维生素E和/或柠檬酸添加剂对军曹鱼稚幼鱼ACO和PPARα基因、抗氧化酶活力和多不饱和脂肪酸代谢的影响
2019-01-02丁兆坤黄金华李伟峰许友卿
■丁兆坤 黄金华 李伟峰 许友卿
(广西大学水产科学研究所广西高校水生生物健康养殖和营养调控重点实验室,广西南宁530004)
军曹鱼是一种生长快、养殖发展迅猛的优良海水鱼[1-4]。然而,为了开发适宜的人工配合饲料,还需要进一步研究军曹鱼的营养生理学[5-8]。
维生素E是军曹鱼和其他鱼类必不可少的微量营养物质[9-10],具有重要的生理生化功能[11-14]。日粮添加维生素E还可以防止鱼体发生脂质过氧化反应,维护鱼体氧化稳定性,降低鱼的氧化应激,提高鱼的免疫力和抗病力,促进鱼生长性能和存活率[15-18]。过氧化物酶体增殖物激活受体α(PPARα)是机体内调控脂质代谢和基因表达的一种基因[19-20]。维生素E是PPARα的一种激动剂,对PPARα的表达有重要影响[7,21-22]。
柠檬酸(CA)能够提高鱼[23-27]和虾[28]的生长性能、饲料利用率和存活率。柠檬酸是一种有机酸,具有多种营养生理功能,是一种环境友好型的饲料添加剂[28-30]。柠檬酸是一种优秀的抗氧化剂,能够有效清除体内外超氧离子和脂质过氧化物,减轻动物的氧化应激[31-32]。柠檬酸也是一种优秀的抗氧化协同剂,能够有效促进抗氧化酶的活力[32-33]。柠檬酸可以通过柠檬酸循环促进机体代谢和顺乌头酸酶(ACO)基因的表达[34]。顺乌头酸酶(ACO)是机体内柠檬酸循环中催化柠檬酸和异柠檬酸相互转化的关键酶[34]。
在日粮中联合添加维生素E和柠檬酸可能对动物的生理生化过程起协同作用[33]。然而,日粮中联合添加维生素E和柠檬酸对军曹鱼稚幼鱼的影响还未见报道。本文研究不同剂量维生素E和/或柠檬酸添加剂对军曹鱼稚幼鱼主要组织/器官的ACO和PPARα基因表达、抗氧化酶活力和多不饱和脂肪酸代谢的影响,同时讨论其机制,为开发适合军曹鱼稚幼鱼配合饲料提供理论依据。
1 材料与方法
1.1 主要饲料原料
维生素E购于美国Sigma-Aldrich公司,是从植物油中提取而得的(±)-α-生育酚,V型,1 000 IU/g(Sigma T3634)。北太平洋白鱼粉(蛋白质>65%)和鱼油,购于美国海鲜公司。其他原料均是人类食品级,购于国内不同生物技术公司。
1.2 实验鱼的驯化和养殖
从湛江流沙海水鱼种苗场购进2 000尾军曹鱼稚鱼,鱼龄为22 d,体重约2.5 g,体长约3.68 cm。先按照Ding等[7]的方法在广西防城港市北部湾海滨室内海水养殖系统驯化2周。驯化结束后,挑选大小一致,体长(8.02±0.32)cm、体重(7.32±0.26)g的健壮稚鱼420尾随机分成7组,每组3个平行,共21桶(每桶容积为400 L),每桶20尾鱼,用与驯化相同的养殖系统和海水(过滤、充气的海水,水温26.0~32.0℃,盐度27~30 g/l,pH值7.8~8.0,溶氧量≥6 mg/l)养殖。按照Ding等[7]方法饲养和处理鱼。每天定时在8:00和17:00投喂两次,分别用7种(组)饲料投喂鱼。这7种(组)饲料分别是:对照组(D0),只含基础饲料,不添加VE和CA;实验饲料D1~D6组,在每千克干的基础饲料中分别添加VE 100、0、100、75、50、25 IU和CA 0、12、12、6、3、1.5 g(见表1)。
基础饲料是在我们前期的试验[3-4,7]基础上设计的,如表1所示,由白鱼粉、豆粕、酪蛋白、大豆磷脂、鱼油、混合维生素(不含VE)、混合矿物质和α-淀粉组成,其蛋白质和脂肪的含量分别约为47%和11%。维生素E的添加量参考了其他学者的研究结果和我们的前期试验[4,7,9-10,17,35]。据报道,维生素 E(α-tocopherol或者DL-all-rac-α-tocopherol)在对照组日粮中的含量为25.70 IU/kg干饲料(1.1 IU DL-all-rac-α-tocopherol相等于1 mg)[36],已经基本满足军曹鱼稚幼鱼生存的需求,因为普通养鱼的VE需求在6.25~200.0 mg生育酚乙酸酯/kg干饲料(1 IU生育酚乙酸酯相等于1 mg)[36]范围之内[9-10]。柠檬酸的添加量参考了Sarker等[37]的研究,他们称在以部分植物蛋白质为蛋白源的鱼类日粮中,高效的柠檬酸添加水平应不超过10.0 g/kg干饲料。

表1 实验饲料的基础成分及营养水平
这个实验设计参考了 Cheng等[38]、Pohlenz等[39]和Ding等[7]的方法,设计不仅考察了单独添加VE(D1组)或CA(D2组)对军曹鱼稚幼鱼的影响,又考察了联合添加VE和CA(D3~D6组)对鱼的影响。
按照Ding等[7]方法,采用丸状日粮投喂鱼,即按照表1的饲料成分、数量、比例和分组进行,在室温下制成丸状日粮,在60℃下烘干,于4℃冰箱中密封保存备用。在投喂之前,取出在室温下解冻待用。日总投喂量约为当时鱼体湿重的5%并要求达到明显的饱感。实验历时12周。
1.3 取样和化学分析
于第12周末,每个养殖桶随机取6尾鱼解剖取组织/器官样品。取样前,鱼禁食24 h。按照Ding等[7]的方法采样和保存所取得的鱼肝、肌肉、脑、心脏、肾和血清样品。
按Xu等[40]的方法,用气相色谱仪(GC,Agilent 6890 N,Agilent Company,USA)测定鱼组织/器官中的脂肪酸含量。分别按照McCord等[41]、Aebi[42]和Noguchi等[43]的方法,测定鱼组织/器官中的超氧化物歧化酶(SOD)、过氧化氢酶(CAT)和谷胱甘肽过氧化物酶(GPx)活力。用南京建成生物公司生产的谷胱甘肽-S转移酶(GST)试剂盒,按照使用说明书,测定鱼组织/器官中的GST活力。
分别按照Peng等[16]和Weikle[44]的方法,用高效液相色谱仪(HPLC,Agilent 1200,Agilent Company,USA)测定饲料中VE和CA含量。
用德国产梅特勒pH计测定饲料pH值。采用AOAC(1995)[45]标准方法测定饲料中的粗蛋白质、粗脂肪、灰分和水分含量。
1.4 鱼组织器官中顺乌头酸酶(ACO)基因序列的测定
军曹鱼的ACO基因序列还未见报道。我们从NCBI(the National Center for Biotechnology Informa-tion)数据库(http://www.ncbi.nlm.nih.gov)搜索到从斑马鱼(Danio rerio)克隆获得的ACO基因的保守序列(NM_198908),以其作为模板合成军曹鱼稚幼鱼的首条cDNA。选择特定正向引物5′CTAGTGCTGGGGCTAAATATGG 3′和 反 向 引 物 5′TGACATTGGGACACTTCAACTC 3′来扩增cDNA片段。按Ding等[7]的方法,测定军曹鱼ACO基因序列。
1.5 ACO mRNAs和 PPARα mRNAs的表达
分别用合成的ACO和PPARα基因首条cDNA为模板,进行实时定量PCR测定。按照上述ACO基因测序所描述的克隆军曹鱼cDNA的方法,设计ACO基因的正向引物和反向引物。
根据已公布的军曹鱼cDNA(http://www.ncbi.nlm.nih.gov)来设计PPARα基因的正向引物和反向引物。分别为ACO和PPARα设计β-肌动蛋白(EU266539)的引物(见表2)。按Ding等[7]的方法进行ACO或者PPARα基因的RTQ-PCR测定。用2-ΔΔCT法计算ACO和PPARα mRNA的相对表达量[46]。

表2 ACO、PPARα和β-actin基因的RTQ-PCR引物
1.6 统计分析
实验数据用“平均值±标准误”表示,用统计软件SPSS18.0对数据进行方差分析,差异显著时采用Duncan′s法进行多重比较,当P<0.05时,为差异显著。
2 结果
2.1 军曹鱼稚幼鱼的ACO基因序列
以斑马鱼ACO基因的保守序列为模板,合成军曹鱼ACO基因的部分DNA保守序列是219 bp。该序列如表3所示,未提交到GenBank基因库。用NCBI BLAST(National Center for Biotechnology Information's Basic Local Alignment Search Tool),把军曹鱼ACO基因的部分DNA保守序列与斑马鱼(Danio rerio)(NM 198908.1)、尼罗罗非鱼(Oreochromis niloticus)(XM 003438531.1)的序列比较,发现军曹鱼ACO基因的部分DNA保守序列与斑马鱼(Danio rerio)(NM 198908.1)和尼罗罗非鱼(Oreochromis niloticus)(XM 003438531.1)的序列相似性分别是86%和90%。

表3 军曹鱼ACO基因部分DNA的保守序列
2.2 用不同剂量VE和/或CA添加剂投喂军曹鱼稚幼鱼12周,对鱼ACO mRNAs相对表达量的影响(见图1)
用不同剂量VE和/或CA添加剂投喂军曹鱼稚幼鱼12周,鱼肝、肌肉、心脏、脑和肾ACO mRNAs相对表达量分别显著高于对照组(D0)(P<0.05)。投喂联合添加VE和CA日粮的鱼(D3~D6组)肝、肌肉、心脏、脑和肾ACO mRNAs的相对表达量,分别显著高于投喂单独添加VE(D1组)或CA(D2组)日粮的鱼相应组织/器官ACO mRNAs的相对表达量(P<0.05)。其中D5组鱼肝、肌肉、心脏、脑和肾ACO mRNAs的相对表达量最高,分别显著高于其他组鱼ACO mRNAs的表达量(P<0.05)。
2.3 用不同剂量VE和/或CA添加剂投喂军曹鱼稚幼鱼12周,对鱼PPARα mRNAs相对表达量的影响(见图2)
用不同剂量VE和/或CA添加剂投喂军曹鱼稚幼鱼12周,鱼肝、肌肉、心脏、脑和肾PPARα mRNAs的相对表达量分别显著高于对照组(D0)(P<0.05)。投喂联合添加VE和CA日粮的鱼(D3~D6组)肝、肌肉、心脏、脑和肾PPARα mRNAs的相对表达量,分别显著高于投喂单独添加VE(D1组)或CA(D2组)日粮的鱼相应组织/器官PPARα mRNAs的相对表达量(P<0.05)。其中D5组鱼肝、肌肉、心脏、脑和肾PPARα mRNAs的相对表达量最高,分别显著高于其他组鱼PPARα mRNAs的表达量(P<0.05)。

图1 用不同剂量VE和/或CA添加剂投喂军曹鱼稚幼鱼12周,对鱼ACO mRNAs表达量的影响
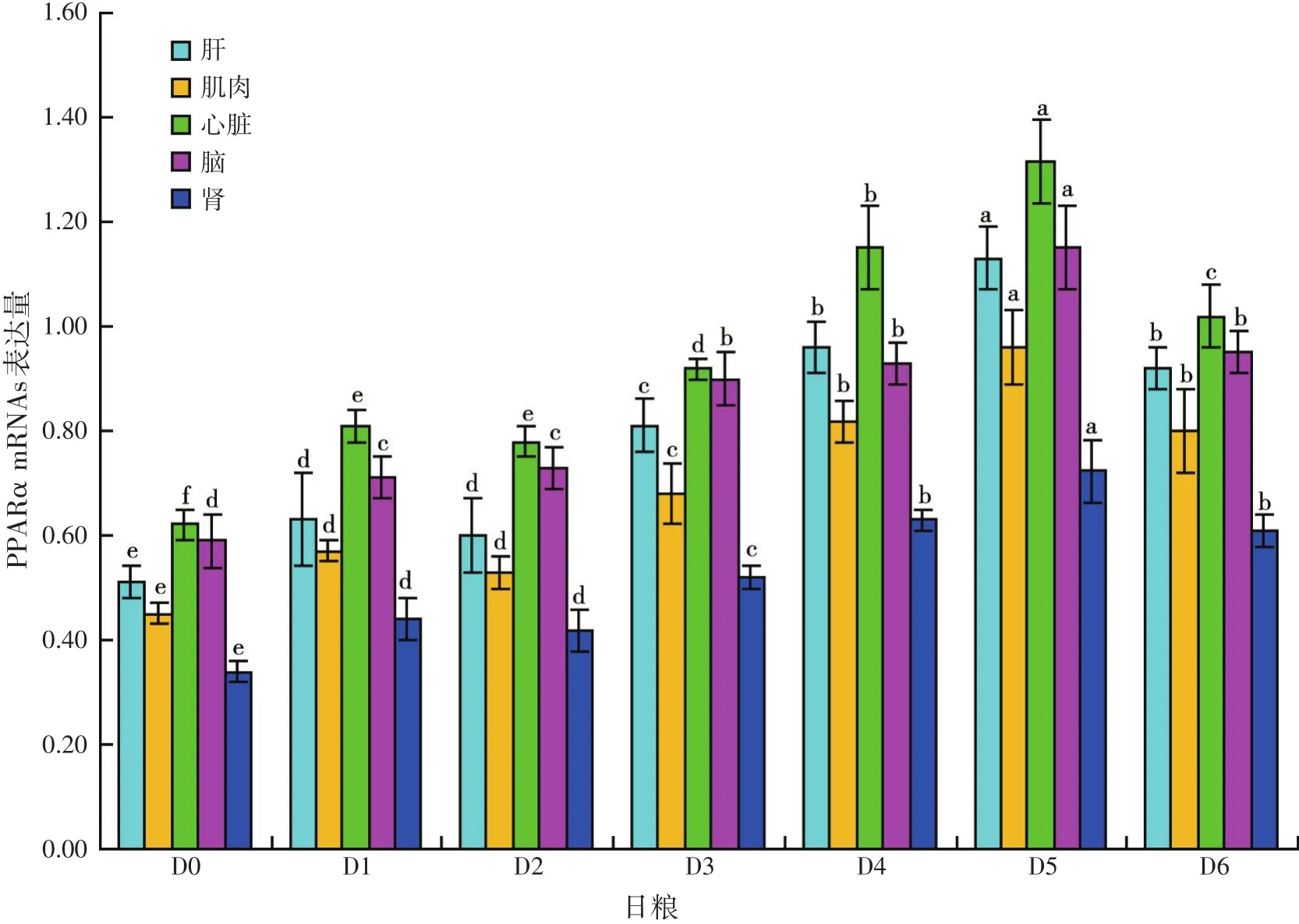
图2 用不同剂量VE和/或CA添加剂投喂军曹鱼稚幼鱼12周,对鱼PPARα mRNAs表达量的影响
2.4 用不同剂量VE和/或CA添加剂投喂军曹鱼稚幼鱼12周,对鱼SOD、CAT、GPx和GST活力的影响(见表4)
由表4可知,用不同剂量VE和/或CA添加剂投喂军曹鱼稚幼鱼12周,鱼肝、肌肉、心脏、脑和肾SOD、CAT、GPx和GST活力分别显著高于对照组(D0)(P<0.05)。投喂联合添加VE和CA日粮的鱼(D3~D6组)肝、肌肉、心脏、脑和肾SOD、CAT、GPx和GST活力,分别显著高于投喂单独添加VE(D1组)或CA(D2组)日粮的鱼相应组织/器官的抗氧化酶活力(P<0.05)。其中D5组鱼肝、肌肉、心脏、脑和肾SOD、CAT、GPx和GST的活力最高,分别显著高于其他组鱼相应组织/器官的抗氧化酶活力(P<0.05)。

表4 不同剂量维生素E和/或柠檬酸添加剂投喂军曹鱼稚幼鱼12周对鱼肝、肌肉、心脏、脑、肾和血清的SOD、CAT、GPx和GST(U/mg prot.,但血清为U/ml)活力的影响
用不同剂量VE和/或CA添加剂投喂军曹鱼稚幼鱼12周,鱼血清SOD、CAT、GPx和GST的活力分别显著高于对照组(D0)(P<0.05)。投喂联合添加VE和CA日粮的鱼(D3~D6组)血清CAT和GST的活力,分别显著高于投喂单独添加VE(D1组)或CA(D2组)日粮的鱼的血清CAT和GST活力(P<0.05)。然而,血清SOD、CAT、GPx和 GST的最高活力不同于肝、肌肉、心脏、脑和肾的结果。D5组鱼血清SOD活力最高,显著高于其他组鱼血清SOD活力(P<0.05)。D4组鱼血清CAT活力最高,显著高于其他组鱼血清CAT活力(P<0.05)。D6组鱼血清GPx活力最高,显著高于D0、D1~D4组鱼血清GPx活力(P<0.05)。D4组鱼的血清GST活力最高,显著高于D0、D1~D3组和D6组鱼血清GST活力(P<0.05)。
2.5 用不同剂量VE和/或CA添加剂投喂军曹鱼稚幼鱼12周,对鱼脂肪酸含量的影响(见表5~表10)
用不同剂量VE和/或CA添加剂投喂军曹鱼稚幼鱼12周,显著影响其多不饱和脂肪酸代谢,鱼肝、肌肉、心脏、脑和肾PUFAs和n-3 HUFAs分别显著高于对照组(D0)(P<0.05)。投喂联合添加VE和CA日粮的鱼(D3~D6组)肝、肌肉、心脏、脑和肾PUFAs和n-3 HUFAs,分别显著高于投喂单独添加VE(D1组)或CA(D2组)日粮的鱼相应组织/器官的PUFAs和n-3 HUFAs(P<0.05)。其中D5组的鱼肝、肌肉、心脏、脑和肾PUFAs和n-3 HUFAs最高,分别显著高于其他组鱼相应组织/器官的PUFAs和n-3 HUFAs(P<0.05)。
不同剂量VE和/或CA添加剂对鱼血清中的脂肪酸代谢的影响不同于对鱼肝、肌肉、心脏、脑和肾脂肪酸代谢的影响。D4组鱼血清的PUFAs和n-3 HUFAs含量最高,显著高于D0、D1~D3、D5组和D6组鱼血清的PUFAs和n-3 HUFAs含量(P<0.05)。
3 讨论
与单独添加VE(D1组)或CA(D2组)或对照组(D0)比较,用联合添加VE和CA(D3~D6组)的日粮投喂军曹鱼稚幼鱼12周,显著提高鱼肝、肌肉、心脏、脑和肾ACO和PPARα基因的表达量、SOD、CAT、GPx和GST活力以及PUFAs和n-3 HUFAs的含量。这些结果表明,添加在日粮中的VE和CA能协同作用,显著提高军曹鱼稚幼鱼ACO和PPARα基因表达、抗氧化酶活力以及PUFAs和n-3 HUFAs的含量。VE和CA协同作用的原理可能是:①VE和CA能够协同调节某些基因的表达、提高抗氧化酶的活力。研究表明,VE是PPARα基因的激活剂,能够促进PPARα基因的表达[7,21-22]。VE 能够显著提高鱼体中的抗氧化酶活力[7,17,47-48]。而
CA给VE提供了一个适宜的酸性环境,促进VE发挥更好的作用。②VE通过激活脂肪丙烯醛基去饱和并延长之,促进PUFAs合成,同时保护PUFAs免受过氧化。有研究报道,饲料添加VE可显著提高鱼肝和肌肉PUFAs含量而显著降低SFAs含量[49-50]。③VE和CA都是高效抗氧化剂,它们通过分解和消除军曹鱼代谢过程中产生的活性氧自由基,来协同保护和促进军曹鱼的生理生化过程。VE和CA都能够通过有效地清除超氧阴离子(·O2-)来协同抑制和切断脂质过氧化反应[51-53]。VE提供H原子给脂质过氧物自由基(LOO·)生成脂质过氧化物LOOH,以切断脂质过氧化自由基链式反应[51]。脂质过氧化物LOOH是一种氧化性非常强的物质,在有金属离子中介的情况下,进一步分解为自由基LOO·、LO·和·OH[33],而VE不能进一步清除LOOH,必须由其他抗氧化剂来清除LOOH[51]。当日粮同时添加VE和CA时,CA能够抑制LOOH生成[33]。尽管CA不是主要的抗氧化剂,但是CA是一个良好的抗氧化剂协同剂。VE和CA也可能互相再生来扩增它们的功能[33]。因此,在日粮中联合添加VE和CA能够协同提高军曹鱼的ACO和PPARα基因的表达、抗氧化酶活力、PUFAs和n-3 HUFAs含量。

表5 不同剂量维生素E和/或柠檬酸添加剂投喂军曹鱼稚幼鱼12周对鱼肝脏脂肪酸的影响(%)

表6 不同剂量维生素E和/或柠檬酸添加剂投喂军曹鱼稚幼鱼12周对鱼肌肉脂肪酸的影响(%)

表7 不同剂量维生素E和/或柠檬酸添加剂投喂军曹鱼稚幼鱼12周对鱼心脏脂肪酸的影响(%)
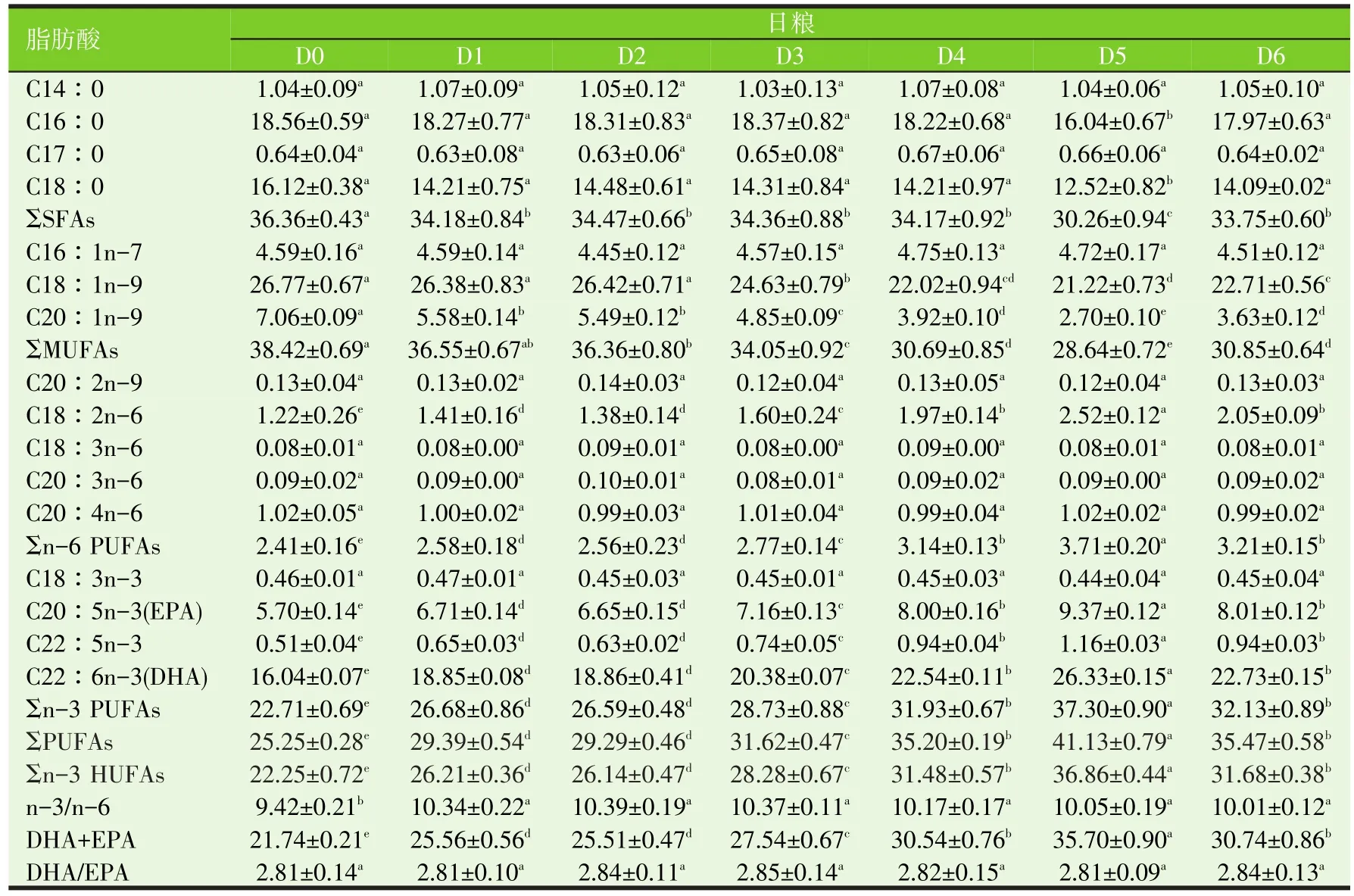
表8 不同剂量维生素E和/或柠檬酸添加剂投喂军曹鱼稚幼鱼12周对鱼脑脂肪酸的影响(%)
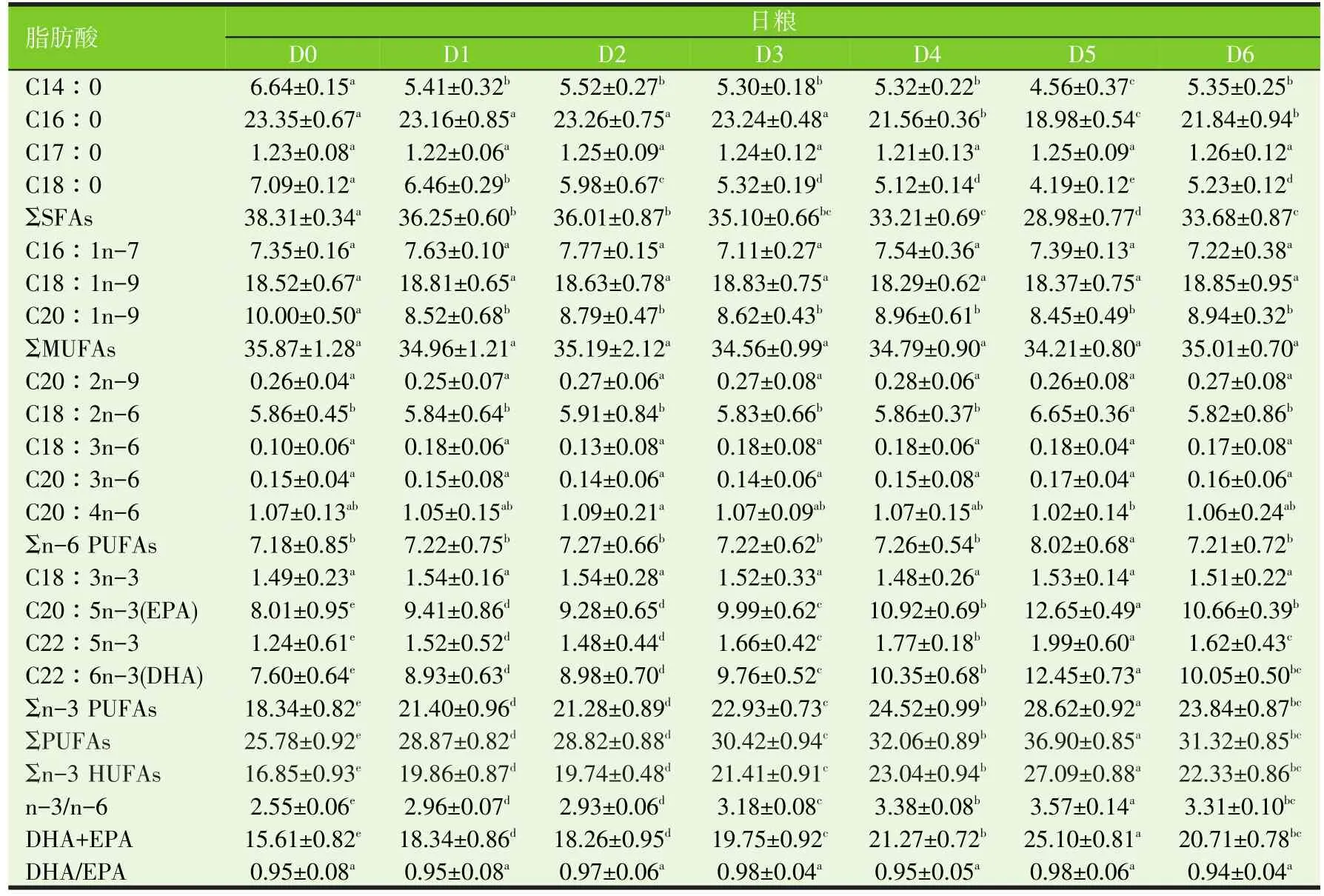
表9 不同剂量维生素E和/或柠檬酸投喂军曹鱼稚幼鱼12周对鱼肾脂肪酸的影响(%)

表10 不同剂量维生素E和/或柠檬酸投喂军曹鱼稚幼鱼12周对鱼血清脂肪酸的影响(%)
尽管D3、D4组和D6组的日粮也联合添加了VE和CA,但是D5组鱼的效果最佳,D5组鱼的ACO和PPARα基因表达、抗氧化酶活力,以及PUFAs和n-3 HUFAs含量显著高于D3、D4组和D6组的鱼。这些结果表明:日粮中联合添加较高或较低的VE和CA的添加量(D3、D4组和D6组)的效果都不理想。因此,在日粮中适宜添加VE和CA是有益的。VE和CA的协同作用是剂量依赖性的,但有一个适宜的范围。本实验D5组的VE和CA添加量(VE 50 IU和CA 3 g/kg干饲料)最适宜,因此发挥了最佳的协同作用。以前对罗非鱼(Oreochromis niloticus×O.aureus)[54]、鲫鱼(Carassius auratusgibelio)[55]和凡纳滨对虾(Litopenaeus vannamei)[28]等的研究也表明,CA的适宜添加量为2.0~3.0 g/kg干饲料。蔡超等[56]也认为,在鱼饲料中添加CA不宜超过0.5%。
与对照组(D0)比较,单独添加VE(D1组)能够显著提高军曹鱼肝、肌肉、心脏、脑和肾ACO和PPARα基因表达量、SOD、CAT、GPx和GST的活力以及PUFAs和n-3 PUFAs的含量。这个结果与Kim等[57]的试验结果一致,他们在雄性小鼠日粮中添加VE提高了小鼠肝脏中的PPAR-α基因表达量。Zhou等[17]、Lu等[48]和Ding等[7]也报道,日粮中添加VE能够提高抗氧化酶的活力。这是因为VE能够降低氧化应激。当活性氧自由基过度累积时,PUFAs和抗氧化酶的活力会受到抑制[58],而VE能够有效地清除活性氧自由基[51]。
单独添加CA(D2组)也能够显著提高军曹鱼肝脏、肌肉、心脏、脑和肾ACO和PPARα基因表达量、SOD、CAT、GPx和GST的活力以及PUFAs和n-3 PUFAs的含量。理由可能是:CA能够提高抗应激能力和抗氧化作用。作为一个高效抗氧化剂,CA能够有效地清除超氧离子和螯合金属离子来切断脂质过氧化物自由基链式反应[59-60],提高抗氧化能力。CA也是一种优秀的抗氧化协同剂,能够有效促进抗氧化酶的活力[32-33]。Su 等[28]报道,日粮添加CA 2.0~5.0 g/kg干饲料,能够显著提高白虾血清中的SOD活力。
本实验发现,饲料营养成分显著影响着军曹鱼肝、肌肉、心脏、脑、肾和血清的生理生化过程。然而,不同剂量VE和/或CA添加剂对军曹鱼血清中的抗氧化酶的活力、PUFAs含量的影响不同于对军曹鱼肝、肌肉、心脏、脑和肾相应参数的影响。原因是:血清是鱼体内营养物质不断发生变化的一种介质,血清酶的含量和功能会随着许多因素如生理、营养、时间、温度和其他环境因素的变化而变化[61],不像肝、肌肉、心脏、脑和肾那样相当稳定[7]。
4 结论
本研究发现,在本实验条件下,日粮中添加不同剂量的VE和/或CA添加剂能够显著提高军曹鱼稚幼鱼的ACO和PPARα基因表达量、抗氧化酶活力、PUFAs和n-3 HUFAs含量。联合添加VE和CA具有协同作用,显著高于单独添加VE(D1组)或CA(D2组)者。最佳添加量是VE 50 IU和CA 3 g/kg干饲料。
