抑菌剂浸泡液对冷藏高白鲑鱼肉腐败菌的抑制效果
2018-09-20由高铭陈欣然赵前程马永生
由高铭,陈欣然,赵前程,马永生,李 萌,马 壮
( 1.大连海洋大学 食品科学与工程学院,辽宁 大连 116023; 2.新疆赛湖渔业科技开发有限公司,新疆 博乐 833400 )
新疆赛里木湖养殖的高白鲑(Coregonuspeled)富含营养成分,具有较高的食用价值和经济价值,目前以冰鲜产品为主运输至北京、上海等消费地域[1]。细菌的繁殖代谢是导致鱼肉等水产品腐败变质的重要因素,但只有少数细菌能适应贮藏环境快速繁殖成为优势腐败菌,主导腐败过程[2-3]。可将水产品浸泡于山梨酸钾、乳酸链球菌素、茶多酚或ε-聚赖氨酸等溶液,使抑菌剂吸附至水产品表面后,再加以冷藏达到抑菌防腐的目的[4-7]。分析不同抑菌剂对冷藏鱼肉中微生物的影响,可利用非线性模型如修正Gompertz方程和Baranyi & Robertz方程,它们是常见的预测食品货架期的微生物动力学生长模型,其各项拟合生长参数如延滞期和最大比生长速率等因子可指示抑菌效果[7]。目前,常见化学抑菌剂山梨酸钾和生物抑菌剂ε-聚赖氨酸盐在冷藏鱼肉基质中抑菌性能的研究较少,同时缺少不同浸泡技术对其抑菌性能影响的研究。因此,笔者对冷藏高白鲑鱼肉的腐败菌进行分离鉴定,后续采用山梨酸钾和ε-聚赖氨酸盐浸泡液经真空浸泡和常压浸泡对高白鲑鱼肉进行抑菌处理,利用非线性模型拟合生长参数,分析山梨酸钾和ε-聚赖氨酸盐浸泡液对鱼肉基质中腐败菌的抑菌活性,为后期冷藏鱼肉防腐抑菌剂的开发和利用奠定研究基础。
1 材料与方法
1.1 原料与试剂
高白鲑由新疆赛湖渔业科技开发有限公司提供。
营养琼脂、假单胞菌CFC选择培养基基础、结晶紫中性红胆盐葡萄糖琼脂、CFC添加剂(北京陆桥技术股份有限公司);铁琼脂(青岛海博生物技术有限公司);即用PCR扩增试剂盒(Taq)[生工生物工程(上海)股份有限公司];三氯乙酸(天津科密欧化学试剂有限公司);山梨酸钾(宁波王龙科技股份有限公司);ε-聚赖氨酸盐(浙江新银象生物工程有限公司)。
1.2 仪器与设备
真空干燥机(上海森信实验仪器有限公司);拍打式无菌均质机(宁波新芝生物科技有限公司);电热恒温培养箱(上海森信实验仪器有限公司);全自动凯氏定氮仪(意大利VELP公司);洁净工作台(苏州安泰空气技术有限公司)。
1.3 方法
1.3.1 原料处理
鲜活高白鲑(每条约600 g/尾)从新疆赛里木湖捕捞后立即宰杀,去头、去内脏、去皮洗净后沿鱼背脊将鱼肉平均分为两片,分装至无菌包装袋与碎冰1∶1的比例放置于泡沫箱内,冷链空运至大连。在无菌超净台内,将鱼肉进行分割处理,在无菌水浸泡清洗,取出沥干,置于无菌袋中密封,4 ℃保存。
1.3.2 冷藏高白鲑腐败菌的分离和纯化
高白鲑鱼肉在4 ℃保藏至第8 d时色泽发黄,肉质无弹性、发黏,汁液流失严重,有明显臭味,感官判断为腐败末期,此时无菌取样10 g,加入90 mL无菌生理盐水,均质后,用无菌生理盐水进行10倍稀释,选取合适稀释梯度,取100 μL稀释液在营养琼脂平板上涂布。平板倒置于恒温培养箱内,30 ℃培养72 h。将平板中细菌重复至少3次平板划线,得到纯化后的腐败菌单菌落。
1.3.2 腐败菌分子生物学鉴定
采用沸水浴法提取细菌DNA,构建PCR扩增体系,利用如下PCR扩增条件进行扩增:94 ℃变性5 min,进行35个循环(94 ℃ 30 s,55 ℃ 30 s,72 ℃ 90 s),72 ℃延伸 10 min。取9 μL PCR扩增产物进行1%琼脂糖凝胶电泳,若PCR扩增条带单一明亮,则将PCR扩增产物送至生工生物工程(上海)股份有限公司进行测序工作;16S rDNA序列利用美国国立生物技术信息中心中的Blast进行序列比对。
1.3.3 鱼肉基质的抑菌试验
0.5%的山梨酸钾和ε-聚赖氨酸盐溶液分别经0.45 μm无菌滤器过膜除菌,4 ℃保存备用。将新鲜鱼肉置于无菌烧杯内,按照鱼肉和抑菌剂1∶2(m/V)的比例加入山梨酸钾或ε-聚赖氨酸盐溶液,分别进行真空和常压浸泡处理,得到真空浸泡山梨酸钾鱼肉样品,真空浸泡ε-聚赖氨酸盐鱼肉样品,常压浸泡山梨酸钾鱼肉样品和常压浸泡ε-聚赖氨酸盐鱼肉样品。以无菌水浸泡液为空白对照组,在相同条件下浸泡鱼肉,获得真空浸泡空白对照组和常压浸泡空白对照组。其中,真空浸泡包括两个步骤,在真空度0.08 MPa环境中浸泡处理15 min;恢复至常压条件,再浸泡15 min。
浸泡完成后,在超净台内将鱼肉取出放置无菌托盘内,无菌纱布吸干水分,将鱼肉分装到无菌袋内,每袋约10 g,密封后4 ℃避光保存。在第0、2、4、6、8 d时测定鱼肉的微生物指标变化。
1.3.4 细菌总数和腐败菌测定
根据国标GB/T 4789.2—2010[8]测定:称取10 g鱼肉置盛有90 mL生理盐水的无菌均质袋中,用拍击式均质器拍打1~2 min,制成1∶10的样品匀液。用无菌盐水进行10倍梯度稀释,吸取100 μL不同稀释度的样品溶液,分别均匀涂布于营养琼脂、结晶紫中性红胆盐琼脂培养基、铁琼脂、CFC选择性培养基平板内,培养48 h,测定菌落总数、肠杆菌、产硫菌和假单胞菌数目。
1.3.5 微生物拟合生长参数的测定
修正Gompertz模型应用Origin软件中的Nonlinar Curve Fit功能进行拟合,其中修正Gompertz方程为:
Nt=N0+(Nmax-N0)×exp{-exp[2.718μmax/(Nmax-N0)(Lag-t)+1]}
(1)
式中,Nmax、N0为最大和初始细菌数量对数值,Lag为延滞期,μmax为最大比生长速率[9]。
Baranyi & Robertz模型利用Combase在线软件中DMFit功能进行拟合,软件依据的方程为:
(2)
Y1=μmaxt+ ln[e-μmaxt-e-μmax(t+tlag)+e-μmaxtlag]
(3)
Y2= ln[1+10(Y0-Ymax)(e-μmax(t-λ)-e-μmaxtlag]
(4)
式中,Y0为细菌初始数量对数值,Ymax为细菌达到稳定期时数量的对数值,Lag为延滞期和μmax为最大比生长速率[10]。
采用残差标准误差和决定系数评价模型的拟合能力。
1.4 数据处理
数据取3次平行试验结果,以平均值±标准差表示。利用SPSS 19.0软件进行相关性、单因素方差分析和独立样本t检验等分析,其中P<0.05为显著性差异。采用OriginPro 8.5软件作图。
2 结 果
2.1 冷藏高白鲑腐败菌的分离鉴定
冷藏高白鲑鱼肉贮藏末期,根据肉眼观察菌落形态、菌落颜色、隆起状态、边缘和表面状态,筛选出存在差异的27株菌落,16S rDNA测序比对分析的结果见表1。冷藏高白鲑鱼肉贮藏末期的27株腐败菌中包含9株沙雷氏菌属(Serratia)细菌,5株拉恩氏菌属(Rahnella)细菌,3株哈夫尼菌属(Hafnia)细菌,2株克雷伯菌属(Klebsiella)细菌,即分离筛选的腐败菌中70%属于肠杆菌科;另外,鉴定出4株细菌为弧菌科希瓦氏菌属(Shewanella)的腐败希瓦氏菌(S.putrefaciens),占总比例的16%;其余为芽孢杆菌属(Bacillus)和肉食杆菌属(Carnobacterium)细菌。

表1 冷藏高白鲑鱼腐败菌16S rDNA 鉴定结果
2.2 冷藏高白鲑鱼肉中菌落总数的动态变化
真空浸泡对照组和常压浸泡对照组中鱼肉的初始菌落数对数分别为4.34 log(cfu/g)和4.44 log(cfu/g),随时间的延长,鱼肉中菌落总数对数均呈现上升趋势(图1)。利用修正Gompertz方程和Baranyi & Robertz方程拟合冷藏鱼肉中细菌的生长参数,结果见表2。
两种模型拟合的真空浸泡对照组和常压浸泡对照组生长参数如细菌总数初始密度对数,最高密度对数以及细菌生长延滞期均无差异,说明真空浸泡处理方式对鱼肉的细菌总数增长无影响。山梨酸钾和ε-聚赖氨酸盐浸泡液能有效延长鱼肉中细菌的延滞期,并降低细菌达到稳定期时的最高密度,表明山梨酸钾和ε-聚赖氨酸盐能够有效延缓冷藏高白鲑鱼细菌总数的增长。然而,真空浸泡试验组鱼肉与常压浸泡试验组鱼肉在各项生长参数上并无明显优势。
2.3 冷藏高白鲑鱼肉中肠杆菌的动态变化
利用修正Gompertz方程和Baranyi & Robertz方程拟合冷藏高白鲑鱼肉中肠杆菌动态变化,各项拟合生长参数见表3。
对照组鱼肉样品中肠杆菌的初始菌落数目对数均低于2 log(cfu/g),表明产品加工过程中卫生条件控制较好。依据拟合效果更好的修正Gompertz模型,经山梨酸钾和ε-聚赖氨酸盐浸泡处理的鱼肉中肠杆菌初始菌落数对数均低于对照组,延滞期延长,然而细菌最大比生长速率提高,表明山梨酸钾和ε-聚赖氨酸盐浸泡液在贮藏初期能有效抑制鱼肉中肠杆菌的活性,但在后期抑制效果较差。真空浸泡与常压浸泡试验组肠杆菌的各项生长参数之间并无显著差异,表明真空浸泡方式未能提高抑菌剂的抑菌活性。

表2 冷藏高白鲑鱼肉细菌总数动力学生长模型的各项参数
注:Lag为延滞期,μmax为最大比生长速率,RSE为残差标准误差,r2为决定系数,Y0为细菌初始数量对数值,Ymax为细菌达到稳定期的数量对数值.下同.

表3 冷藏高白鲑鱼肉肠杆菌动力学生长模型的各项拟合参数


图1 冷藏高白鲑鱼肉菌落总数变化情况 a.真空浸泡处理, b.常压浸泡处理. VP.真空浸泡, AP.常压浸泡, C.空白对照, PS.山梨酸钾, PLH.ε-聚赖氨酸盐.
2.4 冷藏高白鲑鱼肉中产硫菌的动态变化
山梨酸钾浸泡液能有效抑制鱼肉中产硫菌的生长繁殖,在鱼肉贮藏过程中,产硫菌数目均低于2 log(cfu/g)。对于空白组和ε-聚赖氨酸盐试验组中产硫菌的生长变化,修正Gompertz方程比Baranyi & Robertz方程的残差标准误差值低,决定系数值高,整体拟合效果更好(表4)。依据修正Gompertz模型拟合结果,ε-聚赖氨酸盐试验组鱼肉中产硫菌的延滞期比对照组延长,但最大比生长速率未降低,说明在贮藏初期ε-聚赖氨酸盐对贮藏鱼肉基质中产硫菌的活性有抑制作用。然而,真空浸泡和常压浸泡试验组鱼肉之间的产硫菌各项生长参数均无显著性变化,表明浸泡方式对抑菌剂抑制产硫菌的效果无影响。
2.5 冷藏高白鲑鱼肉中假单胞菌的动态变化
修正Gompertz方程比Baranyi & Robertz方程的残差标准误差值低,决定系数值高,说明前者能更好的拟合鱼肉中假单胞菌的生长变化(表5)。依据修正Gompertz模型拟合结果,常压浸泡空白组鱼肉中假单胞菌最大比生长速率为(0.55±0.19)/d,真空浸泡对照组为(1.26±0.03)/d,表明真空浸泡导致鱼肉中假单胞菌的生长速率增大。同时,真空浸泡试验组中假单胞菌的延滞期或最大比生长速率要差于常压浸泡组,表明真空浸泡未提高ε-聚赖氨酸盐和山梨酸钾对鱼肉基质中假单胞菌的抑制能力。常压浸泡状态下,山梨酸钾试验组鱼肉中的假单胞菌生长速率和最高细菌密度对数均低于对照组,ε-聚赖氨酸盐试验组的延滞期要长于对照组,说明山梨酸钾对鱼肉基质中的假单胞菌有抑制效果,而ε-聚赖氨酸盐仅在贮藏初期有抑菌效果。

表4 冷藏高白鲑鱼肉产硫菌动力学生长模型的各项拟合参数
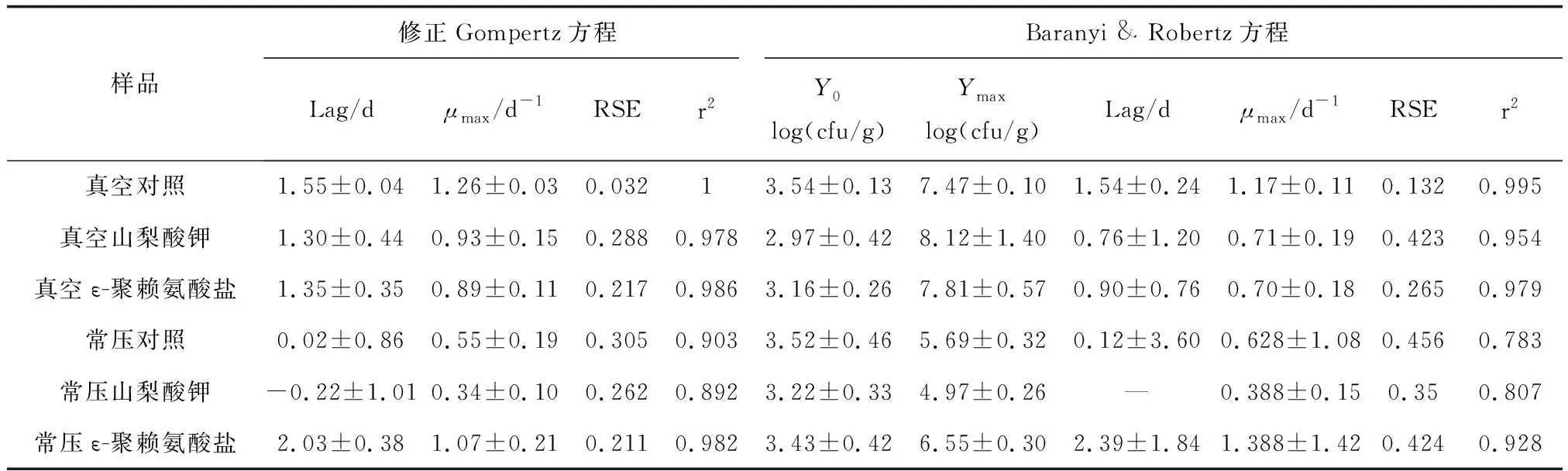
表5 冷藏高白鲑鱼肉假单胞菌动力学生长模型的各项拟合参数
3 讨 论
3.1 冷藏高白鲑鱼肉的腐败菌
引起水产品腐败的主要原因是外源性腐败菌的繁殖,而在不同的贮藏条件下,导致水产品腐败变质的腐败菌种类不同[11]。Sade等[12]的研究表明,气调包装的肉制品中,肠杆菌占总细菌的60%,其中沙雷氏菌属如泉居沙雷氏菌(S.fonticola)、格氏沙雷氏菌、液化沙雷氏菌和变形斑沙雷氏菌等占肠杆菌的42%,哈夫尼菌属占40%。沙雷氏菌属也是真空包装和气调包装的大西洋鲑(Salmosalar)鱼肉中的优势腐败菌[13]。产硫菌,尤其是腐败希瓦氏菌,为淡水鱼低温冷藏过程中的优势腐败菌[14]。与上述研究结果一致,本研究中分离获得的腐败菌中肠杆菌占总细菌的70%,产硫菌腐败希瓦氏菌占16%,可能是导致冷藏高白鲑鱼片产品腐败变质的主要腐败菌。
有氧冷藏条件下,假单胞菌属为生鲜水产品中常见的腐败菌,产生大量醛、酮、酯和有异味的挥发性产物[15]。采用传统检测腐败菌方法未检出假单胞菌,主要是因为基于培养基培养检测腐败菌方法具有一定的局限性[16-18]。为更有效的分析腐败菌的变化情况,在后续的试验中,笔者将对冷藏高白鲑鱼肉中的肠杆菌、产硫菌和假单胞菌均进行选择性培养,探究抑菌剂对主要腐败菌的抑制情况。
3.2 非线性模型拟合微生物生长变化
残差标准误差值越小,决定系数值越高,模型拟合效果越好,表2中各组细菌的拟合模型残差标准误差值均低于0.38,且决定系数均在0.97以上,表明修正Gompertz方程和Baranyi & Robertz方程均能较好地反映出各组冷藏鱼肉中细菌总数的生长情况。由表3~表5中残差标准误差值和决定系数值可知,修正Gompertz方程较Baranyi & Robertz方程的整体拟合效果更好,能够更准确的描述鱼肉中腐败菌的生长动态。而丁婷等[19]的研究表明,Baranyi & Robertz方程来描述冷藏三文鱼片中腐败菌的生长动态比修正Gompertz方程更适合。这可能是由不同的鱼肉基质导致腐败菌生长状态具有差异性所致。
3.3 抑菌剂对鱼肉基质中腐败菌的抑制效果
经山梨酸钾或ε-聚赖氨酸盐浸泡处理后,冷藏高白鲑鱼肉的细菌总数均低于对照组鱼肉,与利用山梨酸钾或ε-聚赖氨酸浸泡液有效降低冷藏扁裸颊鲷(Lethrinuslentjan)[20]、冷藏花鲈(Lateolabraxjaponicus)[6]鱼肉中的细菌总数的研究结果一致。
周婷等[21-22]研究表明,0.5‰的山梨酸钾能有效抑制水产品中腐败菌的繁殖。在本研究中,0.5%的山梨酸钾浸泡液能有效抑制冷藏高白鲑鱼肉中希瓦氏菌和假单胞菌的繁殖,然而,鱼肉中肠杆菌的最大比生长速率高于对照组。Gençcelep等[23]的研究也发现,山梨酸钾浸泡液对冷藏珍珠鲻鱼(Chalcalburnustarichi)中肠杆菌的抑菌能力与浸泡液含量密切相关,5%山梨酸钾浸泡液能有效抑制肠杆菌的繁殖,但1%含量的抑菌剂在贮藏后期无显著抑菌效果。
经0.5%的ε-聚赖氨酸盐浸泡液处理后,冷藏高白鲑鱼肉中的肠杆菌、产硫菌和假单胞菌最大比生长速率均高于对照组。这可能是鱼肉基质中ε-聚赖氨酸盐含量较低的缘故。张全景等[24]发现ε-聚赖氨酸在低质量浓度条件下可能具有杀灭和抑制共同存在的致伤作用,并不能完全杀灭细菌。贮藏末期,受伤害程度不同的微生物经过不同时间的修复能够重新繁殖。
3.4 浸泡方式对抑菌剂抑菌效果的影响
抑菌物质在水产品特别是鱼肉表面的扩散易受到食品基质的影响,导致抑菌活性减弱,通过增强抑菌物质的扩散性,可提高其在食品基质中的抑菌性能。与传统的常压浸泡技术相比,真空浸泡可将溶液快速均匀的渗透到食品内部并且可减少食品中水分和营养物质的流失[25]。Andrés-Bello等[26]发现,真空浸泡能有效提高乳酸菌和乳酸链球菌素在冷藏金头鲷(Sparusaurata)鱼肉中的抑菌活性,有效抑制菌落总数和肠杆菌的繁殖;但在本试验中,真空浸泡并未有效提高山梨酸钾和ε-聚赖氨酸盐在鱼肉基质中对菌落总数、肠杆菌和产硫菌的抑制效果,反而加速了假单胞菌的繁殖。这可能与抑菌剂的种类以及鱼肉本身的组织结构不同有关。
3.5 小结
冷藏高白鲑鱼肉贮藏末期的腐败菌主要由肠杆菌科的沙雷氏菌属以及产硫菌(腐败希瓦氏菌)构成。修正Gompertz方程对冷藏高白鲑鱼肉中腐败菌繁殖变化的拟合效果优于Baranyi & Robertz方程,且通过拟合生长参数得知山梨酸钾和ε-聚赖氨酸盐能有效抑制冷藏高白鲑基质中细菌总数和腐败菌的生长繁殖,但真空浸泡不能促进抑菌剂对鱼肉中腐败菌的抑菌活性,反而导致假单胞菌的快速繁殖。后续可采用非线性模型分析不同含量抑菌剂浸泡或喷淋处理冷藏高白鲑鱼肉的抑菌效果,筛选符合国家标准要求的防腐剂和适宜含量,为延长冷藏高白鲑鱼肉的货架期,提高产品质量和安全性奠定理论基础。
