急性感染性腹泻沙门菌分离株的耐药谱及其相关基因分析
2019-01-10张景皓杨丽华刘文健张艳梅
张景皓,杨 峰,方 毅,杨丽华,刘文健,赵 虎,张艳梅
(1.复旦大学附属华东医院检验科,上海 200040;2.上海市松江区疾病预防控制中心,上海 201620)
急性感染性腹泻是临床常见的肠道感染性疾病之一,在我国具有较高的发病率[1]。导致急性感染性腹泻的病原菌种类较多,包括各种肠道病原菌、肠道病毒和部分真菌及原虫等。沙门菌是导致人类食源性急性腹泻最常见的病原菌[2-3]。由于急性感染性腹泻发病快,多数患者病程较短,而细菌培养、鉴定和药物敏感性试验时间较长,所以临床上多以经验用药为主,导致耐药菌株增加、疗效差和肠道微生态失衡,进而引发多种并发症[1,4]。有研究发现,沙门菌携带的耐药基因中,β-内酰胺酶类耐药以blaTEM-1为主,喹诺酮耐药主要由gyrA和parC点突变引起[5-6],磺胺类耐药基因是sulⅠ[7],而aph(3)-Ⅱa是氨基糖苷类耐药基因的代表之一[8]。本研究分析了分离自400例急性感染性腹泻患者粪便样本的22株沙门菌,检测其药物敏感性及相关耐药基因的携带情况,以了解地区差异,为临床的精准诊疗提供参考。
1 材料和方法
1.1 样本来源
收集2016年5月—2017年4月华东医院急性感染性腹泻患者粪便样本52例、上海市松江区疾病预防控制中心急性感染性腹泻患者粪便样本348例,共400例。
1.2 仪器与试剂
Vitek MS 仪、Vitek 2 Compact全自动微生物分析系统(法国生物梅里埃公司)及配套药敏板条,环丙沙星、左氧氟沙星、头孢呋辛、头孢西丁和头孢噻肟E-test试剂条购自英国Oxoid公司。血琼脂平板、SS平板和M-H平板购自上海科玛嘉微生物技术有限公司。沙门菌血清学分型采用丹麦SSI公司生产的鉴定血清。校准菌株大肠埃希菌(ATCC 8739)由法国生物梅里埃公司提供,质控菌株大肠埃希菌(ATCC 25922、ATCC3 5218)、铜绿假单胞菌(ATCC 27853)由上海市临床检验中心提供。TIANamp Bacteria DNA Ki购自天根生化科技(北京)有限公司,Veriti 96 PCR仪购自美国ABI公司,引物购自生工生物工程(上海)股份有限公司。
1.3 方法
1.3.1 细菌分离培养 挑取粪便样本,接种于SS平板,37 ℃培养18~24 h,直至形成中等大小、透明或半透明可疑菌落。
1.3.2 菌种鉴定 采用Vitek MS 仪进行菌种检测。挑取单个菌落均匀涂布于靶板上,加入1 uL基质(α氰基-4-羟基肉桂酸),室温干燥后上机检测。严格按照仪器说明书进行操作。鉴定结果可信度均为99.9%。
1.3.3 血清学分型 将分离培养阳性的沙门菌转种于血平板进行分纯和传代培养。血清凝集实验的操作按照如下步骤进行:悬空滴加1滴抗血清于载玻片上,用冷却后的灭菌接种环挑取适量生长状态良好的待检细菌,均匀涂布在抗血清中,小幅度晃动载玻片,1 min内观察是否有凝集出现。血清凝集顺序为从多价到单价,从常见血清型到少见血清型。
1.3.4 药物敏感性试验 采用Vitek 2 Compact药敏板条,并补充5种临床常用抗菌药物(环丙沙星、左氧氟沙星、头孢呋辛、头孢西丁和头孢噻肟)E-test试剂条进行药物敏感性试验。参照美国临床实验室标准化协会2016 年版判读标准判定结果[9]。
1.3.5 相关耐药基因检测 采用检测β-内酰胺类耐药基因blaTEM-1、blaCMY-2、blaPSE-1,磺胺类耐药基因sulⅠ、sulⅡ、sulⅢ,氨基糖苷类耐药基因aph(3')-Ⅱ a、aadA1、aadB、strB,喹诺酮类耐药基因qyrA、qnrB、parC。目的基因引物序列和目的片段长度见表1。聚合酶链反应(polymerase chain reaction,PCR)反应体系为:反应总体积25 μL,2xTaq PCR Master Mix 12.5 μL,正、反向引物各1 μL,模板2 μL,最后用ddH2O补至总体积25 μL[8]。

表1 耐药基因引物序列和目的片段长度
2 结果
2.1 沙门菌分离率和血清分型
剔除了分离自同一患者的重复菌株,从400例粪便样本中分离到22株沙门菌,阳性分离率为5.5%。对22株沙门菌进行血清学分型检测,检出率最高的血清型依次为鼠伤寒沙门菌(36.4%,8/22)、肠炎沙门菌(27.3%,6/22)和伦敦沙门菌(18.2%,4/22),德比沙门菌、汤普森沙门菌、纽波特沙门菌和吉伟沙门菌各检出1株(4.5%,1/22)。
2.2 体外药物敏感性试验
沙门菌整体耐药率较高,其中氨苄西林-舒巴坦最高(95.5%),以下依次为氨苄西林、左氧氟沙星、复方磺胺甲噁唑、环丙沙星-头孢噻肟-庆大霉素、头孢唑林-头孢呋辛-头孢他啶、阿米卡星。虽然环丙沙星较左氧氟沙星耐药率低,但其中介率可达63.6%。哌拉西林-他唑巴坦、亚胺培南敏感率100%。见表2。
对不同血清型沙门菌耐药情况进行分析,结果显示,不同血清型沙门菌的耐药情况分布较均一,两者间并未出现明显的相关性。
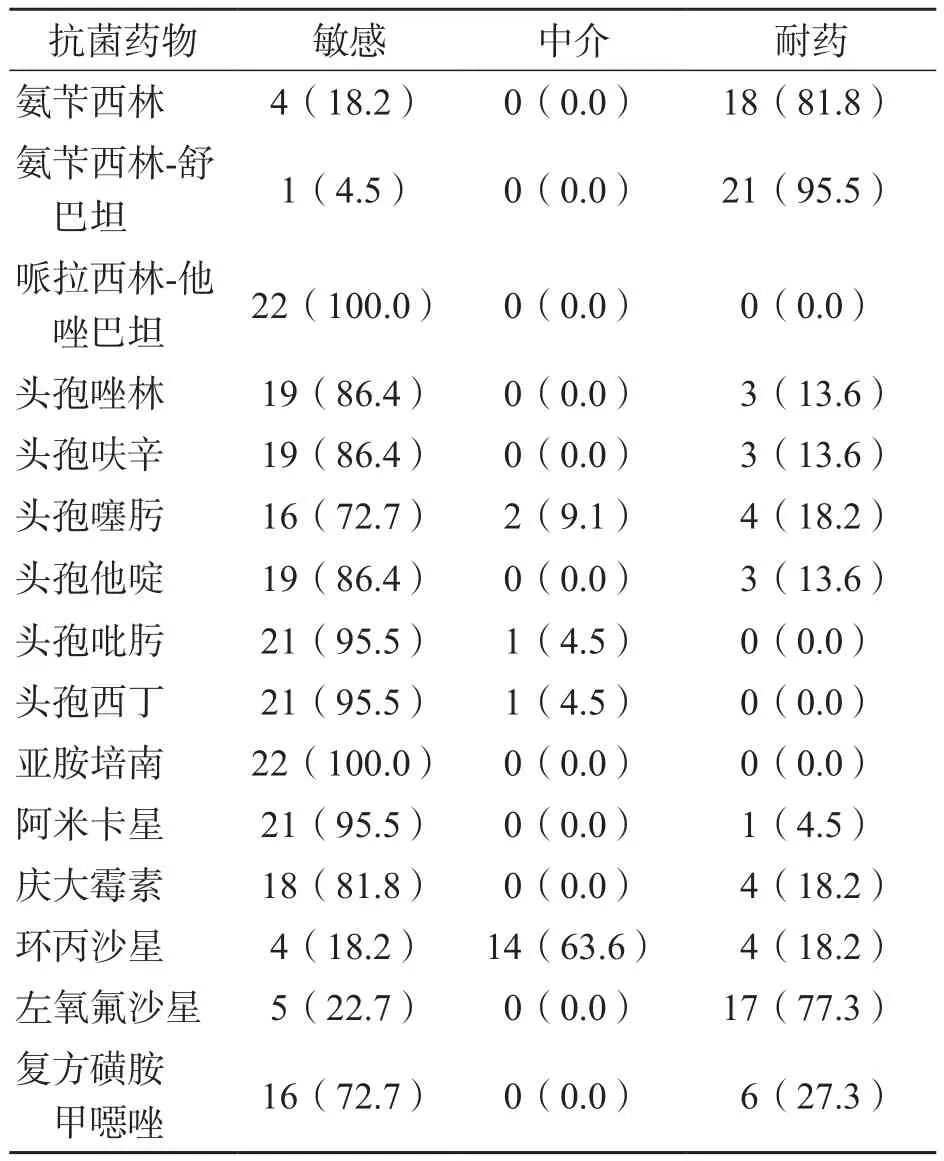
表2 沙门菌药物敏感性试验结果 [例(%)]
2.3 相关耐药基因的检测
β-内酰胺类耐药基因blaTEM-1和DHA仅在耐药菌株中被检测到,检出率均为23.8%(5/21),blaCTX-M1、blaCTX-M2、 blaCTX-M8、blaCTX-M9、blaSHV、blaCMY-2和blaPSE-1基因在所有菌株中均未被检测到。磺胺类耐药基因sulⅠ和sulⅢ仅在耐药株中被检测到,检出率分别为66.7%(4/6)和16.7%(1/6),而sul Ⅱ基因仅在敏感株中被检测到,检出率为6.25%(1/16)。氨基糖苷类耐药基因aadA1在耐药株中的检出率为25%(1/4),aph(3')-Ⅱa、aadA1、aadB和strB基因未在耐药菌株中被检测到,但在敏感株中检出率分别为5.6%(1/18)、22.2%(4/18)、5.6%(1/18)和50.0%(9/18)。喹诺酮类耐药基因qyrA、qnrB、parC和qnrS均只在耐药株中被检测到,检出率分别为47.1%、11.7%、58.8%和17.6%;qnrA在耐药菌株中的检出率为5.9%,其在敏感菌株中也被检出1例;qnrC未被检出。见表3。
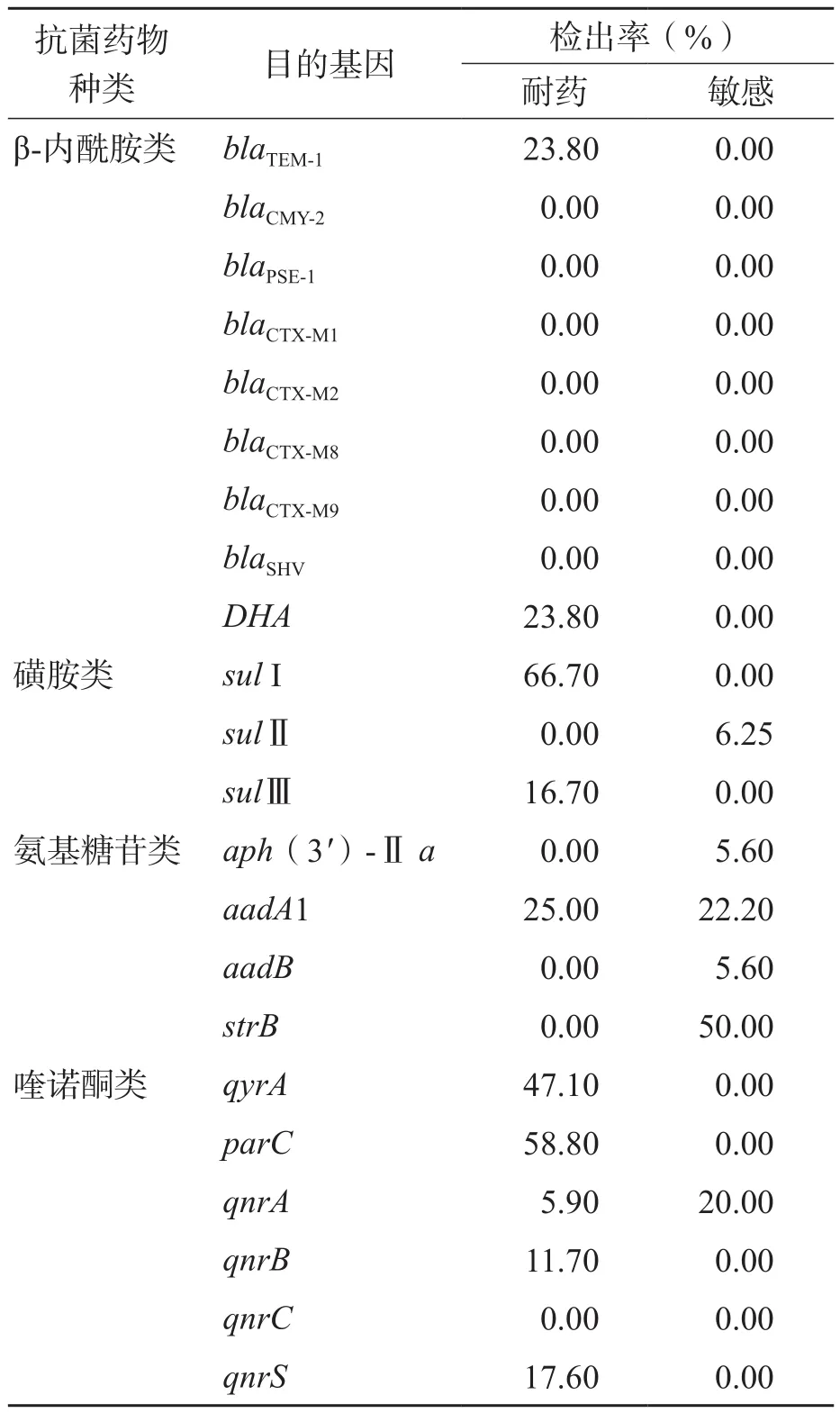
表3 沙门菌耐药基因测序结果及检出率
3 讨论
本研究从收集到的400例临床粪便样本中分离出22株沙门菌,分离率为5.5%,同其他地区报道的沙门菌分离率基本一致[11]。目前临床多将喹诺酮类药物作为对细菌感染性腹泻经验用药的首选抗菌药物,将磺胺类药物作为次选药物[1,12]。然而本研究通过对22株分离自急性感染性腹泻患者粪便样本中的沙门菌进行15种临床常用抗菌药物的药物敏感性试验后发现,沙门菌对氨苄西林-舒巴坦、氨苄西林和左氧氟沙星的耐药率分别高达95.5%、81.8%和77.3%,对复方磺胺甲噁唑及庆大霉素的耐药率也高达27.3%和18.2%,与国内其他报导有较大的差异[5,7,13]。提示在临床用药时应注意本地区沙门菌的耐药谱,并进行精准检测和分析,同时建议各地建立自己的耐药监测网,以指导临床用药。
β-内酰胺类抗菌药物的主要作用机制为结合细菌细胞膜内膜上的靶位蛋白,抑制细菌细胞壁的合成[14]。当细菌产生 β-内酰胺酶后,可使 β-内酰胺类抗菌药物水解失活。β-内酰胺酶分为 4 种类型,其中Ⅱ型酶是由质粒介导的,包括TEM、OXA、CMY 等[7]。本研究β-内酰胺类的耐药基因中只测出blaTEM-1和DHA基因,阳性率均为23.8%(5/21),而blaCTX-M1、blaCTX-M2、blaCTX-M8、 blaCTX-M9、blaSHV、blaCMY-2和blaPSE-1基因在所有菌株中均未被检测到,与国内其他报道在耐β-内酰胺类抗菌药物沙门菌株TEM携带率较高一致[5,13]。
氨基糖苷类抗菌药物的耐药机制主要是氨基糖苷类钝化酶的产生使氨基糖苷类抗菌药物对菌株无法发挥作用,同一种抗菌药物因药物的分子结构中可能存在多个结合位点又可被多种钝化酶所钝化,有报道指出伤寒沙门菌以AAC(3)Ⅱ为主[15]。本研究中,氨基糖苷类抗菌药物耐药株携带钝化酶aadA1基因检出率为25%(1/4),而且在敏感菌株中也有相当高的携带率,提示还有其他耐药机制影响其耐药表型,如抗菌药物的作用靶位改变、细胞壁通透性改变或细胞内转运异常等[15-16]。
喹诺酮类抗菌药物耐药机制主要是其作用于DNA解旋酶和拓扑异构酶,其亚单位分别由gyrA、gyrB和parC、parE基因编码[15]。沙门菌中gyrA基因是氟喹诺酮类药物的最初靶位,位于gyrA蛋白第67和106位氨基酸残基之间,喹诺酮耐药决定区中经常突变而引起对氟喹诺酮类药物的抗性;parC基因是拓扑异构酶Ⅳ编码基因,也可能是喹诺酮类药物的作用靶位,parC中第57位、第80位突变均可引起抗性产生[17]。 本研究结果表明沙门菌对喹诺酮类抗菌药物的耐药率较高,其基因携带率qyrA4为47.1%(8/17)、parC为58.8%(10/17)、qnrC为17.6%(3/17),qnrB为11.7%(2/17),而敏感菌株均未被测出,提示qyrA4、parC、qnrC和qnrB基因仍是喹诺酮类抗菌药物的主要耐药机制。
二氢蝶呤氨苯甲酸合成酶是细菌细胞生长所必需的叶酸生物合成途径中的一部分,磺胺可与二氢蝶呤氨苯甲酸合成酶相互作用,从而阻止细菌叶酸的合成,抑制细菌的生长繁殖。细菌编码二氢蝶酸合成酶的耐药基因有sul Ⅰ、sulⅡ、sul Ⅲ3种[15],本研究结果显示,磺胺类耐药菌株主要携带sul Ⅰ基因[66.7%(4/6)],其次为sul Ⅲ基因[16.7%,(1/6)]。
本研究还发现,在18株对氨基糖苷类敏感的菌株中,有15株分别携带有aph(3')-Ⅱ a基因[5.6%,(1/18)]、aadA1基因[22.2%,(4/18)]、aadB基因[5.6%,(1/18)]和strB基因[50.0%,(9/18)]。同时,在16株对磺胺类敏感的菌株中,有1株携带sul Ⅱ基因[6.25%,(1/16)]。以上结果提示这些耐药相关基因可能不是独立发挥耐药作用。
综上所述,本研究结果提示上海地区沙门菌的主要血清型为肠炎沙门菌和鼠伤寒沙门菌,而且其耐药状况严重,耐药机制复杂,很多机制还有待于深入研究。同时,不同地区耐药情况也有所差异,提示临床应该根据药物敏感性试验结果进行精准的诊断和治疗。
