内蒙古自治区2015-2018年B Yamagata系流感病毒全基因组序列分析
2018-11-02王慧馨张宇耕郭卫东张秀红
跃 华,海 岩,王慧馨,张宇耕,雷 霞,郭卫东,张秀红
国家科技重大专项(No.2017ZX10103006-003)和内蒙古自治区卫生计生科研计划项目(No.201702040)联合资助;(跃华和海岩对本文具有同等贡献,为并列第一作者)
流行性感冒(流感)是由流感病毒引起的急性呼吸道传染病,其主要通过近距离飞沫传播,具有高度传染性,其发病率居法定报告传染病的首位,是第一个实行全球性监测的传染病。由于流感病毒基因的高度变异性,人群对其普遍易感,易造成人群反复感染,发病率高,可在世界范围内广泛流行。每次流感流行后,都会造成不同程度的超额死亡和巨大的经济损失[1-2]。
流感病毒为单股负链RNA病毒,属正粘病毒科,其基因组由8 条分节段的RNA片段组成.根据其核蛋白(nucleoprotein,NP)和基质蛋白(matrixprotein,MP)的特性不同,可将流感病毒分为甲型(A)、乙型(B)、丙型(C)3型,最近有研究表明,在饲养动物(猪、牛) 中发现了丁型(D) 流感病毒[3-4]。与甲型流感相比,乙型流感不分亚型而根据其抗原性和基因特征的不同分为2个谱系,即B Victoria系和B Yamagata系[5]。2017年底至2018年初,中国流感呈现了一个高于同期水平的流行高峰,且以B Yamagata系流感病毒流行为主,与此同时,内蒙古自治区也出现相同的流行峰,为进一步了解内蒙古自治区B Yamagata系流感病毒的变异变迁情况,本文对2015-2018年内蒙古自治区的B Yamagata系流感病毒全基因特征进行分析,为乙型流感防控提供依据。
1 材料与方法
1.1毒株来源 本研究选取的2015-2018年B Yamagata系流感病毒株均来自于内蒙古自治区流感网络实验室。这些毒株是由各哨点医院上送的流感样病例咽拭子经犬肾传代细胞(MDCK Cell)培养和(或)鸡胚培养后所分离,再由血凝实验和血凝抑制实验复核确认后保存于-70 ℃冰箱。其中,2015年3株,2017年3株,2018年2株。
1.2病毒提取及基因扩增 病毒RNA提取采用QIAGEN公司的QIAamp Viral RNA Mini Kit,具体操作方法见试剂盒说明书;一步法PCR扩增试剂采用TaKaRa公司的PrimeScriptTM One Step RT-PCR Kit Ver.2,配置扩增反应体系见说明书,全基因组扩增引物参照文献[6],由上海伯杰生物科技有限公司合成。一步法RT-PCR反应程序如下:50 ℃ 30 min,94 ℃ 2 min;94 ℃ 30 s,60 ℃ 30 s,72 ℃ 1 min,30 cycles;72 ℃ 10 min,4℃保温。反应结束后取PCR产物5 μL进行1%琼脂糖凝胶电泳核实扩增目的条带,PCR产物测序由上海伯杰生物科技有限公司完成。
1.3全基因组序列拼接与分析 测序所得片段用DNAStar中的Seqman进行拼接,利用NCBI网站中的Blast比对验证所得的全基因序列,并用MEGA6.0软件使用Neighor-Joining(NJ)法绘制系统进化树,构建全基因系统发育树,并使用Bootstrap法进行检验,重复次数为1 000次。分析毒株基因遗传进化特征,并与WHO 推荐的疫苗株与和参比株的核苷酸进行比较分析毒株的遗传性、抗原性、耐药性等基因特征。分析中用到的疫苗株和参比株序列均来自GenBank和GISAID网站。
2 结 果
2.1全基因同源性分析 以WHO推荐北半球疫苗株B/Phuket/3073/2013为标准,对2015-2018年内蒙古自治区的8株B Yamagata系流感毒株全基因组序列进行核苷酸同源性分析。结果发现,所有毒株的8个基因节段与疫苗株的核苷酸同源性在97.7%~99.9%之间,其中,B/Inner Mongolia/1200/2015和B/Inner Mongolia/1178/2015毒株的NA基因核苷酸序列与疫苗株的同源性相对较低,分别为97.7%和97.8%,见表1。
表1 内蒙古自治区B Yamagata系流感毒株与疫苗株同源性分析
Tab.1 Homology analysis of sequences of all genes between B Yamagata viral isolates in Inner Mongolia and the vaccine strain

病毒分离株B/Phuket/3073/2013 (同源性 %)PB2PB1PAHANPNAMNSB/Inner Mongolia/113/201899.499.499.499.199.399.799.299.4B/Inner Mongolia/122/201899.399.599.698.899.399.799.599.2B/Inner Mongolia/1995/201799.499.599.698.899.399.799.498.9B/Inner Mongolia/1588/201799.599.599.098.399.699.799.299.1B/Inner Mongolia/1547/201799.499.399.699.199.699.799.099.1B/Inner Mongolia/1176/201599.799.799.999.699.899.999.799.5B/Inner Mongolia/1200/201599.699.499.199.598.297.799.699.4B/Inner Mongolia/1178/201599.899.399.899.598.297.899.799.5
2.2全基因遗传进化分析 通过绘制全基因进化树图发现,除NP基因外,其他节段在进化树中均形成明显的B Yamagata系和B Victoria系两大分枝。内蒙古自治区2015-2018年度的大部分B Yamagata系流感病毒株的各节段基因在进化树中共同处于同一独立分枝中,且与B/Phuket/3073/2013疫苗株距离较近;但B/Inner Mongolia/1178/2015和B/Inner Mongolia/1200/2015两个毒株的NP和NA基因与疫苗株处于不同分枝中,遗传距离较远,且出现HA-BY/NA-BV系间重配现象。HA、NA和NP基因进化情况见图1,其余节段基因进化树均与HA类似。
2.3 氨基酸序列分析
2.3.1血凝素基因(HA)氨基酸分析结果 内蒙古自治区2015-2018年B Yamagata系流感病毒株相对于B/Phuket/3073/2013疫苗株均出现L172Q和D196N变异,2017-2018年毒株均出现M251V突变,其中B/Inner Mongolia/1178/2015和B/Inner Mongolia/1200/2015两个毒株相对于疫苗株还出现T75N、K116R和D232N(近受体结合位点)变异,见表2。
2.3.2神经氨酸酶基因(NA)氨基酸分析结果 由于乙型流感病毒缺乏M2蛋白离子通道[7],故其对金刚烷胺类药物不敏感,故神经氨酸酶抑制剂为其首选治疗药物。NA基因的一些氨基酸突变会导致病毒对神经氨酸酶抑制剂敏感性降低,本次研究发现病毒株的神经氨酸酶活性中心仍保持较高的保守性,其功能部位的氨基酸序列为:E117、R150、D197、I221、H273、R292、N294、R374和G407(B Yamagata Number)[8],故病毒株仍对神经氨酸酶抑制剂敏感。
2.3.3HA糖基化位点分析 氨基酸的潜在糖基化位点是通过N-糖苷键结合在Asn(天冬氨酸)-X-Thr(苏氨酸)或Asn(天冬氨酸)-X-Ser(丝氨酸)的Asn残基上形成的。本次研究发现所有毒株HA蛋白中存在固定的11个潜在糖基化位点,分别位于:第25、59、145、167、196、303、332、491、517、530和577位,其中第196位位于抗原决定簇190螺旋中,而B/Inner Mongolia/1588/2017、B/Inner Mon-golia/1200/2015和 B/Inner Mongolia/1178/2015毒株在第232位出现D→N突变,增加了一个潜在糖基化位点。NA蛋白中存在固定的5个潜在糖基化位点,分别位于:第56、64、144、284和463位,而HA-BY/NA-BV重配株则缺少了第463位潜在糖基化位点。

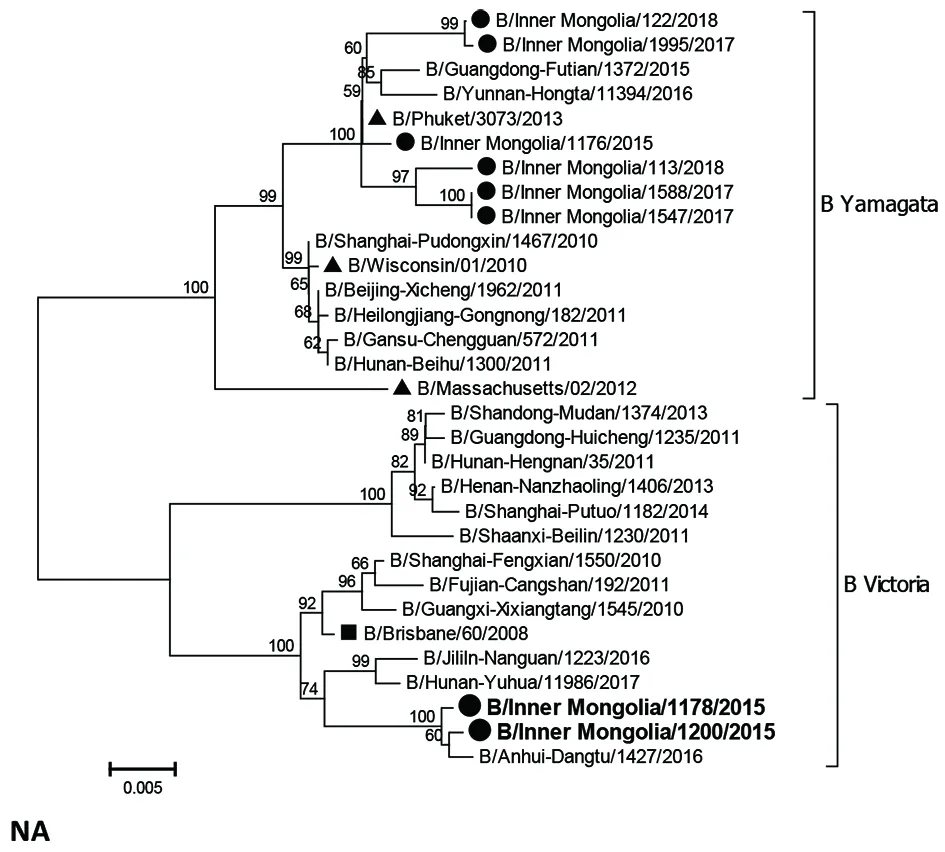

▲ B Yamagata系流感疫苗株, ■B Victoria系流感疫苗株, ●内蒙古B Yamagata系流感病毒株,黑体:系间重组流感病毒株图1 内蒙古自治区B Yamagata系流感病毒基因遗传进化分析Fig.1 Phylogenetic analysis of B Yamagata lineage viruses of Inner Mongolia
表2 内蒙古自治区B Yamagata系流感病毒血凝素基因氨基酸位点变异情况
Tab.2 Mutation sites of amino acid on the hemagglutinin gene of B Yamagata viruses in Inner Mongolia
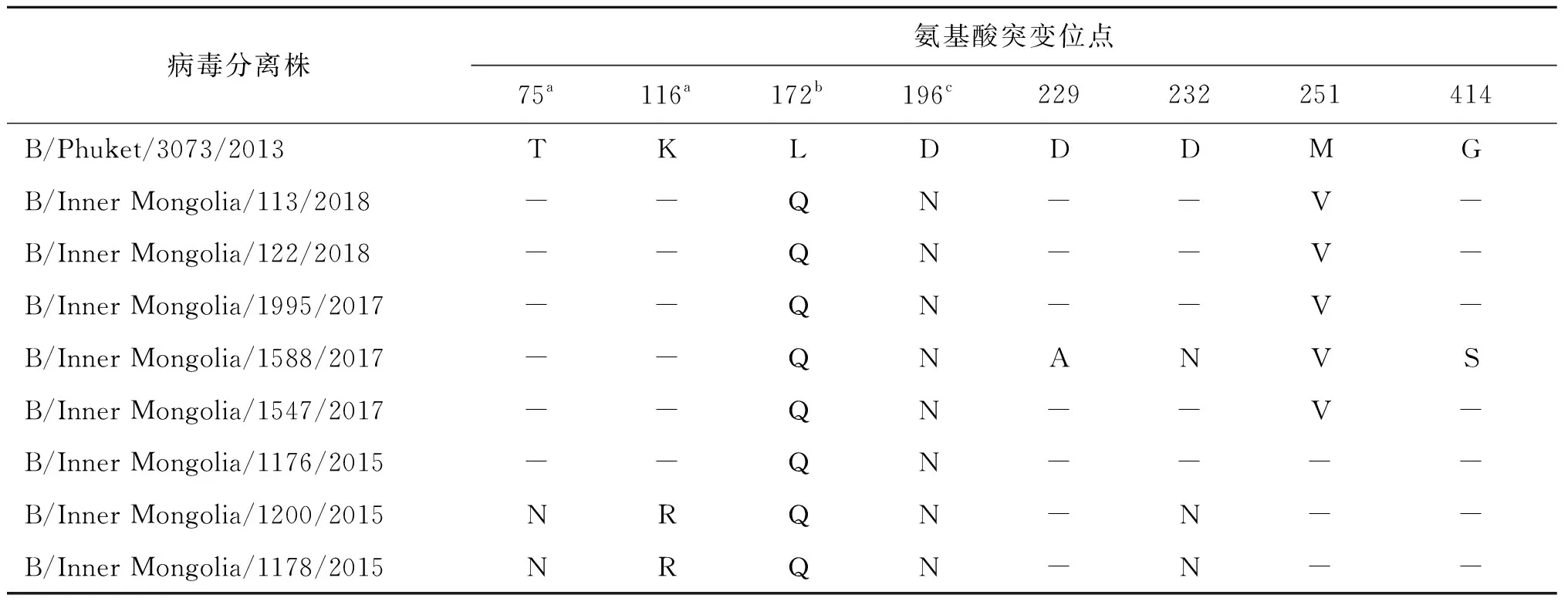
病毒分离株氨基酸突变位点75a116a172b196c229232251414B/Phuket/3073/2013TKLDDDMGB/Inner Mongolia/113/2018--QN--V-B/Inner Mongolia/122/2018--QN--V-B/Inner Mongolia/1995/2017--QN--V-B/Inner Mongolia/1588/2017--QNANVSB/Inner Mongolia/1547/2017--QN--V-B/Inner Mongolia/1176/2015--QN----B/Inner Mongolia/1200/2015NRQN-N--B/Inner Mongolia/1178/2015NRQN-N--
注: a b c分别表示HA 抗原表位120-loop、160-loop和190-helix处的突变位点
3 讨 论
自1983年以来,乙型流感根据其抗原性和基因特征分成了Yamagata和Victoria两大谱系。乙型流感虽然不会引起世界性大流行,但常常导致流感的局部暴发,其也是通过病毒表面蛋白的不断变异来改变病毒抗原性,逃避宿主免疫,从而导致不断的流行[9]。乙型流感的HA基因的抗原决定簇包含了4个区域,分别为:120环(116-137)、150环(141-150)、160环(162-167)和190螺旋(194-202),还包括120环的第75和77位氨基酸。上述氨基酸的改变会导致乙型流感病毒抗原发生漂移甚至发生转变。本次研究的病毒株在上述区域中的氨基酸序列相对保守,相对于B/Phuket/3073/2013疫苗株均出现L172Q和D196N变异。然而,B/Inner Mongolia/1178/2015和B/Inner Mongolia/1200/2015两个毒株除上述2个位点变异外,还出现T75N、K116R和D232N变异。流感病毒出现具有流行意义的新变异被定义为:在其抗原决定簇上有4个以上位点发生氨基酸替换,且替换涉及2个以上抗原决定簇[10],故上述2个毒株可以初步判断出现抗原漂移现象,可进一步结合抗原表型分析来确定是否为疫苗低反应株。流感病毒血凝素糖基化位点对蛋白的结构和功能十分重要,其主要作用是稳定血凝素蛋白结构,防止血凝素被水解以及阻碍被抗体识别等。本次糖基化分析中发现,8个毒株HA1区均有6个潜在糖基化位点,而B/Inner Mongolia/1178/2015和B/Inner Mongolia/1200/2015毒株还增加了第232位潜在糖基化位点,这些均可导致抗原表位被掩盖,可能会造成存在于宿主体内的原有抗体不能与抗原位点结合,成为流感流行的优势株。本次研究的病毒株NA基因的酶活性中心均保持了高度的保守性,未发现神经氨酸酶抑制剂耐药位点变异,故奥司他韦、扎那米韦等抗病毒药物仍是乙型流感的首选药物。
在同源性分析中发现,2015-2018年内蒙古自治区的B Yamagata系流感病毒与B/Phuket/3073/2013疫苗株保持高度同源,但B/Inner Mongolia/1200/2015和B/Inner Mongolia/1178/2015毒株的NA和NP基因核苷酸序列与疫苗株的同源性相对较低,结合遗传进化树图也可发现,这两株毒株的NA和NP基因距离疫苗株较远,其NA基因且出现了HA-BY/NA-BV的谱系间重配现象,进一步佐证疫苗可能对其不具有保护效果。上世纪80年代起,B型流感NP基因从Ⅲ型进化至如今的Ⅱ型,其中B Victoria和B Yamagata系流感病毒的NP基因在Ⅱ型进化树中交织进行[11-12],在进化过程中上述两株病毒的NP基因仍倾向于B Victoria系流感,如图所示,其进化距离已和B/Phuket/3073/2013疫苗株相距较远,推测NP基因可能发生了内部重排,或许与其他节段蛋白的协同作用有关,可进一步开展分子生物学深入研究。通过流感病原学监测发现[13],内蒙古自治区2013年以甲型H1N1流感为流行优势株,2014年以H3N2流感为流行优势株,而2015年初B Yamagata系流感成为人群主要流行优势株,这或许也与本次病毒基因研究发现2015年的毒株发生抗原漂移,HA和NA发生基因谱系间重配,NP发生基因内部重排,而导致人群免疫屏障无法建立有效保护有关。其余6份毒株的8个基因节段与疫苗保持了高度同源,同源性均达99.0%以上,并且在进化树中同处一分枝中,故可以判断疫苗对其具有较好的保护效果。
由于B Yamagata和B Victoria系流感病毒可在人群中同时流行,增加了二者发生基因重配的机会,通过全基因分析发现本次重配仅发生在HA和NA基因之间,未见其他基因的重配现象,重配是否影响乙型流感病毒的生长特性、毒力、致病性等,可进一步深入研究。
