云南玉龙鼠疫疫源地宿主动物群落结构特征及其流行病学意义
2018-11-02,,,,,,,,,,
,,,,,, , ,,,
玉龙鼠疫自然疫源地是2006年在云南新发现的一块鼠疫自然疫源地[1],既往调查研究不仅证实和确定了鼠疫疫源地及其范围,而且还对宿主动物和媒介的组成,以及鼠疫菌的生化特性和遗传特征进行了初步研究[1-5],但是对该疫源地宿主动物群落结构及其分布格局等生物学基础信息尚未系统认识,而鼠疫宿主动物的生物学特征是认知鼠疫自然疫源地结构和分布流行规律的一个关键问题[6]。因此,本研究在玉龙自然疫源地核心区域(以玉龙县鹿子村为中心),系统调查研究宿主动物群落结构特征和分布格局规律及其与动物鼠疫流行的关系,对探讨该疫源地的结构和性质具有重要意义。
1 资料与方法
1.1 资料
1.1.1小型兽类(简称小兽)资料 来源于根据地理环境和植被分布特征,将玉龙鼠疫疫源地划分为4个不同海拔梯度带(I. 2 392~2 650 m,II. 2 650~2 800 m,III. 2 800~3 100 m,IV. 3 100 m~)[7]和2种生境(林地和耕地),于2015年春夏秋冬4个季节,共对32个样点中小兽进行调查取样、分类鉴定和计数。
1.1.2鼠疫疫情资料 丽江市鼠疫疫情报告、疫源地调查资料和相关文献资料。
1.2 方法

1.2.2统计分析 用SPSS17.0软件对群落生态学指标进行统计分析,捕获率比较用χ2检验,不同生境丰富度、多样性、均匀度和优势度比较用用Mann-Whitney秩和检验,P<0.05为差异有统计学意义。
2 结 果
2.1小兽种类组成和数量分布 本次调查共布放回收有效鼠夹7940夹次,捕获小兽1 575只,隶属4目6科12属22种,平均捕获率为19.84%,齐氏姬鼠、大绒鼠和中华姬鼠为疫源地小兽群落的优势种,构成比分别为44.19%、19.62%和14.35%。耕地小兽捕获数量明显高于林地,耕地和林地捕获率分别为23.75%和15.89%(χ2=77.227,P<0.001),构成比分别为60.13%和39.87%,林地优势种依次为中华姬鼠(30.73%)、齐氏姬鼠(19.90%)、大绒鼠(17.04%)、大耳姬鼠(14.65%);而耕地优势种依次为齐氏姬鼠(60.30%)和大绒鼠(21.33%)。见表1,表2。
2.2小兽群落结构的垂直分布 在垂直分布上,小兽捕获数量总体随疫源地垂直梯度增高呈上升的趋势,构成比和捕获率以IV垂直带(3 100m~)最高,分别为42.35%和33.28%,明显超过前3个垂直带,4个垂直带之间捕获率差异有统计学意义(χ2=319.390,P<0.001),其中I垂直带(2 392 m~2 650 m)林地中捕获小兽最少,构成比和捕获率分别为13.22%和 8.57%,IV垂直带(3 100 m~)耕地中捕获小兽最多,构成比和捕获率分别为42.66%和 40.12%;疫源地内各种小兽分布对垂直梯度和植被环境具有一定的选择性,其中齐氏姬鼠、大绒鼠、中华姬鼠和大耳姬鼠的垂直分布范围较宽,在4个垂直带中数量分布均较多,显示出对不同垂直梯度及植被环境具有较强的适应和占领能力,其余种类小兽分布范围相对局限一些,而有的小兽垂直分布范围很窄,仅局限于1个垂直带,如玉龙绒鼠仅见于IV垂直带(3 100 m~)。见表1。
表1 玉龙鼠疫疫源地小兽群落结构的垂直分布
Tab.1 The vertical distribution of community structure of small mammals in Yulong natural plague foci

种 类林地耕地合计构成比(%)IIIIIIIV合计构成比(%)IIIIIIIV合计构成比(%)安氏白腹鼠20.320 1 1 090.950 3 6 0 110.7社鼠335.2512 17 4 060.630 4 2 0 392.48黄胸鼠000 0 0 070.740 5 1 1 70.44大足鼠10.160 1 0 040.422 1 0 1 50.32褐家鼠000 0 0 020.210 2 0 0 20.13巢鼠10.160 0 1 0000 0 0 0 10.06大耳姬鼠9214.654 7 3 78272.850 8 1 18 1197.56齐氏姬鼠12519.913 35 26 5157160.385 98 84 304 69644.19中华姬鼠19330.7328 52 81 32333.480 17 14 2 22614.35大绒鼠10717.041 9 6 9120221.3353 45 52 52 30919.62黑腹绒鼠20.320 1 0 190.953 2 3 1 110.7玉龙绒鼠50.80005151.5800015201.27侧纹岩松鼠10.16100000000010.06珀氏长吻松鼠132.07606130.320030161.02臭鼩鼱10.16100000000010.06灰麝鼩40.640310181.92817221.4中麝鼩10.16010020.21000230.19白尾梢麝鼩60.961500252.6414920311.97南小麝鼩00000010.11000110.06印度长尾鼩81.27004400000080.51树鼩335.2516890121.272550452.86黄腹鼬00000010.11001010.06捕获数量(只)628-83140142263947-1612071754041575构成比(%)-10013.2122.2922.6141.8810017.0021.8618.4842.66-100有效工具数3953-9689989909973987-978102397910077940-捕获率(%)15.89-8.5714.0314.3426.3823.75-16.4620.2317.8840.1219.84-
注:1.垂直梯度(I:2392m~2650m,II:2650m~2800m,III:2800m~3100m,IV:3100m~)
2.安氏白腹鼠(Niviventerandersoni);社鼠(Niviventerconfucianus);黄胸鼠(Rattustanezumi);大足鼠(Rattusnitidus);褐家鼠(Rattusnorvegicus);巢鼠(Micromysminuts);大耳姬鼠(Apodemuslatronum);齐氏姬鼠(Apodemuschevrieri);中华姬鼠(Apodemusdraco);大绒鼠(Eothenomysmiletus);黑腹绒鼠(Eothenomysmelanogaster);玉龙绒鼠(Eothenomysproditor);侧纹岩松鼠(Sciurotamiasforresti);珀氏长吻松鼠(Dremomyspernyi);臭鼩鼱(Suncusmurinus);灰麝鼩(Crociduraattenuate);中麝鼩(Crocidurarussula); 白尾梢麝鼩(CrociduraDracula); 南小麝鼩(Crocidurahorsfieldi); 印度长尾鼩(Soriculusleucops); 树鼩(Tupaiabelangeri); 黄腹鼬(Mustelasibirica)
2.3小兽群落结构的季节变化 在季节分布上,小兽捕获数量随春-夏-秋-冬季节变化呈下降趋势,构成比和捕获率以春季最高,分别为30.03%和23.45%,夏秋两季次之,冬季最低,分别为17.46%和13.76%,4个季节之间捕获率差异有统计学意义(χ2=67.517,P<0.001),其中冬季林地中捕获小兽最少,构成比和捕获率分别为18.31%和11.45%,春季耕地中捕获小兽最多,构成比和捕获率分别为30.41%和27.85%。见表2。
表2 玉龙鼠疫疫源地小兽群落结构的季节变化
Tab.2 The seasonal variation of community structure of small mammals in Yulong natural plague foci

项目指标林地耕地合计春夏秋冬小计春夏秋冬合计捕获数(只)6281851751531159472882512481601575构成比(%)10029.4627.8724.3618.3110030.4126.526.1916.9100有效工具数39539839789881004398710349899709947940捕获率(%)15.8918.8217.8915.4911.4523.7527.8525.3825.5716.119.84
2.4小兽群落多样性分布格局 两种生境中小兽群落物种丰富度相似,林地物种多样性和均匀度较高,生态优势度较低,而耕地则与之相反。林地和耕地多样性指数分别为1.915 3和1.383 9,均匀度指数分别为0.662 7和0.478 8,优势度分别为0.190 9和0.412 9。见表3。
不同垂直梯度小兽群落物种丰富度近似,小兽物种多样性和均匀度随着垂直梯度的升高呈现出下降趋势,而优势度则呈升高趋势,多样性指数在1.354 0-1.875 7之间,均匀度指数在0.527 9-0.710 7之间,优势度在0.236 3-0.353 6之间;其中多样性指数和均匀度指数以II垂直带最高,IV垂直带最低,而生态优势度以IV垂直带最高,I垂直带最低。见表4。
不同季节小兽群落物种丰富度差别不大,小兽物种多样性和均匀度随春-夏-秋-冬季节变化呈上升趋势,而优势度则呈逐渐下降趋势,多样性指数在1.518 5-1.904 2之间,均匀度指数在0.575 4-0.686 8之间,优势度在0.221 5-0.330 0之间;其中多样性指数和均匀度指数秋冬两季高于春夏两季,冬季最高,夏季最低,而生态优势度则反之。见表5。
对两种生境小兽生态学指标进行秩和检验,不同垂直带两种生境之间除均匀度差别具有统计学意义外,其余丰富度、多样性和优势度差别均无统计学意义;不同季节两种生境之间除丰富度差别无统计学意义外,其余多样性、均匀度和优势度差别均具有统计学意义。见表6。
表3 玉龙鼠疫疫源地不同生境小兽群落多样性分布
Tab.3 The community diversity of small mammals in different habitats in Yulong natural plague foci
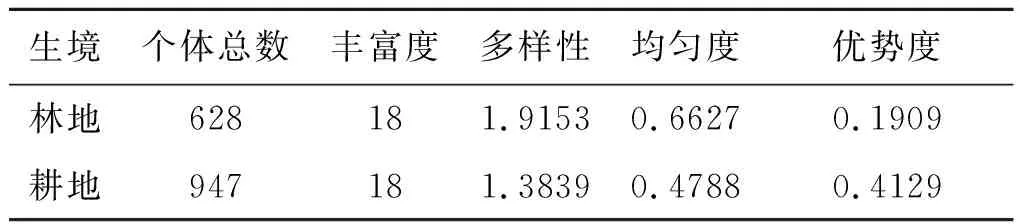
生境个体总数丰富度多样性均匀度优势度林地628181.91530.66270.1909耕地947181.38390.47880.4129
表4 玉龙鼠疫疫源地不同海拔小兽群落多样性分布
Tab.4 The community diversity of small mammals in different altitudinal gradients in Yulong natural plague foci

海拔丰富度多样性均匀度优势度I131.79690.70060.2363II141.87570.71070.2207III151.71020.63150.2478IV131.35400.52790.3536
表5 玉龙鼠疫疫源地不同季节小兽群落多样性分布
Tab.5 The community diversity of small mammals in different seasons in Yulong natural plague foci
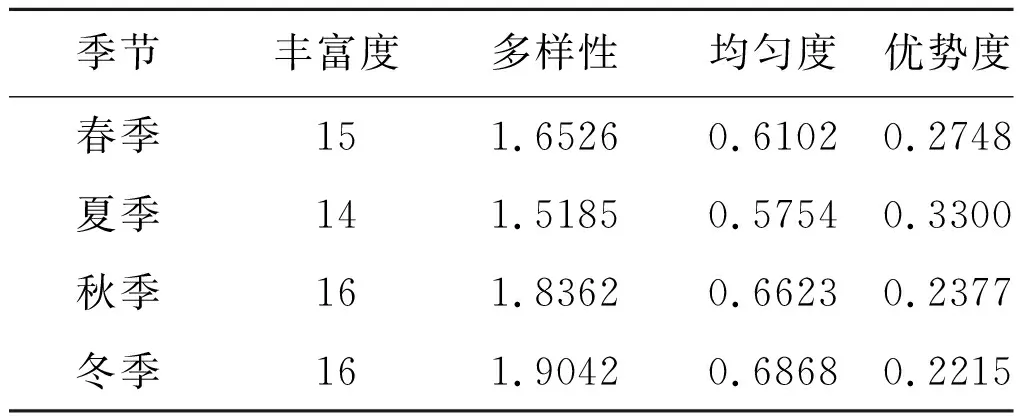
季节丰富度多样性均匀度优势度春季151.65260.61020.2748夏季141.51850.57540.3300秋季161.83620.66230.2377冬季161.90420.68680.2215
表6 玉龙鼠疫疫源地两种生境不同海拔、不同季节小兽群落生态学指标统计检验
Tab.6 The statistical test of community ecological indicators of small mammals in Yulong natural plague foci

生境项目丰富度多样性均匀度优势度林地-耕地海拔Z=-0.730 P=0.465Z=-1.155 P=0.248Z=-2.021 P=0.043Z=-1.732 P=0.083季节Z=-0.749 P=0.454Z=-2.309 P=0.021Z=-2.309 P=0.021Z=-2.309 P=0.021
2.5玉龙鼠疫自然疫源地鼠疫流行情况 2005年以来,先后3次对玉龙鼠疫疫源地开展了调查,依据指示动物血清学调查结果证实[1,3,10],目前玉龙鼠疫疫源地范围分布于海拔在2 230~3 300 m之间,整体环境属于亚高山温性针阔混交林景观类型,位置紧邻剑川野鼠疫源地,涉及2个县(区),8个乡(镇),26个村委会,59个村民小组,面积大约650平方公里,其中核心区域位于海拔2 500~3 000 m的中山林地及山间耕作地。连续多年调查、监测证实[10-13],截至2015年底,疫源地鼠疫流行7个年次,其中发生肺鼠疫疫情1起,鼠疫病人5例,死亡2例,动物间鼠疫6起,分离到鼠疫菌 9 株(分别为齐氏姬鼠3株,大绒鼠2株,大足鼠1株,玉龙绒鼠1株,特新蚤指名亚种2株),检测出鼠疫抗原阳性11份,累计证实疫点11个。从垂直梯度分布来看,除1个疫点位于II垂直带(2 650 m~2 800 m),其余疫点均分布于IV垂直带(3 100 m~);从流行时间来看,有2个年度的疫情发生于11月,其余5次均流行于4-5月。
3 讨 论
齐氏姬鼠和大绒鼠不仅在云南玉龙鼠疫疫源地各海拔梯度中呈连续性分布,也是各海拔梯度带和不同生境中的优势种群,均从两种鼠中分离到鼠疫菌,它们作为相邻鼠疫疫源地(剑川疫源地)的主要宿主地位已得到证实[14],因此,可认为齐氏姬鼠和大绒鼠符合作为该疫源地主要宿主动物的基本条件,对动物鼠疫的保存和流行具有重要的作用。虽然有学者认为玉龙绒鼠是该疫源地的主要宿主[6,15],但调查发现玉龙绒鼠仅集中分布于IV梯度带,显示出该种群对环境梯度选择适应的特异性,虽有局部增高,但其种群优势地位还未确立,尚不能考虑其作为该疫源地的主要宿主。
生境是疫源地地理景观具体环境的呈现,生境条件的不同则物种多样性不同,其复杂程度决定物种多样性变化,耕地中小兽密度高于林地,林地小兽的物种多样性指数、均匀度都大于耕地,优势度小于耕地,这与物种多样性随生境环境复杂性变化相一致,越复杂的生境其多样性和均匀度就大,单一化生境小兽生态优势度相对较高,具体而言,主要宿主动物齐氏姬鼠和大绒鼠在耕地生境中的优势地位突出,说明该疫源地云南松林山间耕作区是鼠疫发生和流行最适生境。
不同海拔梯度中,小兽数量分布随垂直梯度的升高而增多,以IV垂直带小兽捕获数量和捕获率最高,这种现象不是偶然的,IV垂直带土壤为棕红壤,土质为沙土,较松软,农作物主要为土豆、蔓菁、玛卡、豆子、油菜、小麦等根茎植物,食源丰富,且易获得,洞穴遍布田间地埂或树根,有利于小兽的繁殖和生存,使宿主动物容量始终维持在一个较高的水平,这有利于鼠疫的流行和暴发。另外,小兽物种多样性和均匀度随着海拔梯度的升高呈下降趋势,而优势度呈上升趋势,其中以IV梯度的物种多样性最低而优势度最高,反映出宿主动物的空间分布格局表现为一定程度的聚集性分布,从另外一个角度表明IV梯度带是该疫源地动物鼠疫流行的高发区。
不同季节分布中,小兽数量分布随春-夏-秋-冬季节变化呈下降趋势,构成比和捕获率以春季最高,小兽物种多样性、丰富度和均匀度随季节的变化呈上升趋势,生态优势度则与之相反呈递减趋势,小兽群落物种多样性、均匀度均以夏季为最低,而生态优势度则反之。梁健[16]等在对人工林小兽群落季节动态的研究中发现优势种时空变化决定了群落的主要结构特征。因此,一方面提示该疫源地春季气温变暖,小兽觅食、交配等活动频繁,宿主动物达到繁殖高峰,尤其是优势种数量达到峰值,另一方面提示宿主群落物种多样性季节变化与鼠疫暴发流行的时间性关系密切,是该疫源地动物鼠疫反复流行于春末夏初的重要原因。
综上所述,影响鼠疫分布、发生和流行的因素很多,本文仅通过分析宿主动物群落结构及其物种多样性的时空分布格局,结合动物鼠疫流行分布特征,初步阐述了宿主动物群落结构特征的流行病学意义,为及时研判和预警鼠疫疫情,科学制订防治策略提供科学依据。
