光照强度对大豆叶片光合特性及同化物的影响
2018-11-29程亚娇范元芳谌俊旭王仲林谭婷婷李佳凤李盛蓝杨文钰
程亚娇 范元芳 谌俊旭 王仲林 谭婷婷 李佳凤 李盛蓝 杨 峰 杨文钰
光照强度对大豆叶片光合特性及同化物的影响
程亚娇 范元芳 谌俊旭 王仲林 谭婷婷 李佳凤 李盛蓝 杨 峰*杨文钰
四川农业大学农学院 / 作物带状复合种植工程技术研究中心, 四川成都 611130
间套作荫蔽环境下, 光强是影响作物生长和产量的直接因素。本试验通过设定不同光照强度, 分析不同大豆品种的光合特性、光合同化物和叶片中叶绿体超微结构的昼夜变化, 明确光强对大豆叶片结构和同化物积累的影响, 了解品种间差异, 以期为提高大豆产量、改善大豆品质提供理论依据。以耐阴品种南豆12和敏感品种桂夏3号为试验材料, 分别设置CK (正常光照, 遮光度0)、A1 (一层黑色遮阳网, 遮光度10%)、A2 (两层黑色遮阳网, 遮光度36%) 3个不同光照强度。结果表明, 随着遮阴程度的增加, 2个大豆品种净光合速率、气孔导度、蒸腾速率和生物量均呈逐渐降低趋势, 但胞间二氧化碳浓度、叶绿素含量逐渐上升。同一大豆品种在相同处理下叶片蔗糖和淀粉含量昼夜变化差异显著。正常光照和A1处理下的大豆叶片蔗糖含量昼夜变化呈双峰曲线, 波峰分别出现在16:00和次日6:00, A2处理下南豆12蔗糖含量昼夜变化呈单峰趋势, 峰值出现在16:00, 为32.80 μg g–1。淀粉含量昼夜变化呈单峰曲线, 最高值均出现在21:00, 且耐阴品种大豆在A2处理下蔗糖和淀粉含量日变化幅度大于敏感品种大豆。大豆叶片超微结果表明, 弱光处理下大豆叶片叶绿体结构完整, 无破碎现象, 相同品种在同一处理下淀粉粒与叶绿体截面积比值昼夜变化显著, 均呈先增加后降低的变化趋势, 耐阴品种变化幅度大于敏感品种, 最大值均出现在21:00。因此, 随着遮阴程度增加, 大豆叶片生物量积累减少, 光合作用降低, 但耐阴品种在适度遮阴下通过调节光合器官结构的昼夜变化而保持良好的光合作用, 以更好地适应荫蔽环境。
大豆; 光照强度; 光合特性; 同化物; 叶绿体超微结构
大力发展玉米大豆套作种植模式, 有利于提高土地利用率, 扩大大豆生产, 改善大豆品质[1]。在这种种植模式下, 玉米株型的不同导致大豆冠层光照强度不同, 从而影响大豆的生长发育[2]。紧凑型玉米对大豆遮阴程度较轻, 大豆生长表现为植株较矮、茎秆较粗; 而松散型玉米对大豆遮阴程度严重, 大豆表现为株高增长过快、茎叶生长不良、主茎柔弱, 容易倒伏[3]。同时, 低光强使大豆光合能力降低, 干物质积累减少, 茎秆中物质积累比例增加[4-5], 叶绿素含量增加, 叶绿素/降低[5-7], 叶绿体面积减小、数量增多, 嗜锇颗粒增多[8]。而遮阴大豆植株在干旱条件下, 单株叶面积、比叶面积、叶绿素、叶绿素和类胡萝卜素含量、叶片相对含水量、净光合速率、气孔导度、胞间CO2浓度、蒸腾速率、光化学猝灭率和电子传递速率均有不同程度的提高[9]。此外, 不同品种大豆耐阴性对弱光的响应程度不同[10-11], 耐阴品种能够通过调节叶片形态以适应环境变化, 从光能截获和利用方面均表现出了对弱光环境的较强适应能力[12], 而敏感品种大豆光合作用降低、干物质积累减少[5]。目前, 围绕弱光环境下大豆叶片光合特性及干物质积累研究较多, 但对弱光环境下大豆叶片光合同化物积累的昼夜变化趋势以及叶绿体超微结构对不同光照强度响应的昼夜变化趋势尚不明确。本研究人工模拟不同光照强度环境,研究不同耐阴性大豆品种的光合特性以及光合同化物和叶绿体超微结构的昼夜变化, 试图为发展间套作、改善田间配置、提高大豆产量提供理论依据。
1 材料与方法
1.1 试验材料
选择2个耐阴性存在明显差异的大豆品种南豆12 (耐阴品种)和桂夏3号(敏感品种), 由四川农业大学农学院提供材料。
1.2 试验设计
采取二因素完全随机设计, 2个因素分别为不同耐阴性大豆品种和不同光照强度, 用黑色遮阳网人工模拟不同光照强度, 设CK (正常光照, 遮光度为0)、A1 (一层黑色遮阳网遮阴, 遮光度为10%)、A2 (两层黑色遮阳网遮阴, 遮光度为36%) 3个处理(图1)。每个处理4盆, 重复3次。

图1 不同处理下大豆冠层光照强度
图中小写字母表示0.05水平下的显著性差异。CK: 正常光照, 遮光度0; A1: 一层黑色遮阳网, 遮光度10%; A2: 两层黑色遮阳网, 遮光度36%。
Bars superscripted by different letters are significant by different at0.05. CK: normal light, shading 0; A1: black shading net, shading 10%; A2: two black shading nets, shading 36%.
于 2017 年在四川农业大学成都校区进行盆栽种植, 7月23日播种大豆, 2行5穴播种, 出苗后间苗, 每穴保留2株, 在花盆上方2 m处覆盖遮阳网, 模拟不同光照强度环境, 定期管理。
1.3 测定项目及方法
1.3.1 冠层光照强度 在大豆五节期(V5), 用美国Spectrum 3412红外/远红外辐射计于大豆冠层上方5 cm处从左往右依次选3个点测定每个处理下的光量子通量密度, 根据公式遮光度=(0–)/0计算遮光度[13-14],0表示正常光照下的光合有效辐射,表示不同处理下遮阳网内光合有效辐射。间隔2 h测定1次并记录PPFD (photosynthetic photon flux density, 光量子通量密度)数值。同时, 在大豆V5期, 从每个处理选取长势均匀一致的大豆幼苗5株测定相关指标, 重复3次。
1.3.2 光合特性 在上午9:00–11:00, 用Li-6400便携式光合仪(美国Li-COR公司)测定大豆功能叶(倒三叶中间的小叶)的净光合速率(photosynthetic rate,n)、气孔导度(stomatal conductance,s)、胞间二氧化碳浓度(intercellular CO2concentration,i)、蒸腾速率(transpiration rate,r), 计算其平均值为各光合参数数值[7]。
1.3.3 光合色素含量 用孔径为14.22 mm的打孔器在大豆功能叶上打2个孔, 剪成丝状, 放入10 mL 80%丙酮溶液中浸提24 h至叶片变乳白色, 分别在波长663 nm、645 nm、470 nm下测定溶液吸光度, 重复3次, 计算叶绿素、叶绿素、类胡萝卜素的含量, 求平均值为其光合色素含量[15-16]。
1.3.4 生物量 将大豆植株各器官分装在不同信封中, 105℃杀青30 min, 80℃烘干至恒重, 用万分之一分析天平准确称量。
1.3.5 光合同化物 分别在9:00、16:00、21:00、次日6:00选取长势均匀一致的大豆功能叶片, 105℃杀青30 min, 80℃烘干至恒重, 用粉样机粉碎。测定蔗糖和淀粉含量, 在每个时间点设置3次重复[17-18]。
1.3.6 叶绿体超微结构 为了研究正常光照和弱光对叶绿体超微结构的影响, 分别在9:00、16:00、21:00、次日6:00选取CK和A2处理下长势均匀的大豆幼苗3株。参考范元芳等[8]的方法, 在靠近主叶脉基部的1/3处取样(5 mm × 5 mm左右), 用3%戊二醛预固定, 之后用1%四氧化锇再固定, 丙酮逐级脱水, Epon 812包埋, 半薄切片光学定位, 超薄切片, 醋酸铀及枸橼酸铅双重染色。用日立H-600IV 型透射电镜观察拍照。观测每个处理各放大倍数下(5000倍、10000倍、15000倍) 3个视野。
1.4 数据分析与处理
采用Microsoft Excel 2016整理试验数据和作图, SPSS 17.0分析数据。
2 结果与分析
2.1 不同光照强度对大豆叶片光合特性的影响
光合能力是作物产量和品质形成的基础[21]。由表1可知, 遮阴处理下大豆净光合速率、气孔导度、蒸腾速率逐渐降低, 胞间二氧化碳浓度逐渐增加。与CK相比, A1、A2处理下南豆12和桂夏3号净光合速率分别降低3.15%、33.33%和12.93%、45.02%。在A2处理条件下, 南豆12的净光合速率、蒸腾速率降低幅度小于桂夏3号, 胞间二氧化碳浓度增加幅度大于桂夏3号, 而南豆12气孔导度变化差异不显著, 桂夏3号气孔导度变化差异显著。说明不同光照强度环境下, 耐阴品种对弱光适应能力高于敏感品种。
2.2 不同光照强度对大豆叶片光合色素含量的影响
由表2可知, 随着遮阴程度的增加, 2个品种大豆叶绿素含量、类胡萝卜素含量和叶绿素的值均呈先增加后减小的趋势, 表现为A1>CK>A2, 叶绿素含量逐渐增加。与CK相比, A1处理下的南豆12和桂夏3号叶绿素含量分别显著增加54.06%、27.94%, 类胡萝卜素含量分别增加25.73%、8.79%, 表明轻度遮阴可能有利于叶绿素和类胡萝卜素含量的增加。与CK相比, A1、A2处理下的南豆12和桂夏3号的叶绿素含量逐渐增加, 同一品种间差异不显著(>0.05), 表明遮阴环境下作物通过增加叶绿素含量增强光能捕获效率。

表1 不同光照强度对大豆叶片光合特性的影响
标以不同小写字母的值在0.05水平差异显著。CK: 正常光照, 遮光度0; A1: 一层黑色遮阳网, 遮光度10%; A2: 两层黑色遮阳网, 遮光度36%。
Values followed by different letter are significant by different at0.05. CK: normal light, shading 0; A1: black shading net, shading 10%; A2: two black shading nets, shading 36%.
2.3 不同光照强度对大豆生物量的影响
随着遮阴程度的增加, 同一品种大豆的生物量逐渐降低, 茎叶比逐渐增加, 品种间差异显著(<0.05)。与CK相比, 南豆12和桂夏3号在A1、A2处理下生物量分别降低了23.88%、32.20%和15.88%、21.36%, 茎叶比分别增加了4.07%、79.13%和9.88%、37.17% (表3)。表明在遮阴环境下敏感品种茎秆物质积累量显著增加, 耐阴品种茎秆生物量积累也显著增加, 但后者高于前者。
2.4 不同光照强度对大豆叶片光合同化物的影响
由图2可知, 随着遮阴程度的增加, 大豆叶片蔗糖和淀粉含量呈先降低后增加的趋势, 表现为A2>CK>A1, 且同一品种蔗糖和淀粉含量昼夜变化差异显著(<0.05)。CK和A1处理下大豆蔗糖含量昼夜变化呈双曲线趋势, 分别在16:00和次日6:00出现峰值; 与CK峰值比较, A1处理下南豆12蔗糖含量峰值降低了3.92%和6.29%, 桂夏3号蔗糖含量峰值升高了0.13%和4.33%, A2处理下南豆12蔗糖含量呈单峰曲线, 最大值为32.80 mg g–1。不同处理下2个品种大豆淀粉含量昼夜变化均呈单峰曲线, 21:00积累量较多, 并与蔗糖含量的昼夜变化呈互补趋势。A2处理下2个品种大豆淀粉积累量在21:00最多, 与CK相比, 南豆12和桂夏3号淀粉积累量分别增加了52.62%、86.44%, 表明适度遮阴有利于大豆叶片光合产物的积累, 形成适应弱光环境的物质基础。
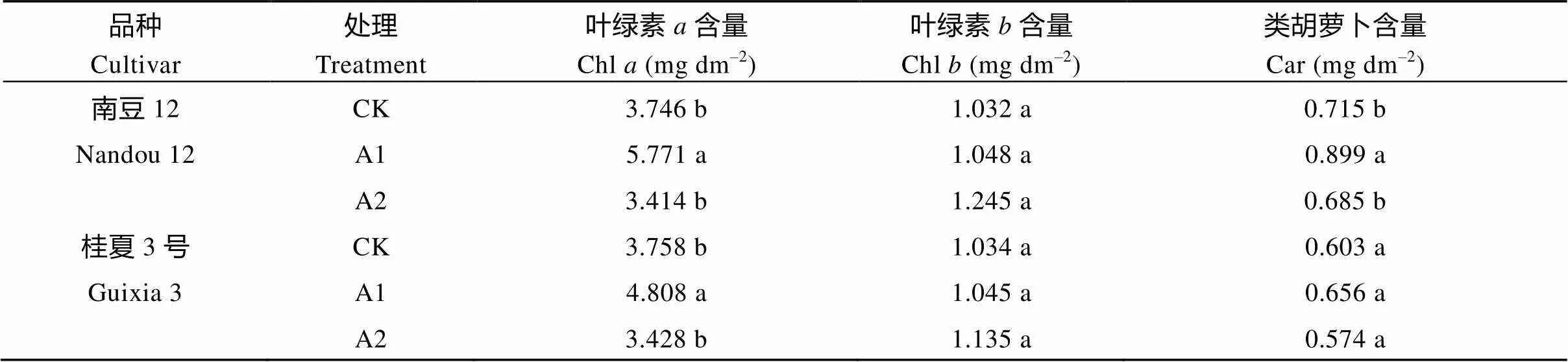
表2 不同光照强度对大豆叶片光合色素含量的影响
标以不同小写字母的值在0.05水平差异显著。CK: 正常光照, 遮光度0; A1: 一层黑色遮阳网, 遮光度10%; A2: 两层黑色遮阳网, 遮光度36%。
Values followed by different letter are significant by different at0.05. CK: normal light, shading 0; A1: black shading net, shading 10%; A2: two black shading nets, shading 36%.
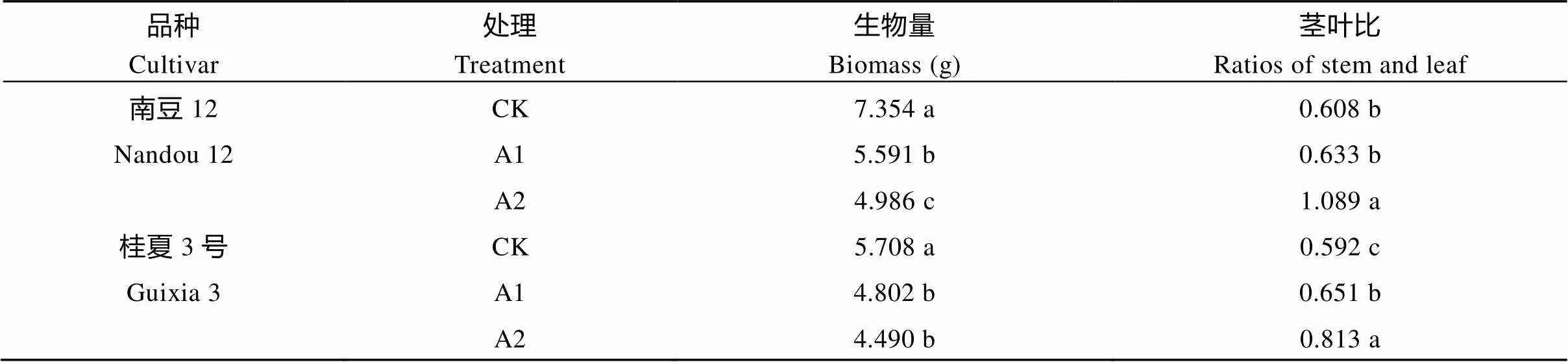
表3 不同光照强度对大豆生物量积累的影响
标以不同小写字母的值在0.05水平差异显著。CK: 正常光照, 遮光度0; A1: 一层黑色遮阳网, 遮光度10%; A2: 两层黑色遮阳网, 遮光度36%。
Values followed by different letter are significant by different at0.05. CK: normal light, shading 0; A1: black shading net, shading 10%; A2: two black shading nets, shading 36%.

图2 不同光照强度对大豆叶片蔗糖、淀粉含量积累的昼夜变化影响
图中小写字母表示同一处理同一品种不同时间点的显著性差异大小(0.05)。CK: 正常光照, 遮光度0; A1: 一层黑色遮阳网, 遮光度10%; A2: 两层黑色遮阳网, 遮光度36%。
Lowercase letters in the figure indicate the significant differences in the same treatment and the same cultivar at different time points. Starch or sucrose content with the same letter did not different at0.05.CK: normal light, shading 0; A1: black shading net, shading 10%; A2: two black shading nets, shading 36%.
2.5 不同光照强度对大豆叶片叶绿体超微结构的影响
为了进一步分析弱光对叶绿体超微结构影响, 选择A2处理与CK处理对比分析。由图3可知, 同一品种在相同处理下大豆叶绿体及淀粉粒形状完整,叶绿体面积及淀粉粒数量昼夜变化显著。为进一步量化淀粉粒和叶绿体变化趋势, 测定了同一品种同一时间点在相同倍镜下的淀粉粒与叶绿体截面积比值。由图4-A可知, 不同处理下南豆12淀粉粒与叶绿体截面积比值昼夜变化趋势相同, 均呈现先增加后降低的趋势, 同一处理不同时间点淀粉粒与叶绿体截面积比值差异显著(<0.05)。与CK比较, A2处理下南豆12在9:00、16:00、21:00、次日6:00分别显著增加了0.72%、8.72%、14.01%和9.04%。桂夏3号淀粉粒与叶绿体截面积比值与南豆12变化趋势基本相同, A2处理较CK在9:00、16:00、21:00、次日6:00分别显著增加了0.83%、8.27%、21.56%和2.18% (图4-B)。
3 讨论
光强是影响大豆生长发育及产量的重要因素。本试验结果表明, 随着遮阴程度增加, 大豆冠层光照强度减弱, 导致大豆净光合速率、气孔导度、蒸腾速率逐渐降低, 胞间二氧化碳浓度逐渐增大, 这可能是由于弱光环境下植株通过调整气孔开放程度,降低蒸腾速率来适应弱光环境, 从而导致光合作用减弱, 减少能量的耗散。光合色素是植物吸收与利用光能的主要色素, 其含量的变化说明植物光合初始反应的优劣[19], 类胡萝卜素与叶绿素、蛋白质构成聚光色素复合体和反应中心复合体, 是植物光合作用吸收光能的重要辅助色素, 光强的变化导致其含量不断的变化[20]。本试验中随着光强降低, 叶绿素含量逐渐增加, 叶绿素和类胡萝卜素含量呈先增加后降低的趋势, 表明弱光环境下叶绿素含量的增加, 有利于捕光色素复合体含量的提高[6]、调节激发能在光系统间的分配[21], 同时不同光敏感型大豆叶片叶绿素、叶绿素含量变化的差异性说明了品种抵御弱光的能力不同[22], 这与王一等[23]的研究结果一致。同时, 在适度遮阴的条件下, 2个大豆品种光合作用均降低, 但耐阴品种降低程度低于敏感品种仍保持较强的光合作用[5], 这与前人研究荫蔽环境对大豆幼苗光合特性影响的结果一致[6-7,24]。
在弱光条件下, 植物通过增加叶片生物量分配和形成大而薄的叶面积提高光截留率[25], 本试验结果表明, 随着遮阴程度的增加, 大豆生物量逐渐降低, 茎叶比逐渐增大, 是由于荫蔽环境下大豆光合作用减弱, 导致生物量积累减少(表3), 植株可通过调节体内物质分配来增强对环境的适应性。大豆植株的茎秆伸长生长可获取更多的光能, 茎秆中生物量积累较叶片生物量增大, 且在适度遮阴环境下强耐阴品种生物量积累高于弱耐阴品种[5]。
蔗糖作为信号提高源-库强度从而提高作物产量[26]。本研究表明, 光强降低, 大豆叶片蔗糖含量降低, 但在适度遮阴下蔗糖含量增加。这是由于遮阴环境下大豆光合作用降低, 导致蔗糖含量降低, 同时蔗糖作为分子信号, 其含量的不足促进叶片光合作用循环不断地进行[26]。因此, 在适度遮阴环境下蔗糖含量的不足促进蔗糖再生, 并在植物体内积累。蔗糖是碳水化合物运输和贮藏的主要形式, 在植物糖代谢中具有特殊的位置, 蔗糖代谢和淀粉合成是2个既相互依赖, 又相互制约的过程[26-28]。本研究中随着光照强度的降低, 淀粉积累量降低, 但在适度遮阴下淀粉积累量增加, 且淀粉含量日积累随着一天光强降低而逐渐增加, 与蔗糖含量变化呈互补的趋势。耐阴品种蔗糖和淀粉含量高于敏感品种, 且昼夜积累波动幅度较大, 可能是其光合作用强于敏感品种, 且在弱光环境下, 蔗糖较多地转变成淀粉贮存在植物体中, 是保证生长的物质基础。
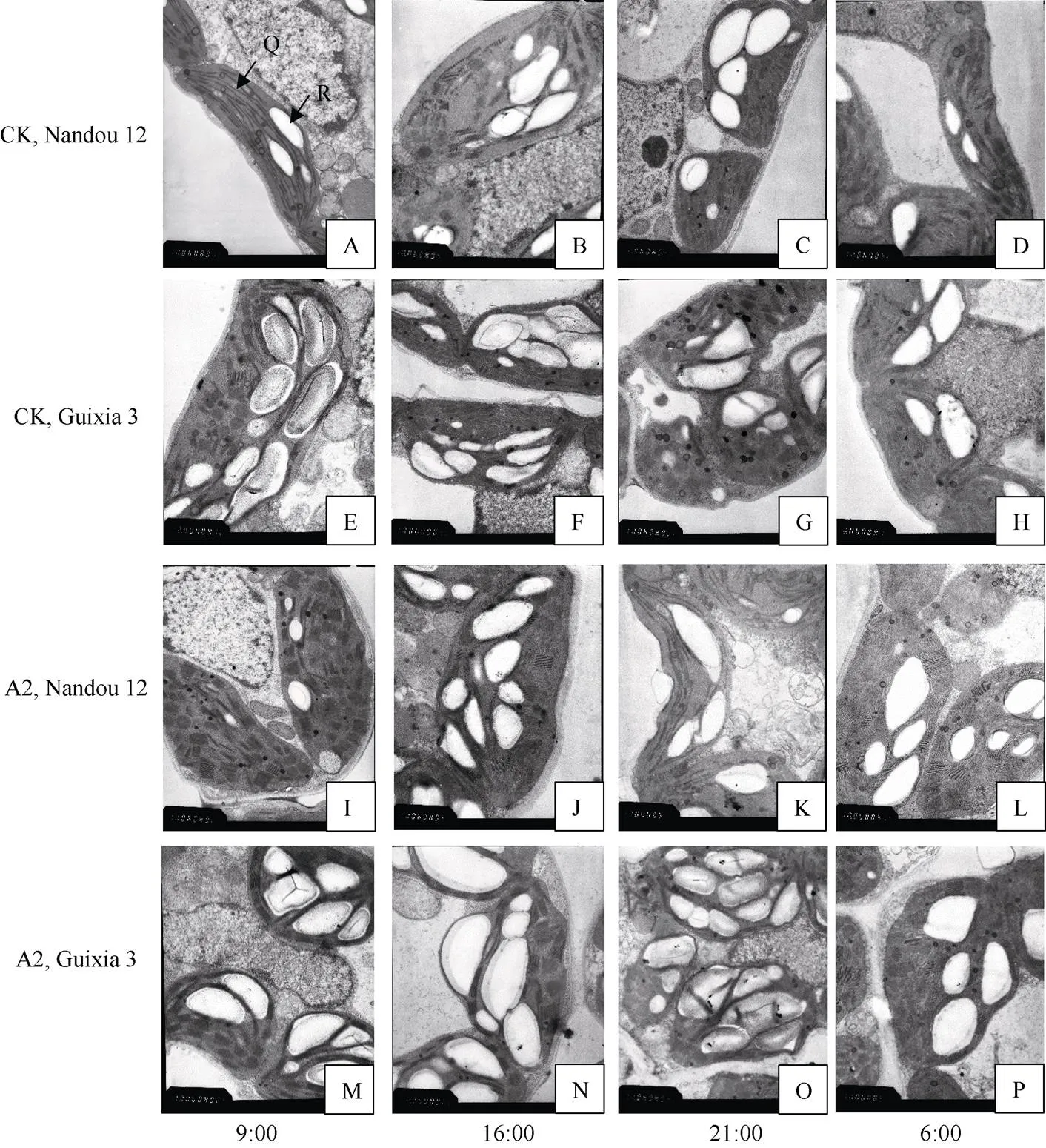
图3 不同光照强度对大豆叶片叶绿体超微结构的影响
上图为10000倍镜下叶绿体超微结构, A~D为正常光照下南豆12叶绿体超微结构在9:00、16:00、21:00、次日6:00的昼夜变化, E~H为正常光照下桂夏3号的叶绿体超微结构的昼夜变化, I~L为A2处理下南豆12的叶绿体超微结构的昼夜变化, M~P为A2处理下桂夏3号的叶绿体超微结构的昼夜变化。Q: 叶绿体, R: 淀粉粒。
The figure shows the chloroplast ultrastructure under 10000 magnifications. A–D is the diurnal variation of chloroplast ultrastructure of Nandou 12 under normal light at 9:00, 16:00, 21:00, and the next day 6:00, E–H is the diurnal variation of chloroplast ultrastructure of Guixia 3 under normal light, I–L is the diurnal variation of chloroplast ultrastructure of Nandou 12 under A2 treatment, and M–P is the diurnal variation of chloroplast ultrastructure of Guixia 3 under A2 treatment. Q: chloroplast, R: starch grains.
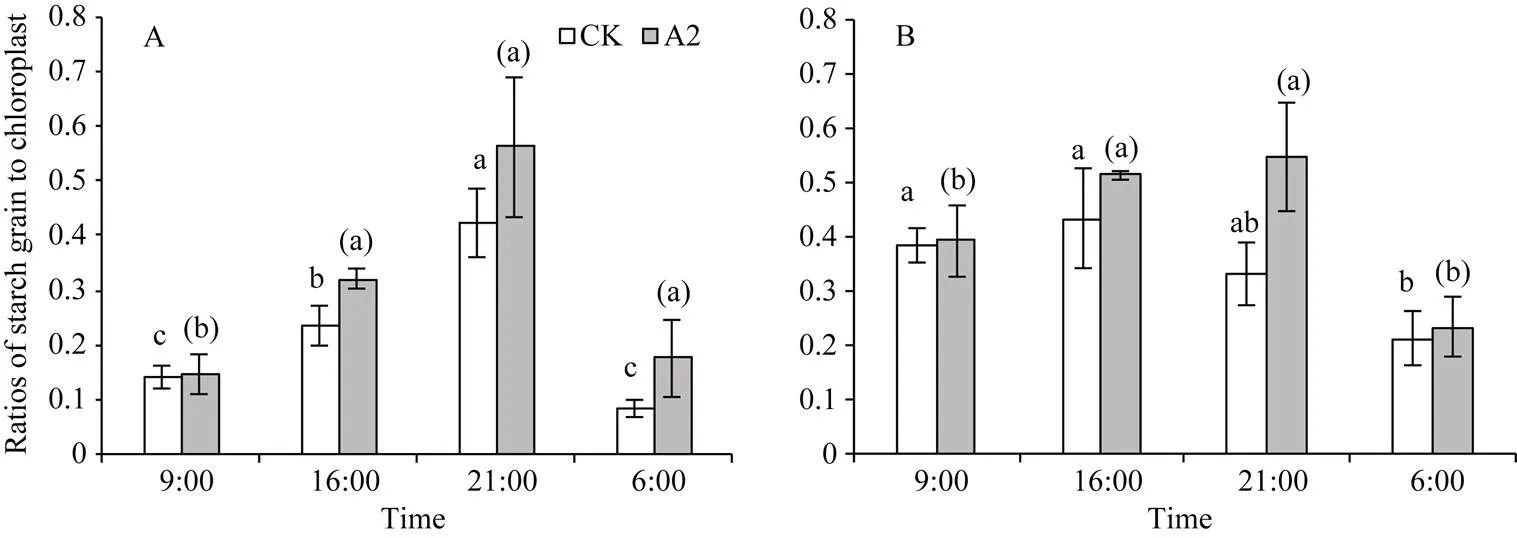
图4 不同光照强度下大豆叶片淀粉粒与叶绿体截面积比值
图A、图B分别为南豆12和桂夏3号在正常光照和A2处理下淀粉粒与叶绿体截面积比值的昼夜变化趋势。标以a、b、c的柱值表示CK处理下相同品种不同时间点的显著性差异, 标以(a)、(b)、(c)的柱值表示A2处理下相同品种不同时间点的显著性差异(0.05)。CK: 正常光照, 遮光度0; A2: 两层黑色遮阳网, 遮光度36%。
Figure A and B show the diurnal variation of ratios of starch to chloroplast cross-sectional area under CK and A2 treatment for Nandou 12 and Guixia 3. Bars superscripted by a, b, and c in the figure are significant by different at different time points for the same cultivar under the CK treatment, and those by (a), (b), (c) are significant by different at the different time points for the same cultivar under the A2 treatment (0.05). CK: normal light, shading 0; A2: two black shading nets, shading 36%.
叶绿体为高等植物特有的能量转换的细胞器, 其发育受到光的调控, 不同光照条件下的植物叶绿体数目会发生变化[19]。本研究中, 正常光照中南豆12淀粉粒与叶绿体截面积比值昼夜变化差异显著, 在21:00比值最大, A2处理下大豆淀粉粒与叶绿体截面积比值昼夜变化与CK相同, 且变化幅度较CK增大。说明光照强度的变化引起大豆光合器官结构的变化来适应环境变化, 且弱光环境下这种变化更显著。同时前人研究表明叶片的净光合速率与叶肉细胞内叶绿体数量极显著正相关[29], 这些变化有利于植物吸收较多的光能以弥补弱光的不足, 进而使光合作用的光化学效率增加[19]。
4 结论
在遮阴环境下, 植株通过调节光合器官结构增强对环境的适应性, 并降低净光合速率减少对同化物的消耗, 有利于光合同化物的积累。适度的荫蔽环境下, 不同品种大豆蔗糖和淀粉含量昼夜变化差异显著, 耐阴品种南豆12变化幅度高于敏感品种桂夏3号。因此, 适度的荫蔽环境有利于促进大豆光合同化物的积累和转化。
[1] 杨峰, 崔亮, 黄山, 刘卫国, 雍太文, 杨文钰. 不同株型玉米套作大豆生长环境动态及群体产量研究.大豆科学, 2015, 34: 402–407 Yang F, Cui L, Huang S, Liu W G, Yong T W, Yang W Y. Soybean growth environment and group yield in soybean relay intercropped with different leaf type maize., 2015, 34: 402–407 (in Chinese with English abstract)
[2] 杨峰, 娄莹, 刘沁林, 范元芳, 刘卫国, 雍太文, 王小春, 杨文钰. 玉米行距配置对套作大豆生物量、根系伤流及养分的影响.中国农业科学, 2016, 49: 4056–4064 Yang F, Lou Y, Liu Q L, Fan Y F, Liu W G, Yong T W, Wang X C, Yang W Y. Effect of maize row spacing on biomass, root bleeding sap and nutrient of soybean in relay strip intercropping systems., 2016, 49: 4056–4064 (in Chinese with English abstract)
[3] 王竹, 杨文钰, 伍晓燕, 吴其林.玉米株型和幅宽对套作大豆初花期形态建成及产量的影响. 应用生态学报, 2008, 19: 323–329 Wang Z, Yang W Y, Wu X Y, Wu Q L. Effects of maize plant type and planting width on the early morphologi calcharacters and yield of relay planted soybean., 2008, 19: 323–329 (in Chinese with English abstract)
[4] 刘鹏, 杨玉爱.钼、硼对大豆光合效率的影响. 植物营养与肥料学报, 2003, 9: 456–461 Liu P, Yang Y A. Effect of molybdenum and boron on photosynthetic efficiency of soybean., 2003, 9: 456–461 (in Chinese with English abstract)
[5] 吴其林, 王竹, 杨文钰. 苗期遮阴对大豆茎秆形态和物质积累的影响. 大豆科学, 2007, 26: 868–872 Wu Q L, Wang Z, Yang W Y. Seedling shading affects morphogenesis and substance accumulation of stem in soybean., 2007, 26: 868–872 (in Chinese with English abstract)
[6] 刘婷, 刘卫国, 任梦露, 杜勇利, 邓榆川, 邹俊林, 方萍, 杨文钰. 遮阴程度对不同耐阴性大豆品种光合及抗倒程度的影响. 中国农业科学, 2016, 49: 1466–1475 Liu T, Liu W G, Ren M L, Du Y L, Deng Y C, Zou J L, Fang P, Yang W Y. Effects of shade degrees on photosynthesis and lodging resistance degree of different shade tolerance soybean., 2016, 49: 1466–1475 (in Chinese with English abstract)
[7] 宋艳霞, 杨文钰, 李卓玺, 于晓波, 郭凯, 向达兵. 不同大豆品种幼苗叶片光合及叶绿素荧光特性对套作遮阴的响应.中国油料作物学报, 2009, 31: 474–479 Song Y X, Yang W Y, Li Z P, Yu X B, Guo K, Xiang D B. The effects of shading on photosynthetic and fluorescent characteristics of soybean seedlings under maize–soybean relay cropping., 2009, 31: 474–479 (in Chinese with English abstract)
[8] 苏本营, 宋艳霞, 陈圣宾, 杨文钰. 大豆幼苗对套作玉米遮阴环境的光合生理生态响应. 生态学报, 2015, 35: 3298–3308 Su B Y, Song Y X, Chen S B, Yang W Y. Photosynthetic responses of soybean () seedlings to shading caused by maize in an intercropping system., 2015, 35: 3298–3308 (in Chinese with English abstract)
[9] 范元芳, 杨峰, 刘沁林, 谌俊旭, 王锐, 罗式伶, 杨文钰. 套作阴蔽对苗期大豆叶片结构和光合荧光特性的影响. 作物学报, 2017, 43: 277–285 Fan Y F, Yang F, Liu Q L, Chen J X, Wang R, Luo S L, Yang W Y. Effects of shading on leaf structure and photosynthetic fluorescence characteristicsof soybean seedlings in maize– soybean relay tntercropping system., 2017, 43: 277–285 (in Chinese with English abstract)
[10] 李春红, 姚兴东, 鞠宝韬, 朱明月, 王海英, 张惠君, 敖雪, 于翠梅, 谢甫绨, 宋书宏. 不同基因型大豆耐阴性分析及其鉴定指标的筛选. 中国农业科学, 2014, 47: 2927–2939 Li C H, Yao X D, Ju B T, Zhu M Y, Wang H Y, Zhang H J, Ao X, Yu C M, Xie F T, Zhu S H. Analysis of shade–tolerance and determination of shade-tolerance evaluation indicators in different soybean genotypes., 2014, 47: 2927–2939 (in Chinese with English abstract)
[11] 武晓玲, 张丽君, 聂邵仙, 杨峰, 佘跃辉, 杨文钰. 弱光对大豆苗期生长及光合荧光特性的影响.大豆科学, 2014, 33: 53–57 Wu X L, Zhang L J, Nie S X, Yang F, She Y H, Yang W Y. Effects of low light on growth and photosynthetic fluorescence characteristics in soybean seedling., 2014, 33: 53–57 (in Chinese with English abstract)
[12] 于晓波, 梁建秋, 何泽民, 廖俊华, 张明荣, 吴海英, 明充, 唐琼英, 李小清. 玉米–大豆带状套作对大豆叶片形态及光合特性的影响.中国油料作物学报, 2016, 38: 452–459 Yu X B, Liang J Q, He Z M, Liao J H, Zhang M R, Wu H Y, Ming C, Tang Q Y, Li X Q. Response of leaf morphology and photosynthetic characteristics of soybean in maize–soybean relay strip intercropping system., 2016, 38: 452–459 (in Chinese with English abstract)
[13] 李艳大, 汤亮, 张玉屏, 刘蕾蕾, 曹卫星, 朱艳. 水稻冠层光合有效辐射的时空分布特征. 应用生态学报, 2010, 21: 952–958 Li Y D, Tang L, Zhang Y P, Liu L L, Cao W X, Zhu Y. Spatiotemporal distribution of photosynthetically active radiation in rice canopy., 2010, 21: 952–958 (in Chinese with English abstract)
[14] 吕丽华, 赵明, 赵久然, 陶洪斌, 王璞. 不同施氮量下夏玉米冠层结构及光合特性的变化. 中国农业科学, 2008, 41: 2624–2632 Lyu L H, Zhao M, Zhao J R, Tao H B, Wang P. Canopy structure and photosynthesis of summer maize under different nitrogen fertilizer application rates., 2008, 41: 2624–2632 (in Chinese with English abstract)
[15] Arnon D I. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in., 1949, 24: 1–15
[16] 舒展, 张晓素, 陈娟, 陈根云, 许大全. 叶绿素含量测定的简化. 植物生理学报, 2010, 46: 399–402 Shu Z, Zhang X S, Chen J, Chen G Y, Xu D Q. The simplification of chlorophyll content measurement.,2010, 46: 399–402 (in Chinese with English abstract)
[17] 张志良, 瞿伟菁. 植物生理学实验指导(第3版). 北京: 高等教育出版社, 2003. pp 127–130 Zhang Z L, Qu W J. Plant Physiology Experimental Guidance, 3rd edn. Beijing: Higher Education Press, 2003. pp 127–130 (in Chinese)
[18] 高俊凤. 植物生理学实验指导. 北京: 高等教育出版社, 2006. pp 144–148 Gao J F. Experimental Guidance for Plant Physiology. Beijing: Higher Education Press, 2006. pp 144–148 (in Chinese)
[19] 姚允聪, 王绍辉, 孔云. 弱光条件下桃叶片结构及光合特性与叶绿体超微结构变化.中国农业科学, 2007, 40: 855–863Yao Y C, Wang S H, Kong Y. Characteristics of photosynthesis machinism in different peach species under low light intensity., 2007, 40: 855–863 (in Chinese with English abstract)
[20] 范元芳, 杨峰, 王锐, 黄山, 雍太文, 刘卫国, 杨文钰. 弱光对大豆生长、光合特性及产量的影响. 中国油料作物学报, 2016, 38: 71–76 Fan Y F, Yang F, Wang R, Huang S, Yong T W, Liu W G, Yang W Y. Effects of low light on growth, photosynthetic characteristics and yield of soybean., 2016, 38: 71–76 (in Chinese with English abstract)
[21] 黄卫东, 吴兰坤, 战吉成. 中国矮樱桃叶片生长和光合作用对弱光环境的适应性调节.中国农业科学, 2004, 37: 1981–1985 Huang W D, Wu L K, Zhan J C. Growth and photosynthesis adaptation of dwarf-type Chinese Cherry (L. cv. Laiyang) leaves to weak light stress., 2004, 37: 1981–1985 (in Chinese with English abstract)
[22] 眭晓蕾, 张宝玺, 张振贤, 毛胜利, 王立浩. 不同品种辣椒幼苗光合特性及弱光耐受性的差异. 园艺学报, 2005, 32: 222–227 Sui X L, Zhang B X, Zhang Z X, Mao S L, Wang L H. Differences of photosynthetic character istics and low light- tolerance in seedlings of four pepper cultivars., 2005, 32: 222–227 (in Chinese with English abstract)
[23] 王一, 张霞, 杨文钰, 孙歆, 苏本营, 崔亮. 不同生育时期遮阴对大豆叶片光合和叶绿素荧光特性的影响. 中国农业科学, 2016, 49: 2072–2081 Wang Y, Zhang X, Yang W Y, Sun X, Su B Y, Cui L. Effect of shading on soybean leaf photosynthesis and chlorophyll fluorescence characteristics at different growth stages., 2016, 49: 2072–2081 (in Chinese with English abstract)
[24] 张瑞朋, 付连舜, 佟斌,吴晓秋, 朱海荣, 孙国伟. 大豆叶片光合作用与光强及二氧化碳的关系. 吉林农业科学, 2015, 40(3): 8–13 Zhang R P, Fu L S, Tong B, Wu X Q, Zhu H R, Sun G W. The relationship of soybean leaf photosynthesis with light intensity and carbon dioxide., 2015, 40(3): 8–13 (in Chinese with English abstract)
[25] Wang J, Feng Y. The effect of light intensity on biomass allocation, leaf morphology and relative growth rate of two invasive plants., 2004, 28: 781–786
[26] Yu S M, Lo S F, Ho T D. Source–sink communication: Regulated by hormone, nutrient, and stress cross-signaling., 2015, 20: 844–857
[27] 王书丽, 郭天财, 王晨阳, 查菲娜, 宋晓. 两种筋力型小麦叶、粒可溶性糖含量及与籽粒淀粉积累的关系. 河南农业科学, 2005, 34(4): 12–15 Wang S L, Guo T C, Wang C Y, Zha F N, Song X. Soluble sugar contents in leaf and grain in two gluten wheats and its relationship with grain starch accumulation., 2005, 34(4): 12–15 (in Chinese with English abstract)
[28] 罗兴录, 池敏青, 黄小凤, 谢和霞, 陆飞伍. 木薯叶片可溶性糖含量与块根淀粉积累的关系. 中国农学通报, 2006, 22(8): 289–291 Luo X L, Chi M Q, Huang X F, Xie H X, Lu F W. Studies on the relationship between soluble sugar content in the leaves and the starch accumulation in the root tuber of cassava,, 2006, 22(8): 289–291 (in Chinese with English abstract)
[29] 许燕. 6个特色茶树品种(系)光合特性及叶绿体超微结构研究. 四川农业大学硕士学位论文, 四川成都, 2016 Xu Y. Studies on Photosynthetic Traits and Chloroplast Ultrastmcture of Six Tea Characteristic Varieties (Lines). MS Thesis of Sichuan Agricultural University, Chengdu, Sichuan, China, 2016 (in Chinese)
Effects of Light Intensity on Photosynthetic Characteristics and Assimilates of Soybean Leaf
CHENG Ya-Jiao, FAN Yuan-Fang, CHEN Jun-Xu, WANG Zhong-Lin, TAN Ting-Ting, LI Jia-Feng, LI Sheng-Lan, YANG Feng*, and YANG Wen-Yu
College of Agronomy, Sichuan Agricultural University / Key Laboratory of Crop Ecophysiology and Farming System in Southwest, Ministry of Agriculture, Chengdu 611130, Sichuan, China
Light intensity plays a significant role in determining the growth and seed yield of crops under the sheltering of intercropping systems. By setting different light intensities, the photosynthetic characteristics, assimilate metabolism and the diurnal variation of chloroplast ultrastructure in leaf of different soybean cultivars to clarify the effect of light intensity on the structure of soybean leaf and the accumulation of carbohydrate, in order to provide a theoretical basis for improving soybean yield and quality. A pot experiment was carried out with three light intensities treatments, including CK (normal light, shading 0), A1 (black shading net, shading 10%) and A2 (two black shading nets, shading 36%) of two soybean cultivars (shade-resistant cultivar Nandou-12 and shade susceptible cultivar Guixia-3). With increased shading, net photosynthetic rate, stomatal conductance, transpiration rate and biomass decreased while intercellular carbon dioxide concentration and chlorophyllincreased in all treatments. There was a significant difference in the diurnal variation of sucrose and starch contents in soybean leaf under the same treatment. The diurnal variation of sucrose content in soybean leaf showed a bimodal curve with the peaks at 16:00 and next day 6:00 respectively under CK and A1 treatment. Under A2 treatment, the highest diurnal sucrose content was 32.80 μg g–1in Nandou 12 and showed a unimodal changing trend with the peak at 16:00. The diurnal variation of starch content showed a single-peak curve and the highest value appeared at 21:00. Compared with the shade sensitive cultivar, the shade-tolerant soybean had greater diurnal variation of sucrose content and starch content under A2 treatment. The chloroplast structure of soybean leaf was intact and unbroken under the low light treatment. The diurnal variation of the cross-sectional area ratio of starch grain to chloroplast in the same cultivar changed significantly under the same treatment, showing a trend of decreasing after increasing. The changing rate was greater in shade-tolerant cultivar than in sensitive cultivar, and the maximum value appeared at 21:00. Therefore, biomass accumulation of soybean leaves and photosynthesis decreased with increasing shading. However, shade-tolerant cultivar maintained a good photosynthesis by adjusting the diurnal variation of photosynthetic organ structure, thus better adapting to the moderate shading condition.
soybean; light intensity; photosynthetic characteristics; assimilate; chloroplast ultrastructure
2018-03-31;
2018-07-20;
2018-07-27.
10.3724/SP.J.1006.2018.01867
通信作者(Corresponding author): 杨峰, Email: f.yang@sicau.edu.cn
E-mail: 2644589298@qq.com
本研究由国家自然科学基金项目(31571615)和大学生创新性实验项目(201710626115)资助。
This study was supported by the National Natural Science Foundation of China (31571615) and the Undergraduate Innovative Experiment Project (201710626115).
URL:http://kns.cnki.net/kcms/detail/11.1809.S.20180726.1156.004.html
