关系
不同CO2浓度升高对水稻产量的影响见表3。不同处理下 SI和 SII水稻产量均不存在显著性差异,不同CO2浓度下SI的产量表现为:T1>CK>T2,SII的产量表现为:T2>T1>CK。相关性分析表明(表4),T2处理下SI的产量与乳熟期的Fm、Fv、Fv/Fm呈显著负相关。CK处理下 SII的产量与扬花期的Fo呈显著正相关,与Fv和Fv/Fm呈显著负相关。T1处理下SII的产量与灌浆期的Fv/Fm呈显著负相关。T2处理下SII的产量与乳熟期的Fo呈显著负相关。
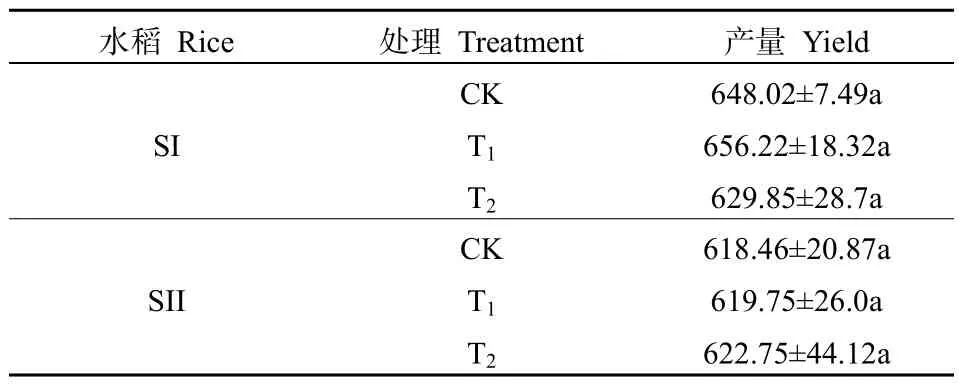
表3 不同CO2浓度水平下水稻产量的变化Table 3 Variation of rice yield under different CO2 concentration levels g·m-2
3 讨论
3.1 叶绿素荧光诱导动力学参数对 CO2浓度升高的响应
基础荧光被认为是 PSⅡ反应中心完全开放时的荧光,也表示为暗适应后的最小荧光强度(李鹏民等,2005)。杨广东等(2002)和许大全等(1992)认为PSⅡ天线色素的热耗散会导致Fo下降,但Fo的增加是由于PSⅡ反应中心的破坏或可逆失活。冯建灿等(2002)认为Fo的大小取决于PSⅡ天线色素的最初激子密度、天线色素之间以及天线色素到PSⅡ反应中心的激发能传递机率的结构状态。本研究表明,不同CO2浓度升高条件下,两组种子粳稻的Fo降低,这可能是大气 CO2浓度升高后粳稻的天线色素的热耗散增加导致的。最大荧光反映的是通过 PSⅡ的电子传递情况。可变荧光是Fm与Fo的差值,Fv随光合活性变化,它代表可参与 PSⅡ光化学反应的光能辐射(张其德等,1996)。本研究表明,CO2浓度增加 80 µmol·mol-1和 200µmol·mol-1,SI和 SII水稻的Fo、Fm和Fv在各生育期均无显著差异,说明CO2浓度升高对上一年经过CO2浓度升高处理的种子粳稻的基础荧光、最大荧光和可变荧光没有显著影响。不同CO2浓度升高条件下,两组种子粳稻的Fm、Fv上升,说明大气CO2浓度升高后,通过PSⅡ的电子传递能力和原初电子受体(QA)的还原能力有所上升,表明大气 CO2浓度升高促进水稻的光合作用。这也可能是由于叶绿素含量增加,叶绿素荧光发射能力提高,成熟期Fm越高,表明叶绿素受损程度越低,光合特性越好(李晶等,2011)。但郝兴宇等(2011)发现CO2浓度升高使绿豆(Vigna radiataL.)在鼓粒期的最大荧光(Fm)和可变荧光(Fv)下降,认为CO2浓度升高可能会导致PSⅡ反应中心的破坏,叶片光合能力下降。
3.2 PSⅡ最大光化学效率对CO2浓度升高的响应
Fv/Fm反映了PSⅡ反应中心的最大光化学效率或 PSⅡ原初光能转化效率,PSⅡ光合化学效率的提高有助于光合色素分子将捕获到的光能以更快的效率转化为化学能,为作物的碳同化提供充足的能量,从而有助于光合速率的提升(张其德等,2001)。Fv/Fm是反映在不同胁迫下植物光合作用受抑制程度的理想指标(张守仁,1999;Appenroth etal.,2001;Strasser et al.,2004)。本研究发现,CO2浓度增加 80 µmol·mol-1和 200 µmol·mol-1,SI和 SII水稻的Fv/Fm在各生育期均无显著差异,说明CO2浓度升高对上一年经过CO2浓度升高处理的种子粳稻的Fv/Fm无显著影响。不同CO2浓度升高条件下,两组种子粳稻的Fv/Fm上升。这说明粳稻叶片的PSⅡ原初光能转化效率提高,光合作用原初反应过程受到促进作用,这与张其德等(1996)的研究结果一致。由此表明CO2浓度升高有利于提高光合系统的光能转换能力,有效改善光合机构的功能。也有研究认为,CO2浓度倍增后冬小麦PSⅡ最大光能转换效率(Fv/Fm)值显著降低(王佩玲等,2011)。胡晓雪等(2015)研究表明,CO2浓度升高对野生大豆的Fv/Fm无显著影响。植物叶片的Fv/Fm对CO2浓度升高的响应因植物种类的不同而不同。CO2浓度升高对两组种子粳稻产量无显著影响,CO2浓度增加200 µmol·mol-1处理下,SI水稻的产量与乳熟期的Fm、Fv、Fv/Fm负相关,可能原因是粳稻后期叶片衰老,光合机构能力减弱,但具体的原因有待进一步研究。

表4 不同CO2浓度水平下水稻产量与叶绿素荧光参数的相关性Table 4 Correlation between rice yield and chlorophyll fluorescence parameters under different CO2 concentration
3.3 PSⅡ反应中心比活参数和反应中心数量对CO2浓度升高的响应
植物叶片天线色素吸收的光能除了小部分以荧光和热能的形式耗散外,绝大部分被PSⅡ反应中心捕获,在反应中心将QA还原成QA-,QA-又重新被氧化,从而产生电子传递,最终电子被用来固定CO2(Rai et al.,2008)。采用 JIP-test分析方法分析植物叶片光合机构的比活性,即单位反应中心的各种量子效率和单位面积反应中心的数量,比活性可以反映植物叶片的光合机构对光能的吸收、转化及耗散等状况(李鹏民等,2005)。ABS/RC和TRo/RC可以作为天线色素指标,这两个数值均增加,说明天线色素增大(Brestic et al.,2012)。本试验中,不同CO2浓度升高条件下,乳熟期、蜡熟期和完熟期两组种子粳稻的ABS/RC、TRo/RC、DIo/RC显著下降,完熟期ETo/RC显著下降。乳熟期、蜡熟期和完熟期两组种子粳稻的RC/CSo显著增加。说明CO2浓度升高虽然降低了粳稻叶片单位反应中心吸收的能量,但同时单位反应中心用于热耗散的能量也显著减少,导致单位反应中心用于还原 QA的能量和电子传递的能量显著减少。这可能是天线色素吸收的光能减少,或者是反应中心的活性受到抑制造成的。
4 结论
(1)在背景大气条件下,CO2浓度增加 80µmol·mol-1和 200 µmol·mol-1处理下,两组粳稻的Fo显著下降,Fm、Fv和Fv/Fm显著上升。
(2)在背景大气条件下,CO2浓度增加 80µmol·mol-1和 200 µmol·mol-1处理下,两组粳稻的ABS/RC、TRo/RC、DIo/RC和ETo/RC显著下降。
(3)两组粳稻相比,二者的 Fo、Fm、Fv、Fv/Fm、ABS/RC、TRo/RC、DIo/RC、RC/CSo、ETo/RC均无显著差异。表明在上一生长季经过CO2浓度升高处理和没有经过CO2浓度升高处理的粳稻,二者的叶绿素荧光参数对CO2浓度升高的响应没有差异,即两个生长季下来,粳稻叶绿素荧光特性对CO2浓度升高的响应没有表现出代际差异。
