“所言非所指”:反语理解的脑机制及其影响因素
2019-04-01廖小根
廖小根
姜孟
四川外国语大学
【提 要】本文基于文献综述,梳理和分析反语理解的脑机制及其影响因素,并得出如下结论:1)左脑背外侧前额叶皮层(BA9)、左脑颞上回(BA21)、左脑颞中回(BA21/22)、左脑连接处(BA39/19)、左脑杏仁核(BA36)、左脑顶下小叶(BA39/40)、左脑额下回(BA44/45/47)等区域参与了反语的理解;2)反语理解有关的大脑基础与心理理论和语义整合两个大脑功能网络有着较高的重合性;3)不同形式非字面义语言理解的脑机制表现出一定程度的异质性,如反语和隐喻;4)反语理解脑机制的影响因素主要体现在主体因素的制约和客体因素的影响两个方面,前者主要包括心理理论能力、执行控制功能、认知策略,后者主要包括语境线索、反语类型和反语熟悉度。
1.引言
反语(irony)是日常生活中普遍存在的语言现象,它指使用与本意相反的语句表达本意。作为一种典型的非字面义语言(non-literal language),反语已受到修辞学、语言学(语用学、认知语言学和神经语言学)、心理学等诸多学科关注。从研究领域来看,反语研究主要关注反语的根本特征,反语的限制条件和引发机制,反语的语用价值(刘正光、崔刚2002),反语理解的心理、认知、神经机制及其影响因素。在反语研究的众多学科视角中,神经语言学算是一门相对稚嫩的学科。对反语理解的神经机制的探索肇始于二十世纪末期。随着认知神经科学的发展,该学科视角下的反语研究如雨后春笋般,日有所长。
在国外,近二十年来神经语言学领域对反语理解的研究正在如火如荼地进行。研究手段主要有fMRI(功能性磁共振成像)和ERP(事件相关电位);研究对象包括正常人群和病理人群,其中病理人群涉及精神分裂症患者(Herold et al 2018;Schnell et al 2016)和自闭症患者(Williams et al 2013)。从神经语言学的角度精细考察反语理解过程中的神经参与,重点在于回答:1)反语理解会激活哪些脑区或功能网络(Bosco et al.2017;Herold et al.2018;Obert et al.2016;Rapp et al.2010;Rapp,Mutschler&Erb 2012;Spotorno et al.2012)?2)在脑机制方面,反语较其它非字面义语言(如隐喻和习语)具有同质性还是异质性(Eviatar&Just 2006;Wakusawa et al.2007)?3)影响反语理解神经机制的因素有哪些(Baptista,Manfredi&Paulo 2017;Cornejo et al.2007;Filik et al.2014;Matsui et al.2016;Regel&Gunter 2017)?
然而,国内反语研究主要集中在对反语心理机制和认知机制的探讨。对反语理解的脑机制研究却近乎空白,只有零星的评介性文章。例如,张萌、张积家(2007)从脑损伤者和神经发展异常者(包括精神分裂症和孤独症患者)角度,评介反语认知神经心理机制的研究。马利军、张积家(2010)将隐喻、反语、谚语和惯用语置身于非字面义语言整个大框架下综述非字面义语言理解的脑机制研究。然而,从Eviatar&Just(2006)和Wakusawa et al.(2007)等研究来看,不同形式的非字面义语言之间存在质或量上的差异。因此,马利军、张积家(2010)的研究虽能够反映非字面义语言脑机制的一个全貌,但同时也可能会遮掩个体的差异。
为此,本文将反语同其它非字面义语言区分开来,梳理和分析国外有关反语理解的脑机制及其相关影响因素的研究成果,冀以此推动国内反语研究的发展,为探索反语理解的神经机制研究提供参考。
2.反语理解的脑机制
2.1 左脑颞顶连接处(BA39/19)
在语篇理解中,左脑颞顶连接处(temporoparietal junction,TPJ)主要负责根据语篇中的主人公信息进行心理化(mentalizing)或推理(infere-ncing)(Mason&Just 2009)。不仅如此,TPJ还在理解交际意图和社会意图归因中起着重要作用(Varga et al 2013)。在一项元分析研究中,Schurz et al.(2014)通过关联分析发现,无论实验采用何种刺激形式,所有心理化或推理任务均显著激活了TPJ。从理论上来说,在反语的理解过程中,受话者需要自动将心理状态归属他人,完成一个推理过程,以填补发话者字面义与实际义(交际意图)的空缺。最近的一些研究均发现反语理解过程中TPJ的激活(Bosco et al.2017;Herold et al.2018;Schnell et al.2016;Spotorno et al.2012;Varga et al.2013)。例如,Bosco et al.(2017)利用fMRI技术,结合反语故事理解任务,对比23个健康被试完成故事理解任务(故事目标句包括三种语用现象:字面义、反语和欺骗)的大脑激活情况。结果显示,反语交际意图的识别激活左侧颞顶连接处、左侧额下回、左侧额中回、左侧颞中回、左背外侧前额叶和右侧小脑。此外,Schnell等(2016)也通过fMRI技术,对比21名罹患精神分裂症的受试和24名健康被试理解反语的神经机制差异。他们同样发现,在反语任务的社会语境加工阶段,精神分裂症患者和健康控制组的TPJ均出现显著激活,但是前者激活的大脑神经网络范围更广,除TPJ外还激活额区、颞区和顶区。
2.2 左脑背外侧前额叶皮层(BA9)
一般认为,左脑背外侧前额叶皮层(dorsolateral prefrontal cortex,DLPFC)是负责执行控制功能的一个重要脑区(Leh,Petrides&Strafella 2010)。在反语言语行为的识别过程中,受话者必须消解发话者字面话语与其真正的交际意图之间的不一致(Bosco et al.2017)。因此,控制功能在反语理解中起着重要作用。具体来说,在理解反语时,执行控制功能的作用是同时加工多重意义,选择合适的意义;加工语境信息,抑制不合适的字面意义(马利军、张静宇、张积家2015)。左侧DLPFC参与反语理解已经为不少研究所证实(Bosco et al.2017;Herold et al.2018;Spotorno et al.2012)。在最近的一项研究中,Herold等(2018)对比12名精神分裂症患者、12名精神分裂症患者的一级亲属和12名正常被试在完成反语理解任务时两个阶段(语境加工和反语陈述理解)的差异。结果显示,在反语任务的语境加工阶段,亲属组左脑DLPFC的激活程度高于健康对照组。Herold et al.(2018)认为,在反语语境信息加工阶段,亲属组左脑DLPFC的激活程度高于健康对照组的原因可能与亲属组额外的认知努力有关,这可以理解为一种认知补偿活动,精神分裂症患者常表现出执行控制功能缺陷。Spotorno et al.(2012)也认为,左脑DLPFC出现显著激活是理解复杂的语言形式过程中高执行负荷的体现。此外,在Bosco et al.(2017)以正常人为被试的研究中同样发现了反语理解过程中左脑DLPFC的显著激活。当然,由于部分研究并未报道反语理解过程中左脑DLPFC的激活,因此它是否在反语理解中起着重要的作用,仍有待进一步考证。
2.3 左脑杏仁核(BA36)
Wang et al.(2006)较早发现反语理解过程中左脑杏仁核(amygdala)的激活。他们使用反语理解作为测试案例,考察儿童和成年人交际意图理解的神经机制。研究中,被试首先看卡通画,同时听短情景故事,故事结束后被试需要判断发话人说的是真实话语还是反语。他们通过fMRI技术扫描被试完成判断任务时的大脑后发现,与成年人相比,儿童理解反语时对内侧前额叶皮质和左侧额叶下回的激活程度更高,成人理解反语时梭状回、纹状体外区和杏仁核激活程度更高。后来,Uchiyama et al.(2012)采用fMRI技术对比正常被试理解反语和隐喻的神经机制差异。研究中,被试首先阅读短故事,然后根据故事提供的语境对目标句进行语义分类。结果发现,隐喻特有的激活出现在尾状核头部,它可能负责将陈述与可能的意义联系起来并将句子的意义限制在说话人想要表达的可能范围内。然而,反语理解过程中出现了左侧杏仁核的特异性激活。他们认为,该脑区是社会行为神经基础的重要组成部分。此外,Shibata et al.(2010)同样发现反语理解过程中大脑的杏仁核的激活。杏仁核还与社会认知功能有关,在情绪面部表情的识别和心理理论能力中扮演着重要的角色(Gibbard 2017)。过去研究发现,无法完成心理化任务的自闭症患者或阿斯伯格综合征患者一致表现出杏仁核结构和/或功能的异常(Li&Zhu 2015)。一方面,反语的理解需要受话者对社会语境和说话者的意图进行解码。因此,反语加工通常涉及到一种心理化过程。另一方面,无论是恭维反语抑或是批评反语,都涉及到不同效价的情感加工。在反语理解过程中,受话者对发话者的情感状态的推理是一个重要的过程。因此,反语加工过程中涉及到的非语言情绪线索的探测可能会激活杏仁核。
2.4 左脑颞上回、颞中回、额下回和顶下小叶
左脑颞中回(middle temporal gyrus,MTG,BA21/22)和左脑额下回(inferior frontal gyrus,IFG,BA 44/45/47)被视为语义网络的重要组成部分,它们在歧义的消解和依赖于语境的理解任务中起着至关重要的作用(Rodd,Gaskell&Marslen-Wilson 2002)。过去的研究发现,左脑MTG与左脑IFG共同负责控制语义提取、语用推理(Jang et al.2013)、语篇的连贯性分析和理解(Ferstl et al.2008)等多项任务。“其言非其义”是反语最为显著的特征,反语的理解是一个根据话语提供的信息或假设去寻找话语的最佳关联性的推理过程。由此观之,反语理解的整个过程与MTG主要的“职责”有较高的重合。从现有的神经影像研究来看,该脑区在反语的理解中确实也出现了激活(Bosco et al.2017;Eviatar&Just 2006;Uchiyama et al.2006;Varga et al.2013;Wakusawa et al.2007;Williams et al.2013)。例如,Eviatar&Adam Just(2006)采用核磁共振技术,要求被试阅读三句话组成语篇,其中关键句(语篇的最后一句)分别为字面义语句、隐喻句和反语句。研究结果显示,同字面义和反语理解相比,隐喻话语的理解显著激活了左脑额下回以及双侧颞下皮层。另外,同字面义理解相比,反语的理解导致颞上回和颞中回高度激活,隐喻的理解导致这些区域理解的中度激活。这表明,不同比喻性语言理解的半球敏感性存在差异。Uchiyama et al.(2006)使用场景阅读任务对20名正常被试进行fMRI研究。在该任务中,先呈现描述场景的句子,然后是主人公对此情景的评论。被试需根据情景,对评论的语义内容进行分类:讽刺、非讽刺或者语境上无联系。研究结果发现,分类过程(讽刺探测)显著激活左侧颞极、颞上沟、前额中皮层和额下回(BA/47)。讽刺探测激活左脑LFG的程度显著高于第一个句子理解。因此,左脑LFG可能是讽刺探测期间心理化和语言加工互动的神经基础。反语理解过程中左脑LFG的激活 还 在 Rapp et al.(2012)、Rapp et al.(2013)和Bosco et al.(2017)研究中有过报道。
此外,反语作为一种复杂语用现象,其理解是一个高级语言加工过程,这个过程可能会激活一个延申至额-颞和额-顶区域网络(Bosco et al.2017)。从相关研究来看,反语的理解过程还激活了左脑颞上回(superior temporal gyrus,STG,BA21)(Herold et al.2018;Obert et al.2016;Rapp et al.2012)。他们一致认为,颞上区域参与分析言语刺激以提取社会意义。左脑STG与左脑顶下小叶(inferior parietal lobule,IPL,BA39/40)共同负责反语理解的语义整合过程。其中,左脑IFG和左脑IPL的岛盖部被认为是人类镜像神经元(mirror neuron system,MNS)的组成部分(Bosco et al.2017)。也有研究发现,MNS参与非字面义语言的加工,这同样为反语理解加工过程中IFG和IPL的激活提供证据(Rapp et al.2010;Rapp et al.2012)。
综上所述,反语理解较为复杂,涉及的脑区较多,主要包括左脑颞顶连接处(TPJ,BA39/19)、左脑颞上回(STG,BA21)、颞中回(MTG,BA21/22)、左脑额下回(IFG,BA 44/45/47)、顶下小叶(IPL,39/40)、左脑杏仁核(amygdala,BA36)和左脑背外侧前额叶皮层(DLPFC,BA9)等区域(如图1所示)。这些脑区在反语的理解过程中扮演着特定的角色,当然,也有部分脑区的功能相似。其中,DLPFC、TPJ、IPL和杏仁核均为心理理论网络的重要组成成分(Spotorno et al.2012),而MTG、STG 和IFG等为语义整合脑功能网络的组成成分。因此,这两个脑功能网络可能在反语的理解过程中起着重要的作用。此外,从以上反语加工的脑区区域激活来看,支持反语加工“左脑优势”说。而且不同非字面义语言(如反语和隐喻)表现出一定程度的异质性。最后,有两篇文献(Bosco et al.2017;Rapp et al.2012)报道反语理解过程中右小脑的激活。当然,该脑区是否参与反语的理解过程还有待进一步考证。
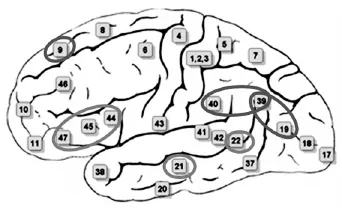
图1 反语理解激活的主要脑区分布
3.影响因素
近二十年来,国际学者围绕反语理解的神经机制开展了较为深入的探讨,然而在此问题上尚无定论。诚然,反语理解本身就受诸多因素影响,这使得其神经机制扑朔迷离。综合现有的相关研究来看,心理理论能力、执行控制功能、认知策略、语境线索、反语类型、熟悉度等均可能对反语理解的神经机制产生影响。本部分将从主、客体两个方面讨论反语理解的影响因素。
3.1 主体因素的制约
3.1.1 心理理论能力
心理理论(theory of mind,ToM)是指个体推测他人的心理状态并由此预测他人行为的能力(Premack&Woodruff 1978)。前面已经提到,反语的理解是一个较为复杂的过程,需要受话者对社会语境和说话者的意图进行解码。因此,心理理论(尤其是二阶心理理论)能力在反语的理解中起着重要作用。许多研究也发现了反语理解过程中心理理论网络典型区域(主要有DLSPFC、TPJ、IPL和杏仁核)的激活(Rapp et al.2013;Schnell et al.2016;Shibata et al.2010;Spotorno et al.2012;)。心理理论能力对反语理解的影响也能从病理人群相关研究中找到证据。例如,Gavilán&García-Albea(2011)通过两组被试(精神分裂症患者和健康受试)考察了语言理解能力与心理理论能力之间的关系。其中,两组受试心理理论能力存在明显的差异,但在人口统计学背景和神经心理行为学(智商、语言优势等)表现上高度匹配。他们发现,在各种比喻性语言的理解任务中,反语理解任务中的表现尤其与心理理论能力相关。此外,Bosco et al.(2018)以脑创伤患者为考察对象,探讨心理理论能力和各种交际行为的理解和产出能力之间的关系。结果发现,同健康对照组相比,脑创伤患者对语用现象(如反语交际行为、欺骗等)的理解和产出能力整体较差。他们认为,精神分裂症患者反语理解和产出能力较差的原因是其心理理论受损。
3.1.2 执行控制功能
执行控制,又称认知控制,指的是在认知资源有限的情况下根据目标和计划对当前任务的加工进行控制的能力,它主要表现在工作记忆、认知灵活性、抑制等方面(Miyake et al.2000)。它的存在保证个体能够以一种灵活的方式综合加工各类复杂的背景信息。理论上来说,在理解反语时,执行控制功能的作用是同时加工多重意义,选择合适的意义;加工语境信息,抑制不合适的字面意义(马利军、张静宇等2015)。此外,与心理理论能力相似,执行控制功能可能影响反语加工的神经机制主要是因为部分负责执行控制功能的大脑区域参与了反语的理解。例如,Spotorno et al.(2012)和Bosco et al.(2017)发现,反语加工过程显著激活了左脑背外侧前额叶皮层。他们认为反语理解是一个高执行需求的任务,执行功能在其中起着重要作用。过去的研究也发现,无论在语言认知任务和一般认知任务中,背外侧前额叶均参与控制认知加工的过程(Gilbert et al.2006;姜庆军、李勃等2013)。例如,在一项fMRI研究中,姜庆军等(2013)采用注意网络实验范式,对15名健康受试在任务态下进行功能磁共振扫描。结果显示,前扣带回、背外侧前额叶皮质等区域出现了显著激活。这表明,在该实验范式下,背外侧前额叶皮质参与执行功能。由此观之,执行控制功能与反语理解共享大脑神经基础。
3.1.3 认知策略
现有的研究表明,认知策略会影响认知加工,不同的认知策略与ERPs和神经影像探测到的大脑信号存在关联。反语理解的认知策略主要有整体性策略(holistic strategy)和分析性策略(analytic strategy)。前者主要从整体去考虑交际情景,而后者主要关注语句中的元素。Cornejo et al.(2007)借助ERP技术,考察不同理解策略对反语理解的影响。实验中,被试被要求对一组反语句子、字面义句子和没有意义的句子进行分类,这些句子均为故事的最后一句话。其中,一半被试判断句子是否有意义(整体性条件),另一半被试则判断是否与上下文一致(分析性策略)。结果显示,与使用分析性策略相比,被试在使用整体性策略诱发较大波幅的左侧额区和中央区负波(N400)。Cornejo et al.认为,被试使用整体性策略时缺乏语境信息,这使得他们在反语理解中出现N400类似的效应。因此,个体在理解反语时使用的策略会影响反语表达的感知与加工。当然,未来还需要神经影像研究的支持。
3.2 客体因素的影响
3.2.1 语境线索
语境在言语交际中扮演着重要的角色,反语理解与具体的语境相适应。直接通达理论(the direct access view)认为,语境信息在词汇激活的初期就会起作用,与语境一致的意义(非字面义)直接被提取,与语境不一致的意义(字面义)则不会被激活(Gibbs 1994)。因此,语境加工可以视为是反语理解的第一个阶段。语言交际过程中,诸如韵律、有关说话人语用风格和交际意图的相关知识等语境线索均可视为语境的一部分。近年来,语境线索对反语理解的影响也受到学者的关注。
在交际意图线索方面,Regel&Gunter(2017)利用事件电位相关技术,考察了线索性交际意图线索(通过给关键词添加引号实现)对反语和字面语言理解的影响。实验结果显示:同理解字面语言相比,被试在理解有线索的反语时诱发波幅为P300效应,该效应出现在刺激呈现200ms后。然而,在理解无线索的反语时诱发P200-P600双相模式。这表明,交际意图线索调制了反语理解的神经机制。
在说话者语用风格方面,过去的行为研究已经表明,听话人在理解不同职业背景的说话者的反语表达时表现出认知难度的差异。例如,与严肃的牧师相比,幽默的喜剧演员发出的反语更易为听话人所理解(Pexman&Olineck 2002)。可以说,越爱讽刺别人的说话者,其评论越容易被听话者理解为反语。为进一步证实交流风格对反语理解的影响,Regel、Coulson&Gunter(2010)借助 ERP技术,探讨有关说话者语用风格的知识对理解其言语反语的影响。研究中,为使被试获得有关说话者的语用风格的知识,整个实验(分两部分进行,间隔24小时)均对说话者反语使用的频率进行操控。在第一部分中,两个说话者使用反语的频率分别为70%和50%。在第二部分中,二者使用反语的频率均为50%。研究结果发现,被试在理解不常使用反语的说话者发出反语话语时诱发的N400波幅更大。然而,只有理解爱使用反语的说话者发出的反语时才诱发了波幅更大的P600,且当说话者语用风格与其使用的表达方式不一致时诱发较大波幅的P200。这表明,有关说话者语用风格的知识对其言语反语的神经机制会产生影响。
反语话语的感知依赖于多方面整合,主要包括听到的话语、话语的特定语境、非言语线索(韵律、语调)等(Woodland&Voyer 2011)。因此,韵律在反语理解中扮演着一定的角色。为揭示情感韵律对听觉模态反语理解神经机制的影响,Matsui et al.(2016)采用fMRI技术,对24个成人被试理解反语时的大脑进行扫描。实验流程包含以下几个阶段:第一阶段介绍相关背景(如A boy was playing with lots of toys.),第二阶段呈现父母对孩子说出来的话语(如His mother told him that he should tidy the toys before having his snack.),第三阶段呈现孩子对父母话语的反应(如The boy started eating his snack before tidying the toys.),第四阶段呈现父母对孩子反应的反语评论或者真诚评论(如His mother said,“You did a great job tidying the toys!”)。最后,被试判断父母的评论话语的情感韵律是积极的还是消极的。其中行为数据显示,积极韵律与积极语义内容结合时增强了话语的整体正性效价。相反,消极韵律与积极语义内容结合时减弱话语的整体正性效价。在神经基础方面,左侧喙中央额下回与上下文呈现出显著的交互效应,这表明不一致的韵律增强左侧额下回对依赖于语境的话语感知的神经反应。左侧额下回被认为是反语理解的神经基础之一(Rapp et al 2010;Spotorno et al.2012;Uchiyama et al.2006)。因此,情感韵律调制了听觉模态反语理解的神经机制。
3.2.2 反语的类型及其熟悉度
反语的分类有许多不同的维度。例如,根据话语的性质及语用功能,可将反语分为反语恭维(ironic compliment)和反语批评(ironic criticism)两类。Baptista et al.(2017)借助经颅直流电刺激和EEG技术,对比被试在理解反语恭维、反语批评、字面义恭维和字面义批评四种任务中的行为和脑电差异。行为结果和脑电结果一致显示,与反语批评相比,反语恭维的理解难度更大。一方面,这可能与两种反语在语言中分布有关,通常认为反语批评在成人言语中的分布频率高于反语恭维,这使得反语恭维意义加工的难度更大(Bosco et al.2017)。另一方面,在理解反语恭维时,受话人需要否定一个否定的陈述,以推断出说话人发出的反语恭维是积极的,理解反语批评过程中的这种双重否定会耗费受话者更多的认知资源(Pexman&Glenwright 2007)。
此外,分级显性意义假说(the graded salience hypothesis)认为,在非字面义语言认知加工中显性意义总是首先被激活(详见刘正光、崔刚2002)。显性(salience)意义是在心理词典中编码的意义,其显性程度主要取决于约定性、熟悉度、使用频率和典型性(Filik et al.2014)。该假说预测了熟悉和陌生反语加工的差异,主张熟悉反语理解通达不需要外在的推理加工,然而对于陌生反语,需要首先计算字面义,语境的失匹配将会导致反语陈述被再理解。Filik et al.(2014)采用眼动和ERPs技术,对比熟悉反语和陌生反语加工的差异。眼动实验结果发现,被试阅读到陌生反语中的无歧义词时眼动中断,阅读熟悉反语时无此现象;此外,同非反语相比,陌生反语关键词的凝视时间更长。ERP实验结果发现,同控制组相比,陌生反语理解过程中产生的N400波幅更大,但熟悉反语理解和控制组无差异。眼动数据和ERP数据均支持分级显性意义假说。由此观之,反语的熟悉度会影响反语的理解。
3.3 主客体因素的交互作用
前面已经从主客体两个维度阐述了影响反语理解的众多因素。那些影响因素之间是否存在交互作用?这是一个揭示反语理解神经机制的影响因素必须回答的问题。诚然,从收集到的文献来看,尚没有相关研究直接指向此问题。但有理由推测,某些影响因素之间会相互影响。例如,在上一部分已经提到,反语的类型及其熟悉度均可能影响反语的理解。但Baptista et al.(2017)发现,与反语批评相比,反语恭维的理解难度更大。反语恭维和反语批评作为两种不同类型的反语,大脑对二者的加工表现出差异。他们将部分原因归结为这两类反语在日常语言使用中频率分布差异所致。若此论断为真,那么对受话人而言,不同类型的反语与其熟悉度可能会相互影响。换言之,二者可能存在交互作用。
此外,尽管从概念解释来看,心理理论和执行功能似乎没有多大交叉关系,但二者的密切关系已经为越来越多的研究所证实。在一项最新的元分析研究中,Devine&Hughes(2014)收集了102项关于执行功能与心理理论能力的研究,共包含9994名3-6岁的儿童受试。其元分析结果显示,执行功能与心理理论间拥有15%的共同变异量,即使控制了年龄和言语能力的影响后,依然有8%的显著的共同变量,而且其研究结果具有跨文化的一致性。此外,在神经机制方面,执行功能与心理理论能力存在一定的重合性,如DLPFC均为二者的子机制之一。因此,二者之间很可能存在交互作用。
4.未来研究的展望
时下,国际上神经语言学视域下的反语研究正在如火如荼地进行,学者们借助不同研究手段,应用不同的研究方法和实验范式对反语理解的脑机制及其影响因素展开深入的探讨,业已取得可观的研究成果。然而,其发展与挑战并存。从目前显露的端倪来看,未来研究应关注以下几点:
第一,从个体功能定位上升到功能网络的探索。诚然,个体功能定位的方法有助于发现精细的结果。但从已有研究来看,反语加工这一复杂的认知过程并非由某个或某些脑区独立完成,而是多个脑区协同作用的结果。因此,从网络的角度探讨脑区之间的协同作用才能更全面地认识反语加工脑机制。通过文献梳理和分析发现,反语理解有关的大脑基础与心理理论和语义整合两个大脑功能网络有着较高的重合性。当然,此结论仍有待进一步考证。
第二,拓展反语理解神经机制的其它影响因素,并探讨各种因素之间的交互作用。尽管当前研究发现了诸多可能影响反语理解神经机制的因素。但尚有些研究结果相互矛盾。因此,一方面,要进一步考证部分已显露出来的影响因素;另一方面,应继续拓展影响因素。例如,实验任务、语言间差异、年龄等均有可能会调制反语理解的神经机制。此外,明晰各种影响因素之间的交互作用也应提上议事日程。
第三,细化反语类型,对比特定类别反语的神经机制差异。现有的相关研究普遍未对反语类型进行细分。然而,不同类型反语加工可能存在质或量的差异。这使得研究结果的可比性大打折扣。例如,恭维性反语与批评性反语虽同属反语的亚类,但二者通常会诱发不同的情感状态和情感属性,二者的脑机制很可能存在差异。因此,未来反语理解的神经机制研究在开展前应细化反语类型,将不同的反语亚类区分开来,去探索其相应的脑机制。
第四,转向对其它语种反语的关注。从近二十多年开展的相关研究来看,反语理解的神经机制研究对象主要集中在英语反语。然而,其研究结果是否存在跨文化的一致性?汉语与英语属于不同种语系,两种语言的反语理解可能表现出较大的差异。此外,国内反语研究目前处于一种“两头(认知机制和心理机制)大,中间(神经机制)小”的不均衡状态。因此,开展本土化(汉语反语)神经机制研究,迫在眉睫。
第五,深入讨论反语与其他形式或者说各种不同形式非字面语言的脑机制的异同。当前国内的研究多将隐喻、反语、谚语和惯用语置身于非字面义语言整个大框架下探讨其脑机制研究。然而,Eviatar&Just(2006)、Wakusawa et al.(2007)发现,不同的比喻性语言之间存在一定程度上的异质性。迄今,不同形式的非字面语言脑机制的对比研究匮乏,现有的研究主要将反语和隐喻对比。因此,各种不同形式的非字面语言脑机制的对比研究也是今后非字面义语言研究的一个方向。
