短期饥饿处理对卵形鲳鲹鱼肉品质的影响
2018-10-29钟金香李俊伟颉晓勇郭永坚马海霞朱长波
钟金香, 李俊伟, 颉晓勇, 郭永坚, 马海霞, 朱长波
(1. 广东省海洋与渔业技术推广总站, 广东 广州 510222;2.中国水产科学研究院南海水产研究所∥广东省渔业生态环境重点实验室∥农业部南海渔业资源开发利用重点实验室, 广东 广州 510300)
卵形鲳鲹(Trachinotusovatus),俗名金鲳、黄腊鲳,地方名称卵鲹、红三、红沙、仓鱼等,属于暖水性鱼类,其适应性强、生长速度快、营养价值高,是中国南方沿海主要经济鱼类品种之一,昔日名贵食用鱼类.近年因养殖密度不断增大、养殖海区环境恶化等因素,导致养殖病害频发,相应造成鱼肉品质下降,甚至引发食品安全担忧[1-3],制约了卵形鲳鲹产业的健康发展.近十几年大规模养殖发展,中国基本解决了鱼类产量问题,人们生活水平从温饱过渡到小康,消费购买力增强的同时愈加关注食品安全和质量差异,卵形鲳鲹养殖产业的最大问题已逐渐转化为产品质量没有跟上消费升级要求,无法满足市场对优质高档水产品的需求.
养殖环境因素对于鱼类品质和营养成分具有显著影响[4-7].国外控制投饲作为改良肉类品质的重要手段而得到广泛应用[8-9],国内近年多地出现企业将养成的商品规格鱼转移至循环水养殖车间,进行品质改良10~30 d再出售,并以其优异品质创建“瘦身鱼”品牌,提升产品附加值.国内外有关卵形鲳鲹养殖技术[10-12]和病害防治[13-14]的报道较多,但是鱼肉品质改良相关研究较为缺乏,饥饿相关研究主要集中于仔稚幼鱼补偿生长[15-19],到目前为止采用饥饿方式对商品规格卵形鲳鲹进行品质改良的内在机理尚不清楚.因此,本研究探讨饥饿过程中鱼体形态学、肌肉生化组成、品质构成、氨基酸、脂肪酸等变化,为池塘养殖卵形鲳鲹品质改良生产提供科学依据.
1 材料与方法
1.1 实验设计
实验鱼取自广东省阳江市沙扒镇海水养殖池塘,鱼体质量均值(535.04±36.33)g.实验前先用金鲳鱼商用配合饲料在车间水泥池中驯养2周,养殖水体溶解氧质量浓度5 mg/L以上,水温(22.5±1.0)℃,待摄食正常后开始饥饿实验.在室内循环水养殖水泥池中放置规格为1 m×1 m×1 m的60目尼龙网做成的网箱,实验设置0(S0)、5(S5)、10(S10)和15 d(S15)共4组,每组实验鱼30尾,设3个重复,为期16 d.饥饿初始,从每个重复中分别随机取鱼2尾,记为S0(饥饿0 d),之后每5天从3个重复中各随机取样2尾,分别记为S5(饥饿5 d)、S10(饥饿10 d)、S15(饥饿15 d).
1.2 样品采集和实验仪器
将所取鱼样在量鱼板上用卷尺测量体长(精确至0.1 cm).解剖,用电子秤称量鱼体质量、内脏质量和肝脏质量(精确至0.01 g).从每尾鱼样取背肌(鱼体左侧,侧线以上背鳍以下位置,10 mm×10 mm×10 mm方块)立即用于品质构成分析.右侧对应部位肌肉样品及肝脏样品于-20 ℃冰箱冷冻保存待测.
实验仪器:凯氏定氮仪,日本岛津公司UDK132型;氨基酸分析仪,日本日立公司L-8900型;气相色谱仪,美国Agilent公司6890型;质构仪,英国CNS Farnell 公司QTS-25型.
1.3 测定指标与方法
肥满度(condition factor,CF,g/cm3)=
100×m/l3;
脏体比(viscerosomatic index,VSI,%)=
100×mV/m;
肝体比(hepatosomatic index,HSI,%)=
100×mH/m.
式中m为鱼体质量(g);l为鱼体长(cm);mV为内脏质量(g);mH为肝脏质量(g).
营养成分测定:水分用常压恒温105 ℃干燥法(GB 5009.3-2016);粗蛋白用凯氏定氮法(GB 5009.5-2016);灰分用550℃高温灼烧法(GB 5009.4-2016);粗脂肪用索氏抽提法(GB 5009.6-2016);氨基酸组成采用酸水解法(GB 5009.124-2016);脂肪酸组成根据国际通用方法(AOAC 996.06-2001)采用气相色谱仪测定.
品质构成试验参数设置参考方静等[20]的设置,采用质构仪测定,记录硬度、弹性、凝聚性等品质构成参数.
1.4 统计处理
2 结果
2.1 短期饥饿对卵形鲳鲹形态学指标的影响
在15 d饥饿处理过程中卵形鲳鲹形态学参数发生明显变化(表1).肥满度、脏体比、肝体比均呈稳定下降趋势,其中脏体比S5组与S0组相比呈显著性差异(P<0.05),肥满度和肝体比S10组与S0组相比有统计学差异(P<0.05).
表1饥饿胁迫对卵形鲳鲹形态学指标的影响
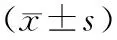
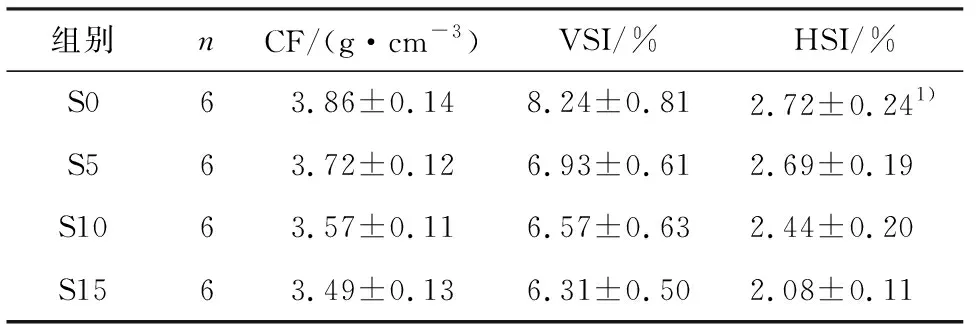
组别nCF/(g·cm-3)VSI/%HSI/%S063.86±0.148.24±0.812.72±0.241)S563.72±0.126.93±0.612.69±0.19S1063.57±0.116.57±0.632.44±0.20S1563.49±0.136.31±0.502.08±0.11
1)P<0.05,有统计学差异.
2.2 饥饿对卵形鲳鲹肌肉和肝脏组织生化组成的影响
在饥饿处理过程中,卵形鲳鲹肌肉组织水分质量分数稳定上升,组间有统计学差异(P<0.05);粗蛋白质量分数略有下降但组间无统计学差异(P>0.05);粗脂肪呈显著下降趋势(P<0.05);灰分质量分数随饥饿时间延长而升高,组间有统计学差异(P<0.05).肝脏组织中粗蛋白和粗脂肪质量分数同时呈显著下降趋势(P<0.05,表2).
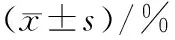
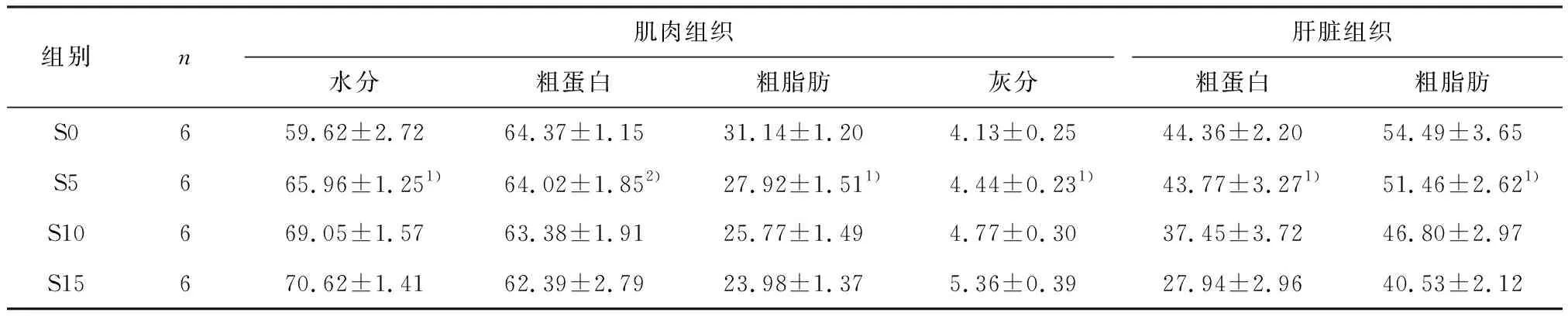
组别n肌肉组织肝脏组织水分粗蛋白粗脂肪灰分粗蛋白粗脂肪S0659.62±2.7264.37±1.1531.14±1.204.13±0.2544.36±2.2054.49±3.65S5665.96±1.251)64.02±1.852)27.92±1.511)4.44±0.231)43.77±3.271)51.46±2.621)S10669.05±1.5763.38±1.9125.77±1.494.77±0.3037.45±3.7246.80±2.97S15670.62±1.4162.39±2.7923.98±1.375.36±0.3927.94±2.9640.53±2.12
1)P<0.05,组间有统计学差异;2)P>0.05,组间无统计学差异.
2.3 饥饿对卵形鲳鲹肌肉组织品质构成参数的影响
随饥饿处理时间延长,卵形鲳鲹背部肌肉组织的凝聚性和弹性指数呈升高趋势,组间有统计学差异(P<0.05).硬度指数随饥饿时间延长呈下降趋势,组间有统计学差异(P<0.05).品质构成参数反映肉质软硬程度和弹性等,凝聚性越高,咀嚼时越细腻,口感也越好[21].本实验结果表明饥饿处理对品质构成参数影响均有统计学差异(表3).
表3短期饥饿对卵形鲳鲹肌肉品质构成指标的影响
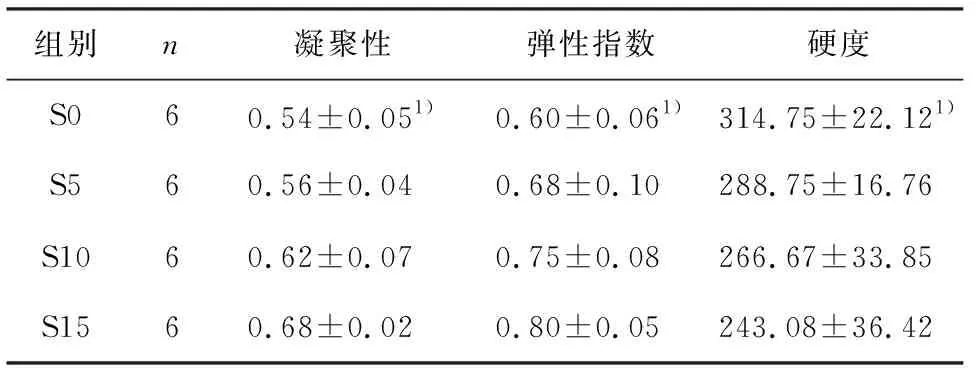
组别n凝聚性弹性指数硬度S060.54±0.051)0.60±0.061)314.75±22.121)S560.56±0.040.68±0.10288.75±16.76S1060.62±0.070.75±0.08266.67±33.85S1560.68±0.020.80±0.05243.08±36.42
1)P<0.05,组间有统计学差异.
2.4 饥饿对卵形鲳鲹肌肉组织氨基酸组成和质量分数的影响
经15 d饥饿处理之后,组氨酸质量分数降低,脯氨酸和精氨酸质量分数升高,且组间存在统计学差异(P<0.05).缬氨酸、蛋氨酸、异亮氨酸、苯丙氨酸等种类质量分数有所降低,苏氨酸、天冬氨酸、甘氨酸等种类有所升高,但无统计学差异(P>0.05,表4).
表4饥饿对卵形鲳鲹肌肉组织中氨基酸质量分数的影响

参数 nS0S15必需氨基酸(EAA) 苏氨酸Thr30.770±0.0261)0.783±0.059 缬氨酸Val30.780±0.0600.730±0.0531) 蛋氨酸Met30.560±0.0360.523±0.0421) 异亮氨酸Iso30.737±0.0550.673±0.0491) 亮氨酸Leu31.457±0.1351.370±0.0791) 苯丙氨酸3Hep30.770±0.0600.747±0.0551) 赖氨酸Lys31.767±0.1551.650±0.1041) 组氨酸His30.513±0.0152)0.443±0.012 精氨酸Arg31.117±0.0061.260±0.0102) 总必需氨基酸∑EAA8.470±0.1378.150±0.2421)非必需氨基酸(NEAA) 天冬氨酸3)Asp31.590±0.0561)1.647±0.125 丝氨酸Ser30.630±0.0261)0.633±0.049 谷氨酸3)Giu32.637±0.1392.590±0.1441) 甘氨酸3)Giy30.977±0.0931)1.013±0.123 丙氨酸3)Ala31.147±0.0571.137±0.0641) 酪氨酸3)Tyr30.663±0.0550.617±0.0381) 脯氨酸Pro30.227±0.0310.287±0.0122) 总非必需氨基酸∑NEAA7.963±0.1857.830±0.3181) ∑EAA/∑NEAA1.064±0.0191.041±0.0361)
1)S0与S15组比较,P>0.05,无统计学差异;2)S0与S15组比较,P<0.05,有统计学差异;3)表示呈味氨基酸.
2.5 饥饿处理对卵形鲳鲹肌肉组织脂肪酸组成和质量分数的影响
饥饿对卵形鲳鲹肌肉组织中脂肪酸组成和质量分数的影响(表5).共检测到饱和脂肪酸(SFA)11种,单不饱和脂肪酸(MUFA)7种,多不饱和脂肪酸(PUFA)10种.饥饿处理后S15组SFA、 MUFA和PUFA均显著低于S0组(P<0.05),PUFA中C18 ∶2、C20 ∶2、C22 ∶2、C18 ∶3等种类质量分数组间有统计学差异(P<0.05),C20 ∶3(n-3)、C20 ∶4、EPA、DHA等种类饥饿处理后质量分数略有降低,但组间无统计学差异(P>0.05).
表5短期饥饿胁迫对卵形鲳鲹肌肉脂肪酸组成和质量分数的影响
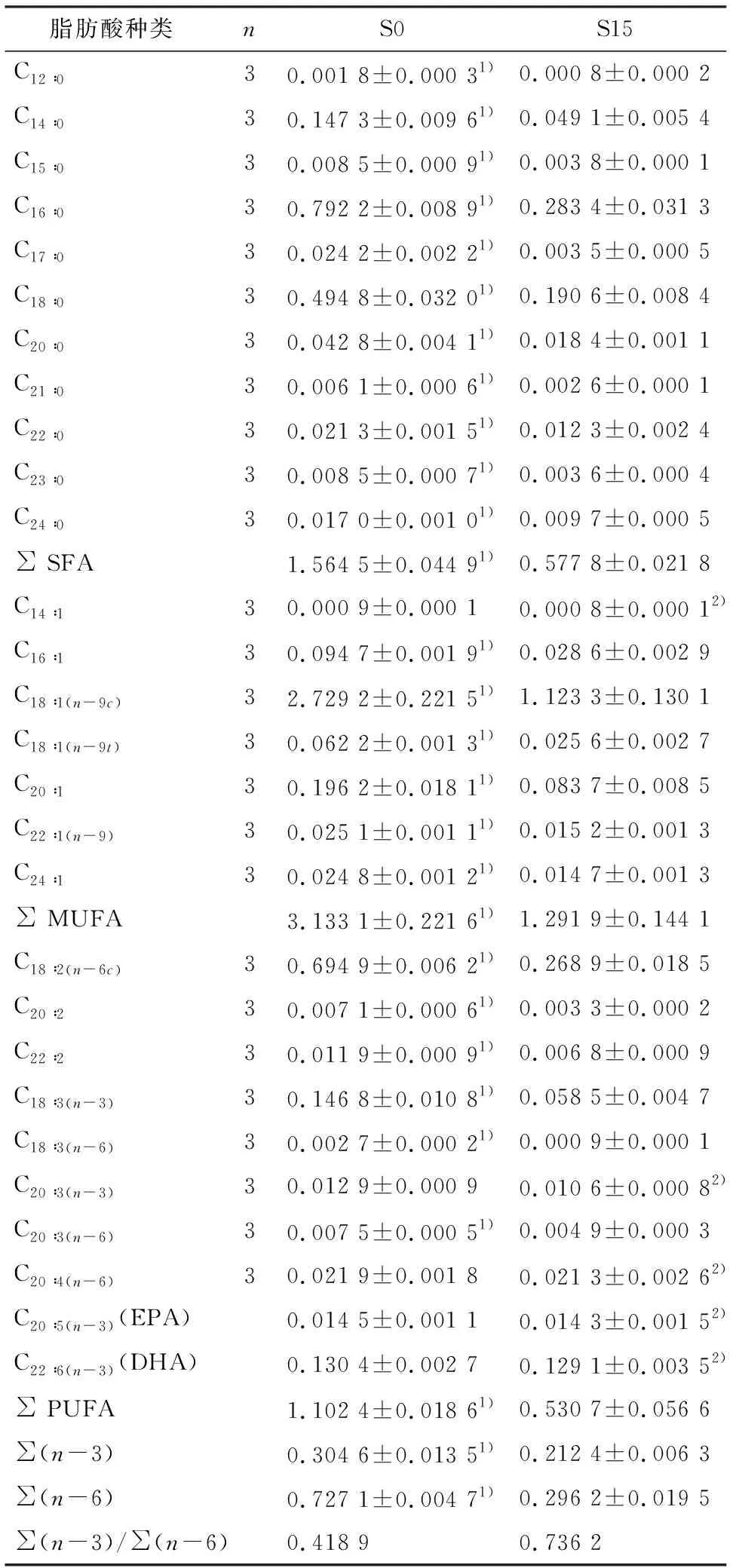
脂肪酸种类nS0S15C12∶030.001 8±0.000 31)0.000 8±0.000 2C14∶030.147 3±0.009 61)0.049 1±0.005 4C15∶030.008 5±0.000 91)0.003 8±0.000 1C16∶030.792 2±0.008 91)0.283 4±0.031 3C17∶030.024 2±0.002 21)0.003 5±0.000 5C18∶030.494 8±0.032 01)0.190 6±0.008 4C20∶030.042 8±0.004 11)0.018 4±0.001 1C21∶030.006 1±0.000 61)0.002 6±0.000 1C22∶030.021 3±0.001 51)0.012 3±0.002 4C23∶030.008 5±0.000 71)0.003 6±0.000 4C24∶030.017 0±0.001 01)0.009 7±0.000 5∑ SFA1.564 5±0.044 91)0.577 8±0.021 8C14∶130.000 9±0.000 10.000 8±0.000 12)C16∶130.094 7±0.001 91)0.028 6±0.002 9C18∶1(n-9c)32.729 2±0.221 51)1.123 3±0.130 1C18∶1(n-9t)30.062 2±0.001 31)0.025 6±0.002 7C20∶130.196 2±0.018 11)0.083 7±0.008 5C22∶1(n-9)30.025 1±0.001 11)0.015 2±0.001 3C24∶130.024 8±0.001 21)0.014 7±0.001 3∑ MUFA3.133 1±0.221 61)1.291 9±0.144 1C18∶2(n-6c)30.694 9±0.006 21)0.268 9±0.018 5C20∶230.007 1±0.000 61)0.003 3±0.000 2C22∶230.011 9±0.000 91)0.006 8±0.000 9C18∶3(n-3)30.146 8±0.010 81)0.058 5±0.004 7C18∶3(n-6)30.002 7±0.000 21)0.000 9±0.000 1C20∶3(n-3)30.012 9±0.000 90.010 6±0.000 82)C20∶3(n-6)30.007 5±0.000 51)0.004 9±0.000 3C20∶4(n-6)30.021 9±0.001 80.021 3±0.002 62)C20∶5(n-3)(EPA)0.014 5±0.001 10.014 3±0.001 52)C22∶6(n-3) (DHA)0.130 4±0.002 70.129 1±0.003 52)∑ PUFA1.102 4±0.018 61)0.530 7±0.056 6∑(n-3)0.304 6±0.013 51)0.212 4±0.006 3∑(n-6)0.727 1±0.004 71)0.296 2±0.019 5∑(n-3)/∑(n-6)0.418 90.736 2
1)S0与S15组比较,P<0.05,有统计学差异;2)S0与S15组比较,P>0.05,无统计学差异.
3 讨论
饥饿处理期间卵形鲳鲹依靠消耗自身营养物质储备维持生命活动所需的能量代谢,在一定程度上可以由形态学的指标变化客观地反映[22-23].本研究结果显示,肥满度、脏体比、肝体比均随饥饿时间延长呈下降趋势.与S0组相比,S15组脏体比降幅23.42%,肝体比降幅23.53%.其中脏体比S5组与S0组间有统计学差异(P<0.05),其后S10和S15组虽然持续降低但与S5组间无统计学差异(P>0.05);肝体比S5组与S0组间无统计学差异,而S15组、S10组与S0组间均有统计学差异(P<0.05),表明不同饥饿处理阶段内脏组织(主要是内脏脂肪组织)和肝脏组织交替作为主要能源,其中S5组由内脏组织供能为主,S10和S15组由肝组织供能为主.经济效益方面饥饿处理后商品鱼体虽然质量下降,即体质量损失,但由品质改良带来的产品附加值的提升得到更多补偿.
生化组成指标是对鱼体营养状态变化更为具体的反映.随饥饿时间延长,肌肉组织中水分和灰分质量分数逐渐升高而粗蛋白和粗脂肪质量分数逐渐降低,表明粗蛋白和粗脂肪被消耗分解,而水分和无机物质量分数相对增加,该4种指标变化趋势与对草鱼(Ctenopharyngodonidellus)[23]和斑点叉尾鮰(Ictaluruspunctatus)[24]的饥饿研究结果相似,但是变化程度则在不同鱼种之间呈现较大差异,饥饿过程中卵形鲳鲹肌肉组织粗蛋白质量分数呈下降趋势,但组间无统计学差异(P>0.05),而粗脂肪S5组与S0组间有统计学差异,提示粗脂肪为饥饿期间的主要能量来源,对斑点叉尾鮰的研究结果显示粗蛋白和粗脂肪同时作为主要能源物质在饥饿处理期间为鱼体提供能量[24].由于蛋白质和脂肪质量分数与鱼肉品质紧密相关,并且造成明显的口感和风味差异,因此有必要针对多种主要养殖鱼类开展更为细致的鱼肉品质改良相关研究.本研究对卵形鲳鲹肝脏组织生化组成分析表明,肝组中粗蛋白和粗脂肪质量分数在饥饿处理过程中均显著下降,与形态学分析肝体比指数变化互相印证.
品质构成参数是衡量肉品品质的主要指标[25],特别是对于肉质细嫩、容易受加工处理过程影响的鱼类,品质构成参数在反映肉质软硬程度和弹性等方面更为灵敏[26].本研究结果显示,随饥饿处理时间延长弹性指数和凝聚性指数呈升高趋势,而硬度呈降低趋势且组间有统计学差异(P<0.05),与对鲫鱼(Carassiusauratus)[26]、异育银鲫(CarassiusAuratusGibelio)[27]、斑点叉尾鮰[24]等研究有相似结果.鱼肉品质与鱼体的生长存活环境紧密相关,在野生环境中鱼类大部分情况下需要通过追逐才能捉获食物,而池塘环境中鱼类更多时间处于相对饱足状态,因此野生鱼类个体形态上相对瘦长,游泳能力也相对更强.本研究品质改良过程中采取持续不间断循环流水刺激,保证鱼体始终处于运动状态,运动能力锻炼与提升相应造成肌肉凝聚性指数和弹性指数升高,改良后鱼肉品质更接近于野生状态个体.
不同氨基酸种类在生物体类参与不同生理代谢过程.对鮸鱼(Miichthysmiiuy)研究显示天冬氨酸、甘氨酸等种类在饥饿过程中持续降低,苏氨酸、丝氨酸等种类先下降后上升,缬氨酸呈波浪型变化趋势[28].综观饥饿处理相关文献,不同氨基酸种类的质量分数变化存在明显种间差异性.本研究结果显示,与S0组相比,S15组缬氨酸、蛋氨酸、异亮氨酸、苯丙氨酸等的质量分数有所降低,而苏氨酸、天冬氨酸、甘氨酸等质量分数有所升高但无统计学差异(P>0.05),精氨酸质量分数在15 d饥饿处理之后升高12.8%.精氨酸作为一种条件性必需氨基酸,正常情况下大部分成年动物可自身合成,对幼年动物却是维持最佳生长的必需氨基酸[29].除一般营养功能外,精氨酸还参与调节脂质和蛋白质代谢[30],原因可能是精氨酸刺激某些酶类的分泌,从而调控脂肪酸合成和分解代谢途径[31],具体可能是精氨酸调节鱼类肝脏p38 MAPK 信号通路活性[32].动物试验显示摄入外源精氨酸可以促进脂肪分解,降低机体脂肪质量分数[33].医学研究显示提高精氨酸供给可以提高男性生育能力,若成年男子食品中连续9 d精氨酸缺乏,其精子数量和活力降低90%[34].此外,精氨酸在人类胚胎发育进程中对胎盘供血、着床和后期蛋白质合成中亦具有举足轻重的作用,近年来医学上越来越多地采用精氨酸以减少胚胎早期死亡、改善妊娠期间胎儿成活情况和促进胎儿出生后生长发育[35].本研究饥饿处理后卵形鲳鲹肌肉组织中精氨酸质量分数升高可能与动物本身抗逆适应性有关,于人类反而造就优质蛋白源,有助于提高从鱼肉中摄取精氨酸数量,对于辅助治疗男性不育及促进胎儿发育具有重要意义.
脂肪是动物体内重要的储能物质.本研究检测到卵形鲳鲹肌肉组织中11种SFA经15 d饥饿处理后质量分数显著降低(P<0.05),MUFA共7种除C14 ∶1外降幅均有统计学差异(P<0.05),显示出循环水系统中饥饿处理对商品规格卵形鲳鲹鱼体脂肪酸质量分数的影响.肉类在加热过程中产生的香味成分主要是脂肪类物质,高质量分数的 PUFA 能显著地增加香味,同时在一定程度上反映肌肉的多汁性[36].本研究结果表明15 d饥饿处理造成部分PUFA种类质量分数降低,其中高不饱和脂肪酸种类如C20∶4、C20∶5、C22∶6质量分数有所降低但无统计学差异,即饥饿处理后SFA和MUFA的显著降低使得PUFA在总脂肪酸中质量分数有明显提高,可更好地为人类摄食利用.∑(n-3)/∑(n-6)脂肪酸比值越高,间接说明脂肪酸组成更加适合人体营养需求[37].本研究结果显示饥饿处理造成卵形鲳鲹肌肉组织∑(n-3)/∑(n-6)比值从0.418 9升高至0.736 2,升幅达75.7%,说明短期饥饿处理大幅改良了卵形鲳鲹肌肉中高不饱和脂肪酸的组成.
综上,经历15 d循环水系统的饥饿处理,卵形鲳鲹形态上明显趋瘦,体内粗脂肪质量分数显著降低,肌肉组织凝聚性和弹性指数升高,精氨酸质量分数升高12.8%,∑(n-3)/∑(n-6)比值升高,饥饿处理过程从多个方面提升了商品规格卵形鲳鲹鱼肉品质.
