水稻耐淹成苗率相关性状全基因组的关联分析
2019-02-20孙凯李冬秀杨靖董骥驰严贤诚罗立新刘永柱肖武名王慧陈志强郭涛
孙凯,李冬秀,杨靖,董骥驰,严贤诚,罗立新,刘永柱,肖武名,王慧,陈志强,郭涛
水稻耐淹成苗率相关性状全基因组的关联分析
孙凯,李冬秀,杨靖,董骥驰,严贤诚,罗立新,刘永柱,肖武名,王慧,陈志强,郭涛
(华南农业大学国家植物航天育种工程技术研究中心,广州 510642)
【背景】耐淹成苗率低是限制直播稻产量的重要因素,挖掘高种子活力、低氧萌发能力强的水稻材料是提高耐淹成苗率的关键,但控制耐淹成苗率的遗传位点的挖掘仍然比较有限。【目的】利用来源广泛的自然种质,分析影响耐淹成苗率的关键表型性状,挖掘相关的遗传位点和候选基因,为直播稻耐淹成苗机理研究提供一定的理论和材料基础。【方法】以200份来源广泛的水稻种质为材料,在有氧环境下进行发芽试验,测量种子发芽率、发芽指数和活力指数;在低氧条件下测量芽鞘长和芽鞘直径;进行耐淹成苗试验,水深10 cm,20 d后测量耐淹成苗率。分析各性状间的相关性,挖掘影响耐淹成苗率的关键性状;利用简化基因组测序对以上6个表型进行全基因组关联分析,鉴定与性状显著关联的SNP位点,并在关联区间内筛选候选基因;对02428和YZX 2份材料进行有氧、无氧以及氧气含量转换条件下的转录组检测,结合全基因组关联分析结果,分析候选基因的表达模式差异。【结果】种子活力、芽鞘表型和耐淹成苗率在200份材料间存在广泛的遗传变异,其中,芽鞘长和活力指数的变异系数最大;相关分析结果表明,芽鞘长、活力指数与耐淹成苗率呈极显著正相关;通过全基因组关联分析,共鉴定出8个与活力指数显著关联的位点,15个与芽鞘长显著关联的位点;结合基因组注释,在关联区间筛选出6个与活力指数相关的候选基因,7个与芽鞘长度相关的候选基因;进一步比较13个基因在有氧、无氧及氧气转换条件下的表达模式以及表达量的变化,发现、、表达量变化显著,表现出对氧气处理的敏感性。【结论】种子活力、芽鞘长与耐淹成苗率密切相关,可作为筛选高耐淹成苗水稻材料的重要性状。全基因组关联分析、转录组分析与基因表达模式比较的联合应用可提高候选基因的筛选效率。水稻耐淹成苗过程可能受到与逆境胁迫、光合作用相关基因的调控。
水稻;种子活力;耐低氧萌发;全基因组关联分析;转录组分析
0 引言
【研究意义】水稻是世界上最重要的粮食作物,全世界近1/2的人口以稻米为主食。水稻也是中国最主要的粮食作物,中国2/3的人口以稻米为主粮,水稻播种面积占国内粮食种植面积的27%左右[1]。在水稻各种生产方式中,直播技术由于具有轻型、高效、节水、省工的优点,正逐渐成为水稻轻简高效栽培的理想方式。直播技术分水直播、湿直播和旱直播3种形式,其中,湿直播与旱直播成苗率较高,但田间操作难度大,且容易受到草害、鸟害和虫害影响,而水直播是直接将种子撒播于浅水层覆盖的土壤中,田间操作难度低,杂草危害轻,易于为农民接受,在中国水稻直播生产中得到了广泛应用。然而,水直播的种子萌发时处于淹水条件下,受到低氧胁迫,对品种的耐淹成苗能力具有较高要求,不适合直播的品种通常耐淹成苗能力差,田间秧苗群体不均匀,进而影响直播产量。水稻的胚芽鞘是极少数可以在缺氧条件下生长的植物组织之一[2]。缺氧环境促进水稻胚芽鞘的伸长,抑制幼茎和幼根的生长。水稻胚芽鞘通过快速伸长到达有氧环境,为胚芽和胚根的生长提供氧气。在氧气供应下,水稻种子胚芽和胚根的生长很大程度上受到种子活力的影响,种子活力越高,幼苗生长越快,抗逆性越强,田间群体表现越整齐。因此,水稻耐淹成苗能力受到胚芽鞘性状和种子活力性状的共同控制。在解析水稻耐淹成苗能力与胚芽鞘性状、种子活力性状关系的基础上,进一步发掘与耐淹成苗性状有关的遗传位点并获取分子标记,对于培育适合直播的水稻品种具有重要的意义。【前人研究进展】水稻耐淹萌发的遗传机制比较复杂,已有的研究表明不同水稻品种的耐淹萌发能力存在显著差异,耐缺氧的品种比不耐缺氧的品种有着更长的芽鞘[3-4],表明这一性状受到了遗传位点的控制。陈孙禄等[5]通过对粳稻80380与籼稻RP2334的高代回交自交系连锁分析,发现了4个控制耐淹萌发的QTL。Septiningsih等[6]以淹水发芽21 d的存活率为指标检测到6个QTL,其中位于第7染色体的位点得到了另一个群体的验证,说明其控制耐淹萌发的真实性。芽鞘的生长与中胚轴的发育密切相关,Lee等[7]采取5 cm的深播胁迫,用Kasalath/Nip构建98个回交重组自交系对水稻幼苗中胚轴长度进行QTL定位,发现在第1、3、6染色体有3个关于中胚轴伸长的QTL,依次为、和,其中和的贡献率分别为12.89%和8.28%,的贡献率在2个独立重复中分别为37.56%和26.97%。已有的报道大部分基于双亲分离群体(如RIL、NIL、BIL等)进行遗传位点挖掘,受限于双亲遗传差异或分子标记密度,所能鉴定的耐淹萌发位点比较有限。全基因组关联分析(genome-wide association study,GWAS)以自然群体长期重组交换保留下来的位点间连锁不平衡(linkage disequilibrium,LD)为基础,通过检测表型数据和基因型数据的有效关联来寻找遗传位点[8-9],对于复杂数量性状位点的挖掘具有突出优势。目前,在水稻中展开了大量的GWAS研究,对水稻的代谢性状、叶绿素含量、抽穗期、穗型、种子发芽率、芒长、耐冷、柱头外露等很多性状展开了GWAS分析,解释了各个性状的表型变异和遗传基础[10-14]。同样,水稻种子耐低氧、耐淹相关性状也运用GWAS技术进行了研究,王洋等[15]用91对SSR标记在94个太湖流域核心种质组成的自然群体中采用关联分析的方法挖掘遗传控制位点,共找到6个SSR标记与耐淹萌发关联的位点。HSU等[16]利用高密度SNP标记对153份水稻品种进行关联研究,共找到88个与低氧萌发显著关联的SNPs位点,其中部分位点与前人报道相符。Lee等[17]报道了一个控制水稻淹水萌发的基因,该基因的存在能够使水稻种子在淹水时快速生长出胚芽鞘以应对淹水胁迫,而该基因的功能缺失突变体淹水萌发时不能长出胚芽鞘。随着基因组测序技术的不断完善,利用测序获得极高密度SNP将显著提升GWAS策略在遗传位点鉴定方面的效率。【本研究切入点】耐淹成苗率是影响直播稻产量的重要性状。耐淹成苗率的高低与种子活力以及耐低氧萌发密切相关。现有关于水稻耐淹成苗的研究多是以芽鞘性状作为指标,而耐淹成苗率与种子活力的关系报道较少。此外,前人进行全基因组关联分析定位的与耐淹成苗相关的位点较少。【拟解决的关键问题】对影响耐淹成苗率的关键表型性状进行分析,挖掘相关的遗传位点和候选基因,为直播稻品种选育和耐淹成苗机理研究提供一定的理论和材料基础。
1 材料与方法
1.1 试验材料
材料包括200份种质资源,来自于中国(主要来自广东)、菲律宾国际水稻研究所(IRRI)、越南、印度、尼泊尔、韩国等国家和地区,材料具体名称见电子版附表1。
1.2 正常条件下种子萌发试验
200份种质材料,每个材料设3次生物学重复,每个生物学重复取200粒种子(经过挑选,去除空粒、瘪粒及发霉的种子),共取600粒种子,放置在30℃恒温光照培养箱中培养7 d。测量发芽率、发芽指数、活力指数。
发芽率=(发芽种子数/种子总数)×100%
发芽指数=∑(Gt/Dt),Gt指第t天新萌发的正常幼苗数,Dt指发芽的天数。
活力指数=∑(Gt/Dt)×幼苗的干重
1.3 低氧条件下种子萌发试验
200份种质材料,每个材料设3次生物学重复,每个生物学重复取20粒种子(经过挑选,去除空粒、瘪粒及发霉的种子),共取60粒种子,放置在灌满水的50 mL透明塑料管中,以此来模拟淹水的低氧环境,放置在30℃恒温光照培养箱中培养6 d。6 d后将种子扫描成图像,利用根系扫描仪(Epson LA2000)测量芽鞘长度、芽鞘直径。
1.4 耐淹成苗试验
200份种质材料,每个材料设3次生物学重复,每个生物学重复取30粒种子(经过挑选,去除空粒、瘪粒及发霉的种子),共取90粒种子。在12 cm直径塑料花盆加3 cm厚的砂土,放置种子,种子上覆0.5 cm的砂土,加水至水深10 cm,自然条件下培养20 d,20 d后测量成苗率。以种子的叶伸出水面并具备正常的根为存活个体,调查存活种子占全部种子的比例为成苗率。
成苗率=(成苗数/种子总数)×100%
1.5 简化基因组测序与全基因组关联分析
利用CTAB法进行DNA提取,检测合格后利用Illumina HiSeq测序平台,进行双末端(Paired-End)150测序。采用SAMTOOLS等软件进行群体SNP的检测。利用贝叶斯模型检测群体中的多态性位点,经过条件为dp2、Miss0.9、Maf0.01的过滤后,将得到的高质量SNP,利用ANNOVAR软件进行群体SNP注释。运用TreeBest软件计算距离矩阵,以此为基础,通过邻接法(neighbor-joining method)构建系统进化树。通过GCTA软件计算特征向量以及特征值,并利用R软件绘制PCA分布图。使用GEMMA对性状进行关联分析,通过关联的显著度(-value),筛选出候选SNPs。
1.6 转录组测序试验处理
选用籼稻品种玉针香与粳稻品种02428,分别在30℃有氧条件下培养3 d,第4天转入装满水的离心管中进行低氧处理1 d;同时在30℃的装满水的离心管中,低氧条件下培养3 d,第4天转入有氧条件培养1 d。每个处理设3个重复,每个重复每天取样1 g,锡箔纸包裹置于液氮中速冻,-80℃保存备用。对样品中的转录组RNA进行高通量测序,将得到的测序片段(reads)比对到参考序列上,计算参考序列的每个基因上reads的覆盖深度,并进行归一化,所得的结果用以表示基因的表达量。利用基于R语言的软件包edgeR,处理RNA-seq数据成对样本组间差异显著性。利用FDR与log2FC来筛选差异基因,筛选条件为FDR<0.05且|log2FC|>1。委托广州基迪奥生物科技有限公司构建RNA测序文库并完成高通量测序。
1.7 数据处理
使用Excel2010进行数据的录入和整理,使用SPSS19.1对所有表型数据进行处理分析,计算表型的平均值、标准差等,并进行方差分析、相关性分析等。
2 结果
2.1 200份材料的性状表现
200份材料在种子活力和低氧萌发性状方面具有丰富变异(表1)。其中,活力指数和耐淹成苗率的变异系数均超过了30%,而发芽率的变异系数最小,仅为5.1564%。与种子活力相关的发芽率、发芽指数、活力指数变异系数具有较大差别,发芽指数和活力指数变异系数显著高于发芽率,表明发芽指数和活力指数可以更好地体现材料间的差异。胚芽鞘长变异系数显著高于胚芽鞘直径,表明不同品种的胚芽鞘长度差别明显,而胚芽鞘直径的差别较小。耐淹成苗率在不同品种间也具有较大差异,说明研究材料在此性状上具有广泛的遗传分离。由性状频率分布图(图1)可见,上述性状均呈连续分布,表明是由微效多基因控制的数量性状。
2.2 各性状间的相关性分析
相关分析表明,耐淹成苗率与发芽率、发芽指数、活力指数、胚芽鞘长度呈极显著正相关,其中,胚芽鞘长与耐淹成苗率的相关性最高,达0.271(表2)。此外,不同性状间表现出复杂的相关性,发芽率与发芽指数呈极显著正相关,与胚芽鞘长显著正相关,与胚芽鞘直径呈极显著负相关;发芽指数与发芽率、活力指数、胚芽鞘长均呈极显著正相关,与胚芽鞘直径呈极显著负相关;活力指数与发芽指数、胚芽鞘长呈极显著正相关;胚芽鞘长与发芽指数、活力指数呈极显著正相关,与发芽率、胚芽鞘直径显著正相关;胚芽鞘直径与胚芽鞘长显著正相关,与发芽率、发芽指数、活力指数呈极显著负相关。上述结果表明,胚芽鞘长和活力指数是影响耐淹成苗率的关键性状。
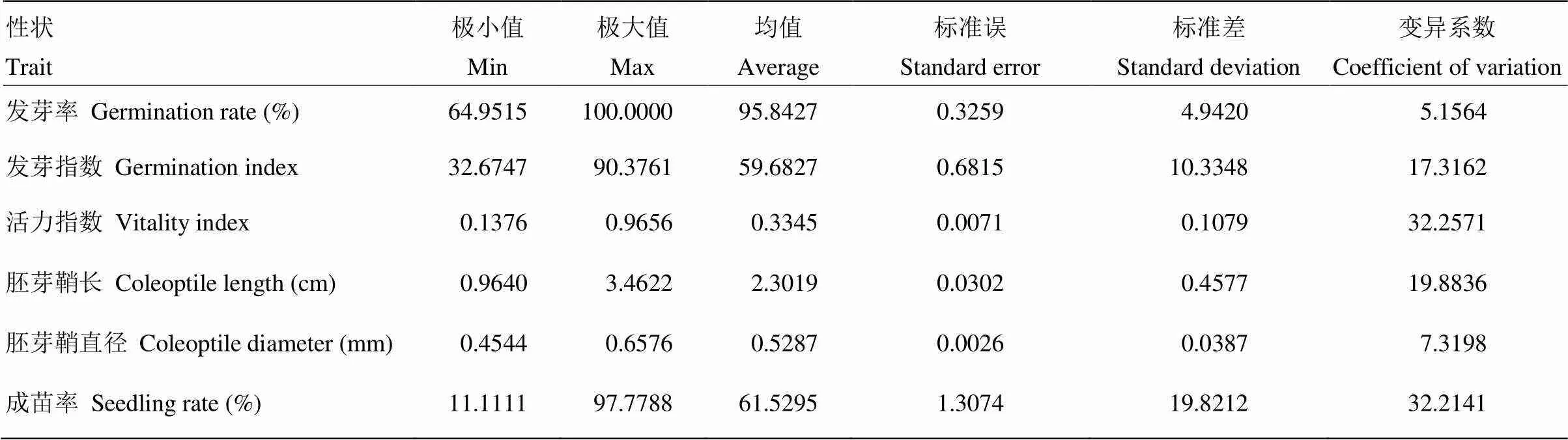
表1 表型性状

表2 表型相关性分析
*<0.05条件下差异显著;**<0.01条件下差异极显著
*represent significant variation at<0.05; **represent highly significant variation at<0.01
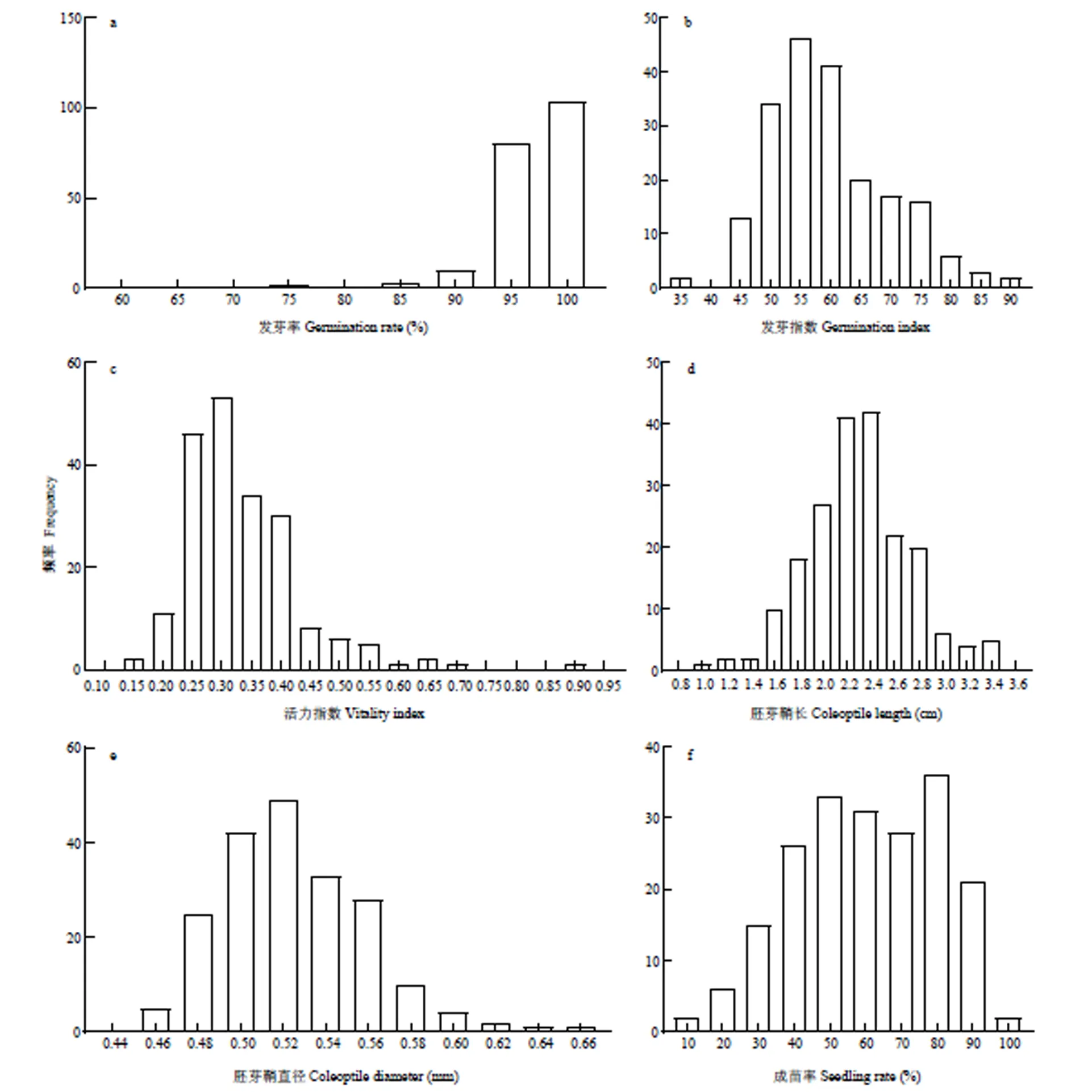
a:发芽率;b:发芽指数;c:活力指数;d:胚芽鞘长;e:胚芽鞘直径;f:耐淹成苗率
2.3 胚芽鞘长和活力指数性状的全基因组关联位点检测
2.3.1 SNP获取 利用200份种质进行简化基因组测序,基因组的平均测序深度为14.16X,平均覆盖度为11.24%。利用SAMTOOLS软件检测共获得了652 457个SNP位点,经过条件为dp2、Miss0.9、Maf0.01的过滤后,最后共获得了161 657个高质量的SNP位点用于后续分析。
2.3.2 群体结构分析 利用SNP数据对200份种质的群体进行结构分析,可见此关联群体主要可分为2个亚群,第一个亚群包括6个材料;另外一个亚群包括其他194份材料(图2-a)。通过GCTA软件计算特征向量以及特征值,并利用R软件绘制PCA分布图(图2-b),PCA分析结果与系统进化树结果一致。
2.3.3 活力指数关联位点鉴定 对200份材料的活力指数进行全基因组关联分析(图3)。左侧为曼哈顿图,图中水平的虚线表示显著性水平,当-lg10()值高于虚线时,认为该SNP与该性状显著关联;右侧为Q-Q图(Quantile-quantile plot),表示实际值和无关联零假设期望值的分布。Manhattan图与Q-Q图均表明在全基因组范围内存在与活力指数显著关联的位点。以-lg10()=6为阈值,共检测到了8个与活力指数显著关联的SNP位点(表3),分布在第1、2、5、6、10、11和12染色体上。依据LD衰减距离,对关联SNP位点上下游各100 kb区域进行了基因筛选,共筛选得到154个基因。依据基因组注释,筛选出6个与物质代谢、逆境胁迫相关的基因(表4)。


图3 活力指数曼哈顿图及Q-Q图
2.3.4 胚芽鞘长关联位点鉴定 图4为胚芽鞘长关联分析的曼哈顿图和Q-Q图(quantile-quantile plot)。以-lg10()=4为阈值,共检测到了15个与胚芽鞘长度显著关联的SNP位点,分布在第3、4、5、6、8和11染色体上(表5)。基于水稻基因组注释,依据LD衰退水平,本研究在关联 SNP位点上下游区域100 kb,共检测到296个基因。依据基因组注释,筛选出6个与种子生长发育以及逆境胁迫相关的基因(表6)。

表3 活力指数关联位点

表4 活力指数关联位点候选基因

图4 胚芽鞘长曼哈顿图和Q-Q图
2.3.5 候选基因表达模式分析 基于粳稻02428与籼稻玉针香转录组测序结果,获得上述13个基因在有氧、无氧及氧气转换下的基因表达热图(图5)。有3个基因表现出对氧气处理的敏感性,分别为、和进一步对这三个基因的表达进行分析(图6),可知与在02428和玉针香中表达模式相近,主要体现在:(1)有氧条件下的表达高于无氧;(2)从有氧转换到低氧条件,其表达显著降低;(3)从无氧转换到有氧,其表达上升。与活力指数性状关联,编码AP2结构域蛋白的基因;与胚芽鞘长性状关联,编码含BURP结构域蛋白的基因。编码叶绿素a-b结合蛋白基因,与胚芽鞘长性状关联,该基因在2个材料中的表达模式有较大不同。在02428中,该基因在低氧转换到有氧时表达量显著上调,并且在有氧转换到低氧时表达量也有上调;在玉针香中,该基因同样在低氧转换到有氧时表达量明显上调,但是在有氧转换到低氧时表达量并无明显差异。

表5 胚芽鞘长关联位点

表6 胚芽鞘长关联位点候选基因
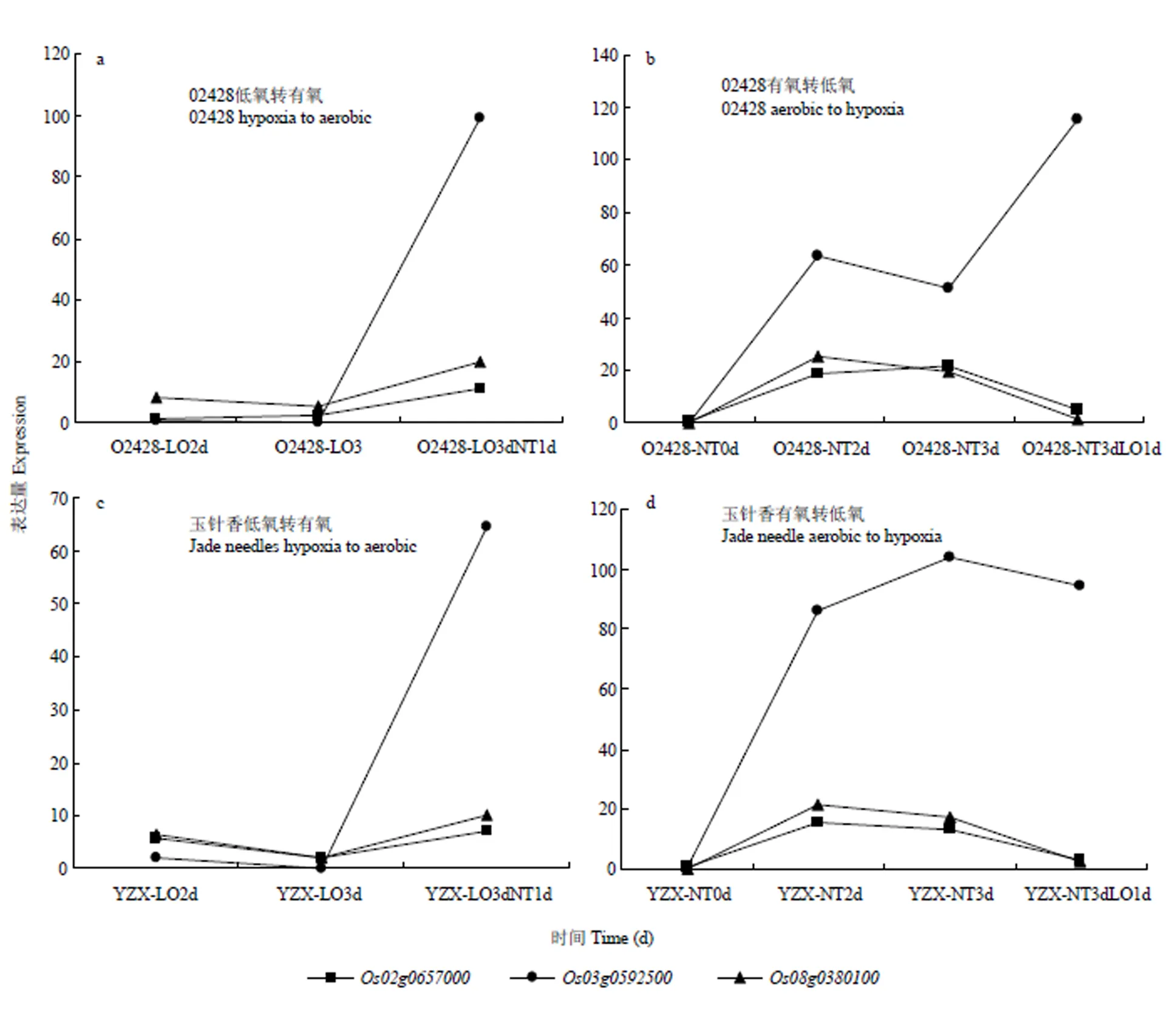
图6 候选基因表达模式
3 讨论
3.1 活力指数和胚芽鞘性状是影响直播稻全苗率的关键性状
全苗率低一直是直播稻产量不高不稳的首要问题,它直接影响群体的起点苗数,进而影响群体质量。一般认为三叶期成苗数达到落田稻粒数70%以上,基本上达到全苗目标,但生产上往往达不到50%[30]。有学者提出通过选用耐逆境发芽出苗较好的水稻品种可有效解决直播稻全苗问题。因此,种子活力和耐缺氧能力是直播稻品种选育的重要指标。
直播稻在淹水萌发期间可以分为2个时期,第一个时期为胚芽鞘的快速生长,此时通过芽鞘的快速伸长突破水面以获取氧气,根和第一片叶的生长受到抑制[31-32];第二个时期为胚芽和胚根的形态建成,此时胚芽鞘停止生长,胚芽和胚根在胚芽鞘输送氧气的支持下快速发育,完成幼苗形态构成,种子活力在此过程起重要作用[33]。本研究结果表明,活力指数与芽鞘长的变异系数远大于其他表型,因此活力指数和芽鞘长可以更好地反映遗传差异。进一步分析后发现耐淹成苗率与活力指数以及胚芽鞘长度呈极显著正相关,与前人研究相符[33-34]。因此,高活力指数与发达的胚芽鞘是选育优良直播稻品种必须考虑的重要性状,对控制这些性状的遗传位点挖掘具有重要意义。
3.2 GWAS是鉴定活力指数与芽鞘长性状遗传位点的有效方法
本研究利用基于重测序的高密度SNP标记,对活力指数和胚芽鞘性状进行全基因组关联分析。与基于RIL、BIL、DH群体的QTL定位方法比较,全基因组关联省去构建家系的时间,所能鉴定的遗传位点更加精细,更适用于耐低氧萌发复杂性状的分析。本研究共检测到8个与活力指数显著关联的SNP位点,其中有4个位点与前人报道区间位于同一位置,第2染色体26540479位置与Bi等[18]报道的基因座位相同,同时也与Sugiyama等[19]报道的基因座位相同;第6染色体5542656位置与Moeder等[20]报道的基因座位相同;第10染色体13589547位置与Mori等[21]报道的基因座位相同,同时也与Miyoshi等[22]报道的基因座位相同;第12染色体23449435位置与Miyoshi等[22]报道的基因座位相同,同时也与Zeng等[23]报道的基因座位相同,这4个位点在前人研究中分别控制了水稻细胞的增殖发育、细胞程序性死亡以及水稻花粉管发育等性状。本研究同时检测到15个与胚芽鞘长度显著关联的SNP位点,有4个位点与前人报道区间位于同一位置,第5染色体15450007位置与Liang等[24]报道的基因座位相同;第6染色体21932341位置与Singh等[25]报道的基因座位相同,同时也与Zhu等[26]报道的基因座位相同;第6染色体29822164位置与Yong等[27]报道的基因座位相同,同时也与Zhong等[28]报道的基因座位相同;第8染色体5689664位置与Cheung等[29]报道的基因座位相同,这4个位点分别控制了水稻能量运转、抗病以及干旱胁迫、高温胁迫等性状。这些共定位的位点一方面验证了本研究结果的可靠性,另一方面也提供了更多遗传信息,前人并未对共定位位点与水稻种子活力和耐淹能力的性状进行描述,暗示这些位点可能具有多效性。此外,本研究也获得19个前人未报道的控制种子活力或耐低氧萌发位点,为候选基因的筛选提供了基础。
3.3 水稻耐低氧萌发过程可能受到逆境胁迫、光合作用相关基因的调控
根据02428和YZX 2个材料的转录组测序结果,本研究对筛选出的13个关联位点区间候选基因在有氧条件、低氧条件及氧气条件转换下的表达模式进行了分析,发现有3个基因表现出对氧气处理的敏感性,分别为、和。其中,位于种子活力指数显著关联区间,该基因编码含AP2结构域蛋白的基因。在高等植物中,具有AP2结构域的转录因子在植物的开花发育以及抗逆过程中起重要作用[35],根据转录组结果可知,该基因在低氧条件下的表达量较低,而在有氧条件下表达量较高,因此,该基因的高表达对种子活力增强具有促进作用。位于胚芽鞘长显著关联区间,为编码叶绿素a-b结合蛋白基因,此类蛋白复合体能捕获光能,并迅速把光能传到PSⅠ和光系统Ⅱ的反应中心,引起光化学反应,将光能转化为化学能[36],是植物进行光合作用的重要组成部分。该基因在两个材料中的表达模式有较大不同,在02428中,该基因在低氧转换到有氧时表达量显著上调,并且在有氧转换到低氧时表达量也有上调;在玉针香中,该基因同样在低氧转换到有氧时表达量明显上调,但是在有氧转换到低氧时表达量并无明显差异。在低氧转换到有氧的处理中,因种子在淹水萌发前期只进行芽鞘的快速生长,芽鞘并不含叶绿素,几乎不进行光合作用,转换到有氧条件后,茎秆以及叶片开始生长,叶绿素大量合成,光合作用增强,因此表达量大幅上升;而在有氧转换到低氧的处理时,种子在有氧条件下正常萌发生长,光合作用随着种子萌发即开始进行,转换到低氧后由于叶片已经长出,光合作用仍在进行,因此,表达量变化并不明显,推测该基因对水稻种子耐淹生长具有正调控作用。此外,由于低氧条件下02428的胚芽鞘生长快于玉针香,因此,该基因还可能是低氧条件下02428和玉针香胚芽鞘生长差异的影响因子。位于胚芽鞘长显著关联区间,该基因编码含BURP结构域蛋白的基因,此类蛋白不仅与植物的生殖发育相关,而且在植物耐受非生物胁迫的过程中具有重要的作用[37]。根据转录组结果可知,该基因的表达模式与相近,在低氧条件下的表达量较低,而在有氧条件下表达量较高,推测该基因的高表达抑制了胚芽鞘的生长,对种子耐低氧胁迫起负调控作用。
4 结论
水稻耐淹成苗率与芽鞘长、活力指数呈极显著正相关,以种子活力、芽鞘长度为指示性状可以筛选出耐淹成苗能力较强的材料。全基因组关联分析、转录组分析与基因表达模式比较的联合应用可提高相关性状候选基因的筛选效率。水稻耐淹萌发过程可能受到与逆境胁迫、光合作用相关基因的调控。
[1] 刘贵富, 陈明江, 李明, 吕慧颖, 葛毅强, 魏珣, 杨维才. 水稻育种行业创新进展. 植物遗传资源学报, 2018, 19(3): 416-429.
Liu G f, Chen M j, Li M, Lu H y, Ge Y q, Wei X, Yang W c. Innovation progress in rice breeding industry., 2018, 19(3): 416-429. (in Chinese)
[2] 章孟臣. 水稻耐淹发芽相关性状的全基因组关联分析[D]. 北京: 中国农业科学院, 2016.
Zhang M C. Genome-wide association analysis of traits related to flooding and germination in rice [D]. Beijing: Chinese Academy of Agricultural Sciences, 2016. (in Chinese)
[3] Yamauchi M, Aguilar A M, Vaughan D A, Seshu D V. Rice (L.) germplasm suitable for direct sowing under flooded soil surface., 1993, 67(3): 177-184.
[4] Yamauchi M, Biswas J K. Rice cultivar difference in seedling establishment in flooded soil., 1997, 189(1): 145-153.
[5] 陈孙禄, 王俊敏, 潘佑找, 马健阳, 张建辉, 张红生, 滕胜. 水稻萌发耐淹性的遗传分析. 植物学报, 2012, 47(1): 28-35.
Chen S l, Wang J m, Pan Y Z, MA J Y, ZHANG J H, ZHANG H S, TENG S. Genetic analysis of rice germination tolerance to flooding., 2012, 47(1): 28-35. (in Chinese)
[6] Septiningsih E M, Sendon P M D, Sanchez D L, Ismail A M, Mackill D J. QTL mapping and confirmation for tolerance of anaerobic conditions during germination derived from the rice landrace Ma-Zhan Red., 2013, 126(5): 1357-1366.
[7] Lee H S, Sasaki K, Kang J W, Sato T, Song W Y, AHN S N. Mesocotyl elongation is essential for seedling emergence under deep-seeding condition in rice., 2017, 10(1): 32.
[8] Mackay I, Powell W. Methods for linkage disequilibrium mapping in crops., 2007, 12(2): 57-63.
[9] Mackay T F C, Stone E A, Ayroles J F. The genetics of quantitative traits: challenges and prospects., 2009, 10(8): 565-577.
[10] Bai X, Zhao H, Huang Y, Xie W B, Han Z M, Zhang B, Guo Z L, Yang L, Dong H J, Xue W Y, Li G W, Hu G, Hu Y, Xing Y Z. Genome-wide association analysis reveals different genetic control in panicle architecture between and rice., 2016, 9(2).
[11] Han Z, Zhang B, Zhao H, AYAAD M, XING Y Z. Genome-wide association studies reveal that diverse heading date genes respond to short and long day lengths betweenandrice., 2016, 7: 1270.
[12] Magwa R A, Zhao H, Xing Y. Genome-wide association mapping revealed a diverse genetic basis of seed dormancy across subpopulations in rice (L.)., 2016, 17(1): 28.
[13] Magwa R A, Zhao H, Yao W, XIE W, YANG L, BAI X. Genomewide association analysis for awn length linked to the seed shattering gene qSH1 in rice., 2016, 95(3): 1-8.
[14] Zhou P b, Xie W b, HUSSIAN S, LI Y, XIA D, ZHAO H, SUN S, CHEN J, YE H, HOU J, ZHAO D, GAO G, ZHANG Q, WANG G, LIAN X, XIAO J, YU S, LI X, HE Y. Genome-wide association analyses reveal the genetic basis of stigma exsertion in rice., 2017, 10(4): 634-644.
[15] 王洋. 适于直播的水稻种质资源筛选及种子活力和幼苗耐缺氧能力优异等位变异的发掘[D]. 南京: 南京农业大学, 2009.
Wang Y. Screening of rice germplasm resources suitable for direct seeding and excavation of seed vigor and excellent allergic variation of seedling tolerance to hypoxia [D]. Nanjing: Nanjing Agricultural University, 2009. (in Chinese)
[16] Hsu S K, Tung C W. Genetic mapping of anaerobic germination- associated QTLs controlling coleoptile elongation in rice., 2015, 8(1): 1-12.
[17] Lee K W, Chen P W, Lu C A, CHEN S, HO Y H, YU S M. Coordinated responses to oxygen and sugar deficiency allow rice seedlings to tolerate flooding., 2009, 2(91): ra61.
[18] Bi F C, Zhang Q F, Liu Z, Fang C, Li J, Su J B, Greenberg J T, Wang H B, Yao N. A conserved cysteine motif is critical for rice ceramide kinase activity and function., 2011, 6(3): e18079.
[19] Sugiyama K, Hayakawa T, Kudo T, Ito T, Yamaya T. Interaction of N-acetylglutamate kinase with a PII-like protein in rice., 2004, 45(12): 1768-1778.
[20] Moeder W, Yoshioka K. CNGCs break through-A rice cyclic nucleotide-gated channel paves the way for pollen tube growth., 2017, 13(11): e1007066.
[21] Mori Y, Yamamoto T, Sakaguchi N, Ishibashi T, Furukawa T, Kadota Y, Kuchitsu K, Hashimoto J, Kimura S, Sakaguchi K. Characterization of the origin recognition complex (ORC) from a higher plant, rice (L.)., 2005, 353(1): 23-30.
[22] Miyoshi K, Ahn B O, Kawakatsu T, Ito Y, Itoh J, Nagato Y, Kurata N. PLASTOCHRON1, a timekeeper of leaf initiation in rice, encodes cytochrome P450., 2004, 101(3): 875-880.
[23] Zeng L R, Qu S, Bordeos A, Yang C, Baraoidan M, Yan H, Xie Q, Nahm B H, Leung H, Wang G L. Spotted leaf11, a negative regulator of plant cell death and defense, encodes a U-box/ armadillo repeat protein endowed with E3 ubiquitin ligase activity., 2004, 16(10): 2795-2808.
[24] Liang Y, Zhao X, Jones A M, GAO Y. G proteins sculp root architecture in response to nitrogen in rice and., 2018, 274: 129-136.
[25] Singh G, Sarkar N K, Grover A. Mapping of domains of heat stress transcription factor OsHsfA6a responsible for its transactivation activity., 2018, 274: 80-90.
[26] Zhu S, Gao F, Cao X, Chen M, Ye G, Wei C, Li Y. The rice dwarf virus P2 protein interacts with ent-kaurene oxidases, leading to reduced biosynthesis of gibberellins and rice dwarf symptoms., 2005, 139(4): 1935-1945.
[27] Yong H C, Kim C Y, Min C K, Kim C Y, Kim M C, Kim I H, Park C Y, Kim J C, Park B O, Koo S C, Yoon H W, Chung W S, Lim C O, Lee S Y, Cho M J. BWMK1, a rice mitogen-activated protein kinase, locates in the nucleus and mediates pathogenesis- related gene expression by activation of a transcription factor., 2003, 132(4): 1961-1972.
[28] Zhong R, Cui D, Phillips D R, YE Z H. A novel rice xylosyltransferase catalyzes the addition of 2-o-xylosyl side chains onto the xylan backbone., 2018, 59(3): 554.
[29] Cheung M Y, Xue Y, Zhou L, Li M W, Sun S S, Lam H M. An ancient P-loop GTPase in rice is regulated by a higher plant-specific regulatory protein., 2010, 285(48): 37359-37369.
[30] 蒋彭炎. 直播稻的生育特点和增产对策. 中国稻米, 1996(4): 30-33.
Jiang P y. Fertility characteristics and yield increase strategies of direct seeding rice., 1996(4): 30-33. (in Chinese)
[31] 侯名语, 江玲, 王春明, 万建民. 水稻种子低氧发芽力的QTL定位和上位性分析. 中国水稻科学, 2004, 18(6): 483-488.
Hou M y, Jiang L, Wang C m, WAN J M. QTL mapping and epistasis analysis of hypoxia germination of rice seeds., 2004, 18(6): 483-488. (in Chinese)
[32] Angaji S A, Septiningsih E M, Mackill D J, Ismail A M. QTLs associated with tolerance of flooding during germination in rice (L.)., 2010, 172(2): 159-168.
[33] 何龙生. 水稻种子活力测定方法的初步研究[D]. 杭州: 浙江农林大学, 2018.
He L s. Preliminary study on the determination method of rice seed vigor [D]. Hangzhou: Zhejiang Agriculture and Forestry University, 2018. (in Chinese)
[34] 曹栋栋, 阮晓丽, 詹艳, 石瑛琪. 杂交水稻种子不同活力测定方法与其田间成苗率的相关性. 浙江农业学报, 2014, 26(5): 1145-1150.
CAO D d, YAN X l, ZHAN Y, SHI Y q. Correlation between different vigor determination methods of hybrid rice seeds and their seedling rate in field., 2014, 26(5): 1145-1150. (in Chinese)
[35] 张淼, 赵畅, 李法莲, 邵群. 具有AP2结构域的转录因子家族的研究进展. 科技信息, 2007(21): 342-345.
ZHANG M, ZHAO C, LI F l, SHAO Q. Research progress of transcription factor family with AP2 domain., 2007(21): 342-345. (in Chinese)
[36] 朱彬彬, 崔百明, 向本春. 番茄叶绿素a/b结合蛋白基因cab-1a的克隆与定位. 江苏农业科学, 2017, 45(14): 20-23.
ZHU B b, CUI B m, XIANG B c.Cloning and localization of tomato chlorophyll a/b binding protein gene cab-1a., 2017, 45(14): 20-23. (in Chinese)
[37] 米子岚, 钟活权, 江年琼, 唐玉林. BURP蛋白家族与植物对非生物胁迫的响应. 中国细胞生物学学报, 2015(9): 1302-1308.
MI Z L, ZHONG H Q, JIANG N Q, TANG Y L. Response of BURP protein family and plants to abiotic stress., 2015(9): 1302-1308. (in Chinese)
Genome-wide association analysis for rice submergence seedling rate
Sun Kai, Li Dongxiu, Yang Jing, Dong Jichi, Yan Xiancheng, Luo Lixin, Liu Yongzhu, XIAO WuMing, Wang Hui, Chen Zhiqiang, Guo Tao
(National Plant Space Breeding Engineering Technology Research Center, South China Agricultural University, Guangzhou 510642)
【Background】The low seedling rate of direct seeding rice is an important factor limiting its yield. Mining rice materials with high seed viability and low oxygen germination ability is the key to improving the seedling rate and solving the problem of seedlings in direct seeding rice. 【Objective】The key phenotypic traits affecting the rate of emergence and tolerance of seedlings were analyzed, and the relevant genetic loci and candidate genes were mined to provide a theoretical and material basis for the study of direct seeding rice cultivars and the mechanism of resistance to flooding. 【Method】Using 200 rice germplasms from a wide range of sources. The germination test was carried out in an aerobic environment, and the seed viability phenotype was measured including germination rate, germination index and viability index; The coleoptile length and coleoptile diameter were measured under hypoxic conditions. The flood-tolerant seedling experiment was carried out, the water depth was 10cm, and the flood-tolerant seedling rate was measured after 20d. The correlation between various traits was analyzed, and the key traits affecting the rate of tolerance to flooding were explored. Genome-wide association analysis was performed on the above six phenotypes by simplified genome sequencing, and SNP sites significantly associated with traits were identified and within the correlation interval. Screening candidate genes related to the research purpose; transcriptome detection under the conditions of aerobic, anaerobic and oxygen content conversion of 02428 and YZX two materials, combined with genome-wide association analysis results, analysis of differences in expression patterns of candidate genes. 【Result】Seed viability, coleoptile phenotype and seedling rate showed extensive genetic variation among 200 materials. Among them, the variation of coleoptile length and viability index was the most abundant, and the coefficient of variation was the largest. At the same time, the results of correlation analysis showed that there was a significant positive correlation between the coleoptile length, viability index and the seedling rate. Through genome-wide association analysis, 8 sites significantly associated with the viability index and 15 sites significantly associated with the coleoptile length were identified. Based on the correlation of seed growth and development and stress resistance, six candidate genes related to viability index and seven candidate genes related to the coleoptile length were screened in the relevant interval. Further comparison of 13 genes in aerobic and anaerobic the expression patterns and expression changes under oxygen conversion conditions showed that the three genes,andshowed different expression patterns when the oxygen content changed, and the expression amount changed significantly, showing sensitivity to oxygen treatment. 【Conclusion】Seed viability and coleoptile length were closely related to the rate of flooding and seedling emergence, which could be used as an important trait for screening flood-tolerant rice materials. Combining genome-wide association analysis, transcriptome analysis and gene expression pattern can improve the screening efficiency of candidate genes for hypoxia tolerance germination of rice seeds; flooding tolerance of rice seedlings may be regulated by genes related to stress and photosynthesis..
rice; seed vitality; hypoxia-resistant germination; genome-wide association analysis; transcriptome analysis
10.3864/j.issn.0578-1752.2019.03.001
2018-09-18;
2018-11-12
国家重点研发计划(2017YFD0100104,2016YFD0102102)、广东省科技计划(2015B02031011)、国家现代农业产业技术体系建设专项(CARS-01-12)
孙凯,E-mail:m15018435726_2@163.com。通信作者郭涛,E-mail:guo.tao@vip.163.com。信作者陈志强,E-mail:zqchen@scau.edu.cn
(责任编辑 李莉)
