兴安落叶松林球囊霉素相关土壤蛋白含量对年际间模拟氮沉降的响应
2019-04-29黄彬彬邢亚娟闫国永王庆贵
黄彬彬,邢亚娟,闫国永,王庆贵
黑龙江大学农业资源与环境学院,黑龙江 哈尔滨 150080
在全球范围内,由于人为来源氮(N)的排放的增加,导致大气N沉降在过去100年间增加了3-5倍(涂利华,2012;Gruber et al.,2008)。先前研究发现,N沉降可能改变土壤碳(C)库,使土壤碳源/汇发生变化(Zak et al.,2017;Chris et al.,2017)。然而,N沉降对土壤C库的影响主要是通过调节土壤有机碳(SOC)的形成和分解来实现的(Zak et al.,2017)。因此,了解N沉降背景下SOC的来源和分解对于认识气候变化驱动的土壤C循环具有重要意义。基于前人研究(Rillig et al.,2003b;Treseder et al.,2007a),一种由丛枝菌根真菌(AMF)产生的糖蛋白(球囊霉素相关土壤蛋白,GRSP)的发现,为了解SOC动态以及森林土壤C循环提供了一种新的认知途径。
GRSP是土壤活性有机C库中C最重要的来源之一(Comis,2004),不同地域每年可能沉积26-180 C·m-2(Rillig et al.,2003a)。研究表明,AMF与大约70%的植物家族形成了共生关系(Treseder et al.,2007b),并且在所有主要的陆生生物群落中都很丰富(Rillig,2004),这说明GRSP在陆地生态系统中普遍存在(Secilia et al.,1987)。GRSP在土壤中发挥着“超级胶”的作用,通过促进土壤团聚体的水稳定性间接保护不稳定SOC(Newman et al.,1987;Treseder et al.,2006)。GRSP 也可以通过刺激微生物的繁殖,从而影响微生物量的变化,使得微生物在加速分解土壤SOM的同时加速腐殖化过程,将不稳定C素组分转化为稳定的腐殖质,使活性C向稳定的有机C转变,间接起到稳定SOC库存的作用(Treseder et al.,2007b)。GRSP在土壤中的周转时间可以达到7-42 a之久,且随成土时间的增加具有逐渐升高的趋势(田慧等,2009;Rillig et al.,2003b;Staddon et al.,2004)。因此,土壤中 GRSP含量的变化,必将改变 SOC含量。
GRSP在土壤中的积累取决于许多因素,包括AMF丰富度、植物群落组成、土地利用系统和土壤理化性质等(Treseder et al.,2007a;Singh et al.,2016)。N沉降速率的增加可能对这些因素产生不同程度的影响。同时,N沉降将导致土壤有效N增加,并影响植物根系的形态结构、菌根侵染率和菌丝长度等(Ochoa-Hueso et al.,2016)。已有研究证实了 N沉降将会改变 AMF的结构组成(Jumpponen et al.,2005)。这些因素的变化都可能影响GRSP的产生,进而影响SOC含量(Schlesinger et al.,2000)。与此同时,N沉降会改变植物光合作用速率,进而影响其净初级生产力(net primary productivity,NPP)。而球囊霉门(Glomeromycota)真菌依赖植物的光合作用并利用植物相当大比例的同化C(大约4%-20%的净光合速率)(Jakobsen et al.,1990),NPP的大小直接影响着植物细根对C的分配(Kathleen et al.,2007b),从而影响菌根对GRSP的产生和C截获能力。已有研究发现N沉降也可能通过影响微生物量和活性及其群落结构,进而影响土壤GRSP的分解和储存。结合这些研究(Wang et al.,2018;Wang et al.,2017),认为 N沉降可能从GRSP的产生到分解等多方面影响着其在土壤中的含量,进而改变土壤C含量。
然而,目前从全球生态系统尺度研究增 N对GRSP的影响结果并不统一。Treseder et al.(2007)研究了3种北方森林生态系统GRSP对模拟N添加的响应,结果表明在幼龄次生林中减少,在中龄山杨(Populus davidiana)林中增加,在成熟黑云杉(Picea mariana)林中没有变化(Treseder et al.,2002)。目前,关于N沉降与GRSP之间的相关性关系尚未确定,这与当前缺乏对N沉降与GRSP之间相互关系的研究有很大关系。因此,N沉降如何影响GRSP的变化,还存在很大不确定性,尚需进一步深入研究。
由于GRSP的研究起步较晚,对N沉降背景下GRSP的驱动因素还鲜有报道。但是作为SOM重要的组成部分,研究GRSP对了解生态系统C动态具有重要意义。本研究旨在了解N沉降如何对GRSP产生影响。据此提出以下两个假设,(1)N沉降可能促进GRSP在土壤中的积累。当前大兴安岭地区森林生态系统中N素仍为一种限制因素,因此随着N沉降的增加,植物的光合速率将得到促进,会分配更多的C给AMF从而加速GRSP的产生。(2)GRSP可以作为一种研究 SOM 动态变化的“指示剂”,SOM作为土壤固相中最复杂的系统,如果能找到一种物质作为其动态变化的“指示剂”,可为研究森林生态系统C动态发现一种新的途径。
1 材料与方法
1.1 样地概况
试验样地位于黑龙江省大兴安岭松岭区的南瓮河国家级自然保护区(51°05′-51°39′N,125°07′-125°50′E),地处伊勒呼里山南麓。该区年均温为-2.4 ℃,属典型的寒温带大陆性气候。冬季受西伯利亚寒流影响,异常寒冷,最低温可达-48 ℃,夏季较短,最高气温可达36 ℃。最高月均气温(7月)约为18.6 ℃,最低月均温(1月)约为-26.3 ℃。年有效积温为1400-1600 ℃。早霜始于9月,晚霜至翌年5月中旬,无霜期为90-100 d。年平均降水量约为500 mm,其中80%出现在7-8月。受蒙古草原风的影响,年蒸发量一般在1000 mm左右,为降水量的2-2.5倍。
以兴安落叶松天然林为研究对象(表 1),该林分主要乔木层树种为兴安落叶松(Larix gmelinii),伴生有少量白桦(Betula platyphylla)、蒙古栎(Quercus mongolica)及毛赤杨(Alnus sibirica)幼苗;灌木主要有兴安杜鹃(Rhododendron dauricumL.)和杜香(Ledum palustravar.dilatatum)等;林下草本物种丰富,主要有禾本科(Gramineae)、菊科(Compositae)、豆科(Leguminosae)、蔷薇科(Rosaceae)、桔梗科(Campanulaceae)和鹿蹄草科(Phrolaceae)等植物。土层较薄,其中0-20 cm为砂质壤土,20-40 cm有较多碎石。
1.2 样地设计
N沉降控制样地始建于2011年5月,采用完全随机区设计,在立地条件完全相同的区域建立了3块大样地,每块样地被随机分为4个小样地,每个小样地面积为20 m×20 m。为了避免小样地间相互干扰,小样地间设置了10 m的缓冲带。中国北方自然N沉降量约为2.5 g·m-2·a-1,模拟N沉降量最好设置为当地实际N沉降量的2-3倍(Liu et al.,2013)。因此,共设置了 4个处理,分别为对照(Control,0 g·m-2·a-1)、低 N(LN,2.5 g·m-2·a-1),中 N(MN,5 g·m-2·a-1)和高 N 处理(HN,7.5 g·m-2·a-1)。4个处理随机分布在同一个大样地中,每个处理3次重复(n=3)。自2011年开始,在每年的生长季(5-9月)将每个处理每年将要添加的N量平均分配至每个月,然后均匀喷洒在样地表面。以硝酸铵(NH4NO3)作为外施N源,利用背式喷雾器进行人工喷洒,对照组喷洒等量的纯净水以消除水分差异。

表1 试验样地概况Table 1 Characteristics of experimental plots
使用内径为5 cm的土钻在每个小样地内进行随机6点取土,取土深度为0-10 cm,除去表层凋落物,将 6个点的土壤混合,装于自封袋中,带回实验室。
1.3 实验方法
将取回的土样进行风干处理,土壤C、N使用TOC/TN分析仪进行测定,土壤P采用钼锑抗比色法进行测定;SOM 采用稀释热-高锰酸钾容量法进行测定;GRSP测定参照Wright et al.(1996)的研究方法进行,实验前将考马斯亮蓝溶液过滤 2-3遍(刘晓蕾,2016);T-GRSP和EE-GRSP对SOM的贡献率(%)分别以 T-GRSP/SOM 和EE-GRSP/SOM的比值表示(Zhang et al.,2015);土壤温度和湿度利用LI-8100自带的TDR水分速测仪和温度传感器测定。
1.4 数据处理
运用Excel软件进行数据整理。采用SPSS 22.0中的单因素方差分析(ANOVA)检验 N沉降对GRSP、EE-GRSP和土壤理化性质的影响,采用Pearson相关分析法分析SOM与GRSP之间的相关性。
2 结果
2.1 不同施N处理对土壤C、N、P的影响
CK、LN、MN处理土壤全碳(TC)含量明显低于HN处理,其中2015年、2016年均表现出显著性差异(P<0.05),年际间没有表现出明显的变化趋势(表2)。土壤全氮(TN)在年内随着施N浓度不同表现出CK<LN<MN<HN,且各个年份的HN处理均显著高于其他 3个处理(P<0.05)。各个年份MN处理显著高于CK处理。年际间TN未表现出显著性差异,但随着年份增加具有明显升高的趋势。土壤全磷(TP)在年内除2016年CK处理明显高于其他处理(P<0.05),其余年份各处理水平均没有显著的差异,年际间未发现规律性变化。
2.2 不同施N处理对年际间SOM的影响
2015年,LN和MN处理SOM质量分数分别为(58.79±1.13) g·kg-1和(60.69±0.69) g·kg-1,显著高于 CK[(55.86±0.60) g·kg-1] 和 HN[(57.67±0.45)g·kg-1]处理(P<0.05)(图 1)。2016 年,LN 和MN处理SOM质量分数分别为(62.75±0.69) g·kg-1和(64.82±0.69) g·kg-1,显著高于 CK[(58.62±0.69)g·kg-1]与 HN [(59.99±0.79) g·kg-1]处理(P<0.05)(图1)。2017年,LN、MN处理SOM质量分数分别为(63.78±1.18)、(66.48±0.66) g·kg-1,显著高于 CK、HN处理(P<0.05)。
不同年份相同处理之间 SOM 含量只在 LN、MN之间表现出差异性,具体表现为2016年、2017年差异不明显,但均显著高于2015年(P<0.05)。
2.3 不同施N处理对年际间T-GRSP含量的影响
2015年,对照组(CK)与处理组(LN、MN、HN)T-GRSP质量分数分别为(2.45±0.15)、(3.08±0.04)、(3.53±0.10)和(2.68±1.11) g·kg-1。MN处理T-GRSP显著高于其他3个处理(P<0.05),LN处理显著高于CK、HN处理(P<0.05),CK、HN之间无明显差异。
2016年,LN和MN处理T-GRSP质量分数分别为(3.18±0.67) g·kg-1和(3.35±0.14) g·kg-1,显著高于 CK[(2.53±0.09) g·kg-1]和 HN[(2.60±0.04) g·kg-1]处理(P<0.05)(图 2B)。LN、MN 之间无明显差异,CK、HN之间无显著差异。
2017年,LN、MN处理T-GRSP含量无显著性差异,但均显著高于CK、HN处理(P<0.05),CK、HN处理之间无显著差异。对试验结果进一步分析发现(图2),除2016年、2017年MN处理显著高于2015年(P<0.05),其余各处理在年际间没有显著变化趋势。

表2 不同施N处理的土壤TC、TN和TP质量分数Table 2 The TC, TN and TP mass fraction under different N treatments g·kg-1

图1 不同施氮处理和不同年份土壤有机质质量分数Fig. 1 Mass fraction of soil organic matter in different nitrogen treatment and different years
2.4 不同施N处理对年际间EE-GRSP含量的影响
2015年,4个处理的土壤中EE-GRSP质量分数分别为(0.41±0.003)、(0.45±0.017)、(0.46±0.02)和(0.38±0.006) g·kg-1,LN、MN 处理无显著差异且显著高于CK、HN处理(P<0.05),CK处理显著高于HN处理。
2016年,LN和MN处理土壤中EE-GRSP质量 分 数 分 别 为(0.48±0.01) g·kg-1和(0.47±0.014)g·kg-1,无显著差异,且都显著高于 CK [(0.40±0.01)g·kg-1]和 HN [(0.37±0.006) g·kg-1](P<0.05)。
2017年,LN和MN处理土壤中EE-GRSP质量 分 数 分 别 为 (0.47±0.01) g·kg-1和 (0.47±0.01)g·kg-1,存在显著差异,且均显著高于 CK[(0.42±0.01) g·kg-1] 和 HN [(0.35±0.02) g·kg-1](P<0.05),而 CK处理亦显著高于 HN处理(P<0.05)。
对比相同处理不同年份之间 EE-GRSP的变化情况(图2A),不同年份4个处理的土壤中EE-GRSP变化情况均表现为 LN、MN显著高于 CK、HN(P<0.05)。LN、MN 处理表现为逐年递增趋势,HN处理则相反,CK处理没有明显的变化趋势。
2.5 GRSP与SOM之间的相关性

图2 不同施氮处理和不同年份EE-GRSP(A)/T-GRSP(B)质量分数Fig. 2 Mass fraction of EE-GRSP/T-GRSP in different nitrogen treatment and different years
对T-GRSP、EE-GRSP和SOM的含量对模拟N沉降的响应情况进行分析(图1和图2),发现三者之间有着相似的变化趋势,LN和MN处理土壤中,三者的含量得到了明显提升;HN处理则提高了T-GRSP含量,减少了EE-GRSP含量,但其幅度明显小于LN和MN处理。基于此,对三者进行了皮尔森(Pearson)相关性检验(图3),结果表明,随着T-GRSP与EE-GRSP浓度的升高,SOM含量呈线性增加(图3A、C)。随着EE-GRSP浓度增加,T-GRSP也呈线性增加(图3B),LN和MN处理提高了T-GRSP对SOM的贡献,HN处理则降低了T-GRSP对SOM的贡献(表2)。与对照组(CK)相比,LN和MN处理大多提高了T-GRSP对SOM的贡献(表2),只有2017年MN处理降低了其贡献率。与对照组(CK)相比,所有HN处理都降低了其贡献率(表2)。EE-GRSP对SOM的贡献表现为,和对照组(CK)相比,除2017年MN处理降低了EE-GRSP的贡献率(0.02%)之外,其他各组分均表现为促进(表3)。此外,不同年份施N处理EE-GRSP对T-GRSP的贡献率与对照组(CK)相比均表现为下降。
3 讨论
3.1 T-GRSP含量对N沉降的响应
GRSP通过直接影响土壤团聚体稳定性来影响土壤C库,此外,GRSP还提供了一个潜在重要的土壤C累积量,许多研究基于此进行GRSP对土壤C库贡献的研究(Zhang et al.,2015)。
本研究结果表明,T-GRSP对LN和MN处理都表现出积极的响应(图 2B),并且多数组分在P<0.05水平下表现为显著增加。HN处理的T-GRSP浓度与对照组(CK)相比与上述表现出相同的趋势,除HN处理,其他处理EE-GRSP与T-GRSP变化表现出一致性(图2A、B),这验证了本文提出的第一个假设。以下3个机制可能解释了低水平N可以促进GRSP增加的原因。
第一,LN和MN处理T-GRSP增加可能是连续N添加的累积效应(Zhang et al.,2015)。因为GRSP是由AMF菌丝释放,而AMF菌丝的平均停留时间(几天到几个月)通常比 GRSP(7-42年)短得多(Treseder et al.,2007b;Staddon et al.,2004);根据GRSP的产生机制:GRSP并不是被菌丝渗出,而是被包裹在菌丝壁内(Driver et al.,2005);所以当菌丝死亡或者分解时,GRSP才会被保留在土壤中(Treseder et al.,2000)。在LN和MN处理下,N素的添加减弱了其对AMF菌丝生长的限制作用,加速了菌丝的繁殖与更新,从而加速了 GRSP生成(Jakobsen et al.,1990),进而实现EE-GRSP含量增加和 T-GRSP的累积。此外,长白山针阔叶混交林中N沉降(50 kg·m-2·a-1)明显提高了AMF群落多样性和物种丰富度(郭伟等,2018);Eom et al.(1999)对连续9年模拟N沉降试验的土壤进行研究,发现AMF的生物量明显增加;Garcia et al.(2008)进行1年和2年模拟N沉降后发现N沉降会增加AMF的生物量,从而可能导致GRSP的增加。
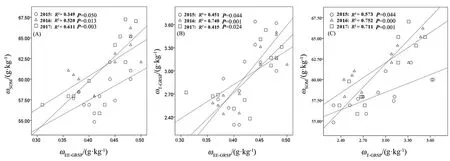
图3 EE-GRSP与T-GRSP和SOM的相关性(A、B)及T-GRSP与SOM之间的相关性(C)Fig. 3 Correlation between the concentration of E-GRSP with T-GRSP and SOC (A, B), and T-GRSP with SOM (C)n=12
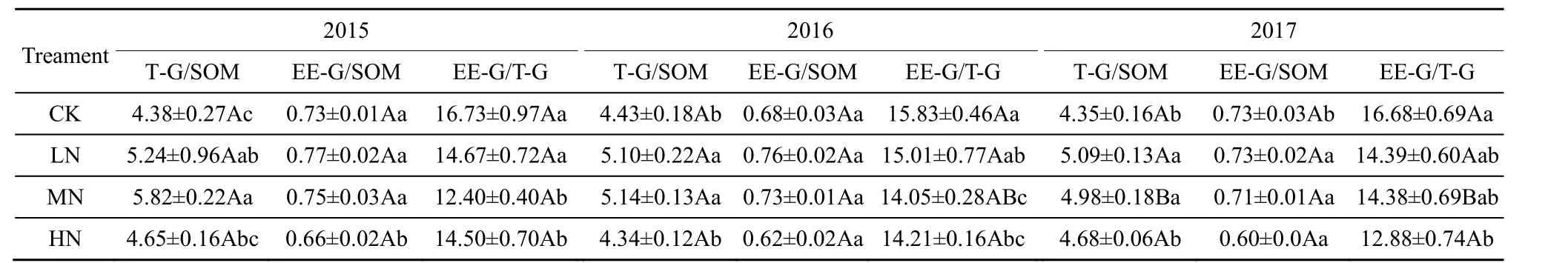
表3 GRSP对SOM的贡献率及EE-GRSP对T-GRSP的贡献率Table 3 Contribution rate of GRSP to SOM and the contribution rate of EE-GRSP to T-GRSP %
第二,虽然 T-GRSP被认为是一种难降解的GRSP,其在土壤中沉积的过程中仍然会受到土壤微生物的分解。Steinberg et al.(2003)和Rillig et al.(2003b)研究发现,黑暗处理150 d期间,来自温带森林土壤的GRSP下降了50%。另外,Wright et al.(1996)记录了在长达400 d的培养期间,俄亥俄州土壤中EE-GRSP和GRSP损失11%-57%。而且,微生物作为土壤养分循环的主要参与者,其对土壤环境的变化情况非常敏感。经过长期的模拟N沉降,土壤理化性质及各种元素之间的比例必定会受到外源N增加的影响,进而影响微生物活性,N添加导致土壤水分和土壤微生物活性下降(Wright et al.,1996;Chen et al.,2012;Mo et al.,2007)。因此,作为AMF的产物,GRSP可以在长期外部N添加下积累,表现出当前的N沉降速率对大兴安岭地区森林生态系统C库的积极影响。
第三,LN和MN处理的GRSP增多也可能是由于AMF获得的可利用C增多。N是大多数陆地植物生长的主要限制因子(王芳等,2017),而当N素作为限制因子时植物就会依赖AMF来促进自身对土壤养分的吸收(黄园园,2004)。研究表明,植物对AMF碳水化合物的投入小于植物通过AMF获得的养分收入。LN和MN处理可能在一定程度上缓解了 N限制,促进了植物净 NPP,使植物对AMF可用C的投入增加。一般而言,中等强度的N添加对植物的光合作用具有明显的促进作用,高强度的施 N量则会减弱这种促进效应甚至产生抑制效应(Bauer et al.,2004;鲁显楷等,2007;张蕊等,2013)。因此,LN和MN处理的GRSP增多可能是由于N沉降促进了植物光合作用速率,增加了NPP,进而促进了植物根系对AM可用C的投入。然而,由于北方针叶树种具有外生菌根(ECM),而GRSP则是由AMF产生。所以,在大兴安岭落叶松林中林下草本植物与灌木可能贡献了大部分的 GRSP,N沉降影响的可能主要是这一部分GRSP。因为相对于乔木,草本和灌木对N的变化可能表现得更加敏感。
结果显示,与对照组相比,HN处理EE-GRSP呈明显的下降趋势,而T-GRSP表现出上升趋势。首先,高水平N沉降可能使土壤养分含量过高,使植物可利用养分过剩,进而抑制了植物细根的生物量。此外,也减弱了植物对AMF的依赖,减少对其C的投入,降低了AMF的生物量,进而抑制了GRSP的产生(Jakobsen et al.,1990)。高强度N施用量也可能抑制了植物光合作用,减少NPP,从而对AMF产生负面影响,造成GRSP含量的下降。其次,HN水平下土壤化学性质的改变,如土壤酸化、地下资源减少,微生物活性在降低的同时其群落结构和物种丰度也发生了变化。因此T-GRSP含量的增加可能源于微生物对其的分解减少,以上因素可能是GRSP在HN水平下变化的一些原因。在LN和MN处理水平下,与EE-GRSP相比,T-GRSP增幅相对明显,这验证了施肥提高土壤 EE-GRSP与 T-GRSP的含量,但前者弱于后者的研究结论(Wright et al.,1998;杜介方,2010),说明在施N处理下T-GRSP的变化比EE-GRSP更加敏感。同时,T-GRSP浸提出的是土壤中全部的 GRSP,而EE-GRSP只是AMF新分泌的那一部分(Nichols et al.,2006),因此T-GRSP对于研究土壤中GRSP的状况更具代表性(杜介方,2010)。此外,基于先前的研究与本研究结论,认为EE-GRSP的浓度变化可作为AMF生物量变化的“指示剂”,其作为AMF所产生的一种糖蛋白,二者含量与生物量动态变化必定有着紧密联系。Lovelock et al.(2004)研究表明,菌丝长度可能不是AMF生物量的唯一指标,因此,菌丝的直径和侵染率等指标都是未来AMF与GRSP相关性研究需要加强的领域,以更明确地解释GRSP和AMF在N沉降背景下对森林SOC库的影响与贡献。
3.2 GRSP对SOM库的贡献
GRSP动态变化与 C动态联系在一起,GRSP中C以蛋白质和碳水化合物的形式存在且其含量丰富(Rillig et al.,2001),是土壤活性有机C库中C最重要的来源之一,其含量是过去认为土壤有机C最主要来源之一的腐殖质含量的2-24倍,是胡敏酸的 9倍(Rillig et al.,2001;Comis,2004;Nichols et al.,2006)。SOM为土壤C库的重要组成部分,无论是对土壤结构的稳定性,还是对地上植物的生长都具有重要意义(Hodge et al.,2001;Batjes,2014)。这种关系可以更好地预测GRSP对全球变化背景下生态系统反馈的贡献(Jakobsen et al.,1990)。本研究发现土壤中GRSP与SOM之间的动态变化具有惊人的一致性(图3),这与Wright et al.(1996)、Zhang et al.(2015)和杜介方(2010)的研究结果相同,同时验证了本研究的第二个假设,即GRSP可以作为土壤中SOC动态变化的“指示剂”。然而,与先前的研究相比,本研究中GRSP对SOM的贡献率相对较低,这可能与北方针叶树种具有外生菌根有关。在对内蒙古草原的研究中发现森林草原群落AM真菌的丰度显著高于其他草原群落(王启,2014),又因为其位于林下对光的接受能力弱等,这些因素都限制了GRSP在土壤中的积累。虽然GRSP与SOM表现出显著的相关性(图3C),但是由于与一些先前的研究相比 GRSP对SOM的贡献率(27%)相差甚远(表3),故本研究结论并不能作为一个代表性的结论去证明第二个假设。此外,本研究还发现EE-GRSP与SOM的相关性强于 T-GESP与 SOM(图 2A、图 2B)。EE-GRSP与一种免疫活性蛋白表现出与SOM更强的相关性(Lovelock et al.,2004;Wright et al.,1996),其具体机制目前尚不明晰(Treseder et al.,2006),可能原因是,SOM与GRSP之间具有一个复杂的相互影响机制,SOM含量同样会影响AMF对GRSP的生产量。这是因为SOC的储存影响着土壤的各种性状,如土壤孔隙度、土壤含水率以及土壤团聚体稳定性等(Wang et al.,2004;Lal,2005),这些性状影响着 AMF的生长和更新状况,与EE-GRSP的联系应该更加紧密。
综上所述,通过对实验结果的研究与分析,表明在当前N沉降持续加剧的背景下,GRSP呈现出累积的趋势,为森林生态系统C库做出积极贡献,加大了森林生态系统的C汇能力。同时,林下草本植物可能是大兴安岭落叶松林 GRSP的主要贡献者,虽然与其他不同林型对SOM贡献率相比较低,但在低水平N沉降速率下表现出逐年增加的趋势,可为将来研究林下草本植物对森林生态系统的C动态影响提供实验依据。
3.3 GRSP对年际间N沉降的响应
经过连续3年的GRSP含量测定,T-GRSP各项指标具有逐年升高的趋势,EE-GRSP在LN、MN处理中表现为逐年升高,HN处理表现为逐年降低;低水平N沉降处理所有指标都表现为促进。说明在当前阶段以及今后一段时间内的N沉降速率下,土壤中GRSP的累积量会逐年增加。因为低水平的N沉降将缓解森林生态系统的N限制,均衡了AM真菌的营养分配,使得土壤C汇能力逐年增加。土壤微域环境得到了很大改善,因为菌根真菌的菌丝是土壤中重要的稳定剂,在大部分土壤中根、菌丝和其他有机成分形成稳定的土壤团聚体(Schreiner et al.,1995;Piotroeski et al.,2004),此外,真菌的菌丝网络起着连接植物群落的功能。但是高水平N沉降(HN)则改变了土壤理化性质,使得土壤中养分过剩,影响了植物-真菌-细菌的养分分配策略,最终减少了GRSP在土壤中的累积量。与此同时,高水平 N沉降下微生物对土壤有机物的分解作用同样受到限制。两个过程对GRSP的作用相互抵消,最终可能间接地增加了GRSP的含量。
总之,通过对GRSP在年际间N沉降的变化情况的了解,加深了我们对未来 N沉降速率不断加快形势下北方森林土壤C库的动态变化的认知。
4 结论
(1)LN和MN处理增加了土壤中所有GRSP的含量,HN处理减少了 EE-GRSP含量但增加T-GRSP含量,表明低中水平 N处理加速了 AMF对GRSP的生产与释放,而HN处理则抑制GRSP在土壤中的消耗。
(2)EE-GRSP、T-GRSP与SOM具有显著的相关性(P<0.05),表明GRSP动态与土壤中C动态之间具有密切关系,但是尚不能得出GRSP可以作为SOM动态变化“指示剂”的假设的代表性结论。低中水平N处理增加了GRSP对SOM的贡献率,施N处理对EE-GRSP与SOM的比率虽有影响,但没有表现出规律性,说明低中水平 N处理增加了 GRSP含量,将促进森林生态系统 C截获能力,有助于缓解气候变化背景下大气CO2浓度增加的压力。
(3)通过对年际间GRSP对N沉降响应的研究,了解到GRSP对大气N沉降速率逐年增加表现出积极的响应,为研究森林生态系统 C库动态提供了一个重要依据。此外,在大兴安岭兴安落叶松森林中林下草本植物对于森林土壤C汇具有相当大的贡献。
