动植物蛋白比恒定的饲料中发酵豆粕对刺参生产性能、消化酶和非特异性免疫的影响
2019-01-02包鹏云丛玉婷李晓宇徐永平
■ 包鹏云 丛玉婷 李晓宇 徐永平
(1.大连理工大学生命科学与技术学院,辽宁大连116024;2.大连海洋大学水产与生命学院,辽宁大连116023;3.动物性食品安全保障技术教育部工程研究中心,辽宁大连116620)
刺参属于“海产八珍”之一,含有丰富的多糖、皂苷及胶原蛋白等多种生物活性物质和矿物质元素,具有药食同补功能,对提高人体免疫力,抵抗各种疾病和应激具有非常显著的功效,颇受广大消费者的认可,成为最受欢迎的海产品之一。通过最近15年的发展,刺参养殖业已处于平稳发展阶段,养殖技术日趋成熟,市场供需处于较为平衡的状态。2015年,刺参全国总产量达到20.58万吨[1],实现产值约300多亿元,刺参已经成为我国海水养殖业中单一品种,产值和经济效益最高的品种。刺参产业已经成为北方沿海地区第一大渔业产业,对于实现渔业增效、渔民增收,促进北方沿海地区新农村建设和渔业产业的发展意义重大。
刺参属于低等的棘皮动物,是底栖的,腐屑食性,杂食性偏植物性的海洋经济类养殖动物。其索饵空间狭小,活动距离短,不能像鱼、虾等游泳动物一样自由活动和选择最适的生存环境和优质饵料,优质天然饵料的获取是刺参生长非常重要的限制性因素之一。随着刺参大规模池塘养殖和海上浮筏网笼养殖的开展,天然饵料已不能满足刺参规模化养殖的实际需要,人工配合饲料的开发和应用成为刺参营养学和饲料学研究的热点。
决定刺参健康、生长和产品品质的主要营养成分是饲料中的蛋白质,饲料中所需的动物蛋白源的价格远高于植物蛋白源的价格,又因刺参的食性决定了其所摄食的配合饲料蛋白质营养浓度较低,动物蛋白源又必不可少,为了控制刺参饲料成本,实现刺参饲料的营养平衡、环保与高效利用,本试验在前期研究确定饲料中动植物蛋白比为1∶3时,刺参有最佳生长效果的基础上,进一步研究饲料中发酵豆粕添加量对刺参生产性能和消化酶、免疫酶活性的影响,以期为动植物蛋白源鱼粉、发酵豆粕、豆粕等在刺参配合饲料中的合理应用提供科学依据。
1 材料与方法
1.1 试验设计与饲料配制
刺参配合饲料按等氮等脂原则设计和配制,生产鱼粉含量为6.5%,动植物蛋白比为1∶3的6种配合饲料(D0为对照组,D4%、D8%、D12%、D16%、D20%五个试验组饲料是用发酵豆粕按4%、8%、12%、16%和20%的比例取代日粮中豆粕)。其中以鱼粉、大豆粕、发酵豆粕、玉米蛋白粉为动植物蛋白源,以马尾藻粉和α-淀粉为碳水化合物,以鱼油为脂肪源。所有饲料原料用超微粉碎机进行粉碎后95%以上过200目筛网,而后试验用制粒机制成直径5 mm的硬颗粒饲料,45℃风干后装于4℃冰箱保存备用。日粮组成及营养成分见表1。
1.2 动物饲养与管理
试验刺参,暂养阶段的刺参放于室内可控温的水槽中投喂商品饲料,暂养15 d,温度控制在(12±1)℃,使之逐渐适应试验水环境。暂养结束后,试验刺参饥饿24 h,称重,挑选个体大小均匀的健康刺参随机分配到18个水槽(60 cm×40 cm×40 cm)中进行饲养试验。选取初始体重为(44.47±1.45)g刺参,随机分为6组,每组3个平行,每个平行组15头刺参。正式试验持续40 d。每天投喂饲料为刺参体重的3%,以后根据每天各水槽中刺参的摄食情况进行适当调整投喂量,以1~2 h摄食完为准,每天早晚各投喂1次,吸除残饵和粪便,每天换新鲜海水三分之一。饲养期间连续充气,水温控制在12~15 ℃,盐度28~32,pH值7.8~8.5,溶解氧高于5 mg/l,氨氮不高于 0.2 mg/l,亚硝氮不高于0.1 mg/l。
1.3 称重与酶活性的测定
试验结束后,将刺参饥饿24 h以排空其肠道内的粪便,用干燥的白毛巾吸干体表水分,称刺参的终末体重;从每个水槽取5头刺参,用医用注射器抽取体腔液4~5 ml后,将刺参于冰盘上解剖,用生理盐水冲洗消化道内容物,然后将生理盐水与消化道内含物按10∶1混合,并于冰浴下进行匀浆,4℃下10 000 r/min离心15 min,取上清液进行消化酶活性的测定。体腔液也进一步在冰浴中用匀浆器匀浆,以便将各种细胞中的酶释放出来,在4℃时,12 000 r/min离心20 min,取上清液分装于离心管中,在-80℃冰箱保存,待测定各项酶指标。超氧化物歧化酶、过氧化氢酶、酸性磷酸酶、碱性磷酸酶活性和总抗氧化能力的测定均采用试剂盒法(南京建成生物工程研究所)测定,操作步骤按说明书进行。

表1 日粮组成及营养水平
1.4 饲料营养成分的测定
试验饲料和刺参体壁营养成分含量采用概率养分分析方法测定:干物质含量采用烘干法测定,105℃烘干至恒重;粗蛋白质含量采用凯氏定氮法测定;粗脂肪含量采用索氏抽提法测定;粗灰分含量采用高温灼烧法测定,在550℃灼烧至恒重[2]。
特定生长率(%/d)=100×(lnW2-lnW1)/t增重率(%)=100×(W2-W1)/W1体壁系数(%)=100×FBW/W2
式中:W1——刺参的初始体重(g);
W2——刺参的末体重(g);
FBW——刺参的终末体壁重(g);
t——试验时间(d)。
1.5 数据处理
采用SPSS17.0软件进行单因素方差分析(One Way ANOVA),采用Duncan′s法多重比较进行差异显著性检验,P<0.05为显著性差异,用不同字母进行标注。
2 结果

表2 饲料中发酵豆粕含量对刺参生产性能的影响
由表2可知,本试验中对照组刺参增重率、特定生长率和终末体壁重均小于发酵豆粕试验组,且对照组的刺参增重率、特定生长率和终末体壁重与试验组D12%、D16%和D20%比较差异显著(P<0.05),而各组间刺参体壁系数差异不显著。刺参增重率、特定生长率和终末体壁重都是随着饲料中发酵豆粕含量的升高而先升高后降低,当饲料中发酵豆粕含量提高到16%时,刺参增重率、特定生长率和终末体壁重达到最大值,高于对照组和试验组。结果表明:当饲料中发酵豆粕含量提高到16%时,本试验中刺参各项生产性能指标达到最优。
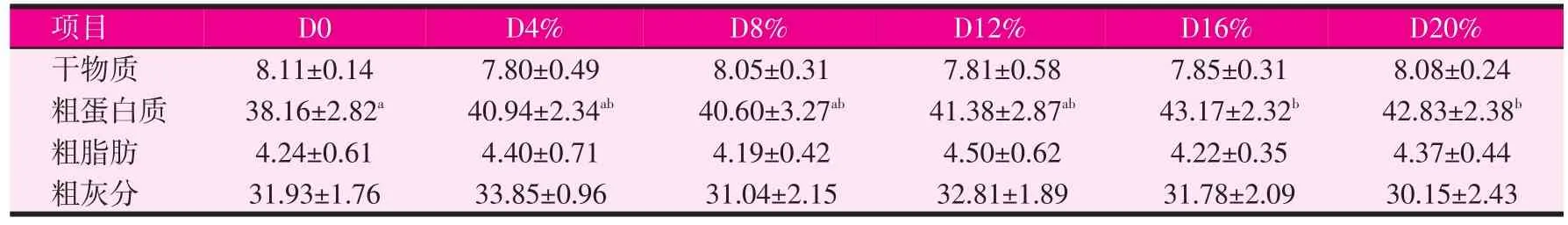
表3饲料中发酵豆粕含量对刺参体壁营养成分的影响(干物质)(%)
由表3可知,与对照组相比,各发酵豆粕试验组对刺参体壁营养成分(干物质、粗脂肪、粗灰分)无显著性影响,但各组间刺参体壁营养成分中粗蛋白质含量存在差异。刺参体壁干物质含量在7.80%~8.11%之间;粗蛋白质含量在38.16%~43.17%之间,发酵豆粕16%试验组刺参粗蛋白质含量最高,豆粕含量20%的对照组刺参粗蛋白质含量最低;粗脂肪含量在4.19%~4.50%之间;粗灰分含量在30.15%~33.85%之间。
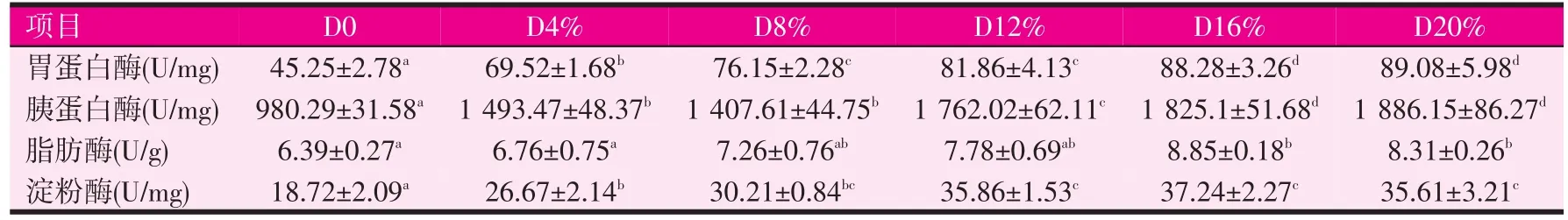
表4 饲料中发酵豆粕含量对刺参消化酶活性的影响
由表4可知,与对照组相比,发酵豆粕显著性影响胃蛋白酶、胰蛋白酶和淀粉酶活性(P<0.05)。试验组中随着发酵豆粕含量的提高,胃蛋白酶和胰蛋白酶活性随之升高,脂肪酶和淀粉酶也有类似的变化趋势。发酵豆粕含量为20%时,刺参胃蛋白酶、胰蛋白酶活性最高,与发酵豆粕含量16%的试验组差异不显著,与其他各组比较差异显著(P<0.05)。豆粕含量20%对照组,刺参胃蛋白酶、胰蛋白酶活性最低,与其它各组比较差异显著。对照组与发酵豆粕4%、8%、12%的试验组之间脂肪酶活性差异不显著,但对照组脂肪酶活性显著性(P<0.05)低于发酵豆粕含量16%、20%的试验组,发酵豆粕16%和20%的试验组间脂肪酶活性差异不显著。
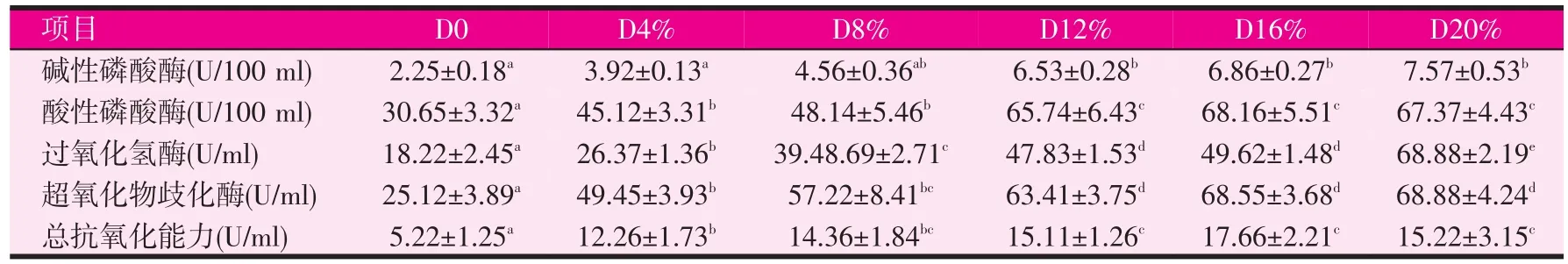
表5 饲料中发酵豆粕含量对刺参体液关键酶活性的影响
由表5可知,本试验中除碱性磷酸酶外,对照组与发酵豆粕各试验组之间酸性磷酸酶、过氧化氢酶、超氧化物歧化酶活性与总抗氧化能力差异显著(P<0.05)。随着发酵豆粕含量的增加碱性磷酸酶、酸性磷酸酶、过氧化氢酶、超氧化物歧化酶活性和总抗氧化能力都表现出升高的趋势。除过氧化氢酶外,当发酵豆粕含量达到16%~20%时,碱性磷酸酶、酸性磷酸酶、超氧化物歧化酶活性与总抗氧化能力升高后趋于一致,二者之间差异不显著(P>0.05),均显著高于对照组和发酵豆粕4%试验组(P<0.05)。发酵豆粕含量达到20%时,过氧化氢酶活性显著高于其它各试验组。豆粕含量20%的对照组的酸性磷酸酶、过氧化氢酶、超氧化物歧化酶活性与总抗氧化能力显著低于其它各试验组。
3 讨论
蛋白质是水产动物生长、繁殖的物质基础,其在刺参营养需要中处于非常重要的地位,饲料中蛋白质的质量影响刺参的摄食、生长、营养物质的消化与吸收等多种生理活动。饲料添加发酵豆粕对改善饲料蛋白质质量有重要作用,可以提供丰富的氨基酸和肽类物质。刺参属于杂食性偏植物性的海洋动物,饲料中同时添加动物性蛋白源鱼粉和植物蛋白源豆粕、发酵豆粕或其它植物饼粕等才能够实现动植物蛋白源优势互补,达到饲料氨基酸相对平衡,促进刺参快速生长的目的。很多试验证明鱼粉含量偏低,对刺参生长有抑制作用[3-5]。本试验中各组饲料中动植物蛋白比为1∶3,鱼粉含量为6.5%,但随着饲料中发酵豆粕含量的升高,鱼粉与发酵豆粕的相对比值也呈现下降趋势(1.63、0.81、0.54、0.41、0.32),发酵豆粕添加量的增加也起到替代鱼粉的作用。在本试验中刺参饲料中发酵豆粕含量对刺参的生长性能有一定的影响,当饲料发酵豆粕含量达到16%时,刺参的特定生长率、增重率、体壁质量达到最大值。沈城等(2015)[6]利用发酵豆粕替代15%鱼粉,中华鳖的增重率和特定生长率均提高显著且达到最大值;杨耐德等(2008)[7]发现凡纳滨对虾养殖实验中发酵豆粕替代鱼粉蛋白超过33.33%,则会降低凡纳滨对虾生长性能和饲料利用率;Lou等(2004)[8]在石斑鱼上也发现用发酵豆粕替代鱼粉比例超过7.4%可抑制生长。水产养殖动物的种类不同,其所需要的必需氨基酸模型不同,配制的饲料中动植物蛋白源鱼粉和发酵豆粕的最佳比例也有很大的差别,特别是发酵豆粕和鱼粉的氨基酸组成差异较大。发酵豆粕做为优质植物蛋白源,虽然氨基酸组成相对平衡,但限制性氨基酸蛋氨酸和赖氨酸远低于鱼粉中的含量,这可能是饲料中发酵豆粕添加量过高,鱼粉蛋白相对含量降低后容易导致饲料利用率降低,影响刺参、鱼虾等水产动物生长的原因。
刺参的可食部分主要是刺参体壁,其营养和保健功能与体壁的质量密切相关。饲料营养成分对刺参体壁质量有一定影响。本试验中饲料中发酵豆粕的含量对刺参体壁中粗蛋白质有一定影响,当饲料中发酵豆粕含量达到16%时,对刺参体壁蛋白质的沉积作用达到最大值。对其它常规营养成分(干物质、粗脂肪、粗灰分)影响不显著。闫磊等(2017)[9]、程成荣等(2004)[10]和Liang等(2017)[11]通过实验在洛氏鱥、杂交罗非鱼和花鲈上证实了类似结果的存在。但是Luo等(2004)[8]、冯建等(2016)[12]、Lee等(2016)[13]和沈城等(2015)[6]研究发现,发酵豆粕替代鱼粉对中华鳖稚鳖、石斑鱼、大黄鱼和许氏平鮋肌肉的粗脂肪、水分、粗蛋白质和粗灰分含量无显著影响。吕云云等(2016)[14]发现发酵豆粕对圆斑星鲽的体成分(干物质、粗蛋白质、粗脂肪、粗灰分)的影响显著(P<0.05)。导致以上差异存在的原因可能是水产养殖动物种类、发酵豆粕质量、饲料中其它原料(如鱼粉、豆粕等)添加量等因素综合作用的结果。
很多学者开展了发酵豆粕对水产养殖动物消化酶活性影响的相关研究,一般认为适量添加发酵豆粕能够提高水产动物的消化酶活性,过量添加则有抑制作用[6,15-17],但对少部分水产养殖动物的蛋白酶的影响不显著,而对淀粉酶的影响显著(P<0.05)[18]。本试验中发酵豆粕(4%~20%)试验组与对照组比较蛋白酶和淀粉酶显著升高,脂肪酶也有升高趋势,但只有发酵豆粕16%和20%的试验组脂肪酶显著性高于对照组。这可能是由于对照组中豆粕含量高,胰蛋白酶抑制因子等抗营养因子较多,抑制消化酶活性,而试验组中发酵豆粕经过酶解和微生物分解,使各种抗营养因子显著性降低或完全去除,并且发酵豆粕中氨基酸和肽类物质可做为底物有诱导消化酶分泌和提高消化酶活性的作用。
刺参属于无脊椎的海洋棘皮动物,不具备脊椎动物所特有的免疫组织和器官,主要通过非特异的细胞免疫和体液免疫完成对病原微生物和其它抗原物质的免疫防御以及完成对体内产生的过氧化物的清除工作,体液免疫反映主要由体腔液中溶菌酶、超氧化物歧化酶、过氧化氢酶、酸性磷酸酶等多种免疫因子共同决定[19-21]。刺参体腔液中主要免疫或抗氧化酶活性是衡量刺参机体生理状态的重要指标。本试验中发酵豆粕试验组的酸性磷酸酶、过氧化氢酶、超氧化物歧化酶活性与总抗氧化能力都显著高于对照组(P<0.05),刺参体腔液中的这些酶类在清除机体氧自由基,防止生物分子损伤方面具有重要作用,是刺参非特异性免疫水平的高低的标志。当发酵豆粕替代鱼粉量增加到25%以后,酸性磷酸酶、碱性磷酸酯酶、超氧化物歧化酶活性明显下降,因此使用发酵豆粕时必须保证鱼粉的最适使用量,否则会对中华鳖的非特异性免疫产生明显的影响[22]。彭松等(2015)[23]利用发酵豆粕替代鱼粉显著性的提高凡纳滨对虾超氧化物歧化酶(SOD)活性,类似的试验结果在许氏平鮋上也被发现。Song等(2014)[24]证实过多添加大豆蛋白水解物降低鱼粉在饲料中的含量则会使星斑川鲽的超氧化物歧化酶活性与总抗氧化能力降低。由以上现象可以得出在刺参、鱼虾等饲料中动物蛋白源鱼粉适量时,添加一定量的发酵豆粕可提高机体的免疫力和抗氧化能力,但具体机理和途径,因水产动物的种类不同差异较大,有待进一步深入研究。
4 结论
在动植物蛋白比为1∶3的刺参饲料中,发酵豆粕添加量达到16%时,刺参生长、体壁蛋白质含量达到最佳,消化酶活性、免疫酶活性都得到提高,具有一定的理论研究价值和生产应用价值。
