晚期糖基化终末产物受体对宫颈癌细胞生物学行为的影响
2018-09-18周璐璐朱雪洁李如意朱雪琼
周璐璐 朱雪洁 李如意 朱雪琼
晚期糖基化终末产物受体(receptor for advancedglycation end products,RAGE)是一种具有多配体的跨膜信号转导受体[1]。研究发现RAGE在卵巢癌[2]、乳腺癌[3]、子宫内膜癌[4]及结直肠癌[5]等肿瘤中的表达明显上调,其异常表达被认为与肿瘤的发生、发展相关。本课题组前期研究发现RAGE蛋白在正常宫颈鳞状上皮组织、宫颈上皮内瘤变Ⅰ级、Ⅱ级、Ⅲ级以及宫颈鳞癌中的表达逐渐增高[6]。但是,RAGE对宫颈癌细胞生物学行为的影响及其机制尚未明确。因此,本研究通过转染pLenti-C-mGFP-RAGE质粒构建RAGE基因稳定过表达的宫颈鳞癌SiHa细胞株,观察RAGE表达改变对宫颈鳞癌SiHa细胞增殖、凋亡及裸鼠成瘤能力的影响。
1 材料和方法
1.1 材料 SiHa细胞及人胚肾细胞293T细胞购于中国科学院上海细胞研究所细胞库,DMEM培养基及FBS均购于美国Gibco公司。大肠杆菌菌株DH5α购于北京天根生物科技有限公司。RAGE抗体购于美国Santa-Crutz公司,增殖细胞核抗原(proliferating cell nuclear antigen,PCNA)、B 细胞淋巴瘤/白血病-2(B cell lymphoma/lewkmia-2,Bcl-2)、Bcl-2 相关 X 蛋白(Bcl-2 associated X protein,Bax)、与 Bcl-2相互作用的细胞死亡调节因子(Bcl-2 interacting mediator of cell death,Bim)抗体均购于美国Cell Signaling Technology公司。慢病毒空载体pLenti-C-mGFP-vector及pLenti-C-mGFP-RAGE质粒均购于美国Origene公司。CCK-8试剂盒购于日本同仁化学研究所;PE/7-AAD购于美国BD公司。
1.2 方法
1.2.1 细胞培养 将SiHa细胞及人胚肾细胞293T细胞加到改良DMEM培养基中,放入37℃、5%CO2细胞培养箱,每隔48h换液1次。
1.2.2 质粒转染
1.2.2.1 pLenti-C-mGFP-RAGE过表达慢病毒的包装、收集 人胚肾细胞293T细胞用含10%FBS的DMEM培养基培养,接种于T75培养瓶,待细胞汇合度达60%~80%,用慢病毒包装质粒psPAX2、pMD2.G以及目的质粒共转染,参照Lipofectamine3000说明书进行转染。转染6h后换液。转染48h后观察细胞状态,并在荧光显微镜下观察绿色荧光蛋白(GFP)表达情况并观察转染效率。用0.45μm的无菌聚偏氟乙烯(PVDF)针式过滤器过滤收集病毒液。
1.2.2.2 细胞转染及分组 将细胞进行分组培养,空白对照组为仅添加凝聚胺,空载体组为添加凝聚胺+转染空载体质粒,过表达组为添加凝聚胺+转染RAGE过表达质粒。每个培养皿中加入5×105个SiHa细胞,培养24h,细胞汇合度约50%,用终浓度为8μg/ml凝聚胺的培养基培养细胞30min,再加入1ml病毒液感染细胞,24h后换液,48h后荧光显微镜下观察转染效率。
1.2.3 Western blot法检测 RAGE、PCNA、Bcl-2、Bax及Bim蛋白表达水平 分别提取各组细胞总蛋白,测定蛋白浓度。取40μg蛋白上样,常规十二烷基硫酸钠聚丙烯酰胺凝胶电泳、转膜。用5%脱脂奶粉封闭2h后,分别加入 RAGE(1∶2 000)、PCNA(1∶2 000)、Bcl-2(1∶1 000)、Bax(1∶1 000)、Bim(1∶1 000)及 GAPDH(1∶2 000)抗体,4℃孵育过夜,洗膜后分别加入辣根过氧化酶标记IgG抗体,室温下孵育2h,洗膜后将显影液加于PVDF膜上。用凝胶成像系统扫描并分析。以目标蛋白与内参蛋白条带灰度比值计算各组蛋白的相对表达量。
1.2.4 CCK-8法检测细胞增殖情况 SiHa细胞用新鲜完全培养基重悬后,调整细胞密度为5×104/L,向96孔板加入细胞悬液100μl。置于37℃、5%CO2的细胞培养箱中培养48h后,每孔加入CCK-8试剂10μl+DMEM 90μl,避光孵育2h。取出96孔板置于酶标仪上测定450nm处的吸光度(OD)值,每组实验重复3次。1.2.5 流式细胞仪检测细胞凋亡情况 用不含乙二胺四乙酸的胰酶消化各组细胞,1 000r/min离心5min,移入流式管,用预冷的PBS洗2次。每管加入500μl 1×binding buffer悬浮细胞,使细胞密度约为 1×106/ml。向流式管中加入5μl PE/5μl 7-AAD,混匀后避光孵育15min。1h内上流式细胞仪检测结果,每组实验重复3次。
1.2.6 裸鼠皮下成瘤 将过表达组及空载体组SiHa细胞按5×106/只接种在裸鼠背部右侧腋下皮下。待细胞成瘤后,每隔4d测量皮下移植瘤体积,观察移植瘤生长情况,8周后处死裸鼠,称量瘤体质量。
1.3 统计学处理 采用SPSS 19.0统计软件。计量资料以表示,多组间比较采用单因素方差分析,两两比较采用LSD-t法。P<0.05为差异有统计学意义。
2 结果
2.1 转染pLenti-C-mGFP-RAGE质粒后SiHa细胞中RAGE蛋白表达水平 Western blot法检测发现,过表达组可检测GFP-RAGE融合蛋白,表明pLenti-C-mGFP-RAGE质粒成功转入SiHa细胞中,RAGE蛋白表达水平约为空载体组的(2.13±0.57)倍,见图1。

图1 转染pLenti-C-mGFP-RAGE质粒后SiHa细胞中RAGE蛋白表达的电泳图
2.2 RAGE蛋白表达增高对SiHa细胞增殖的影响 CCK-8法检测发现,过表达组SiHa细胞的增殖力均明显高于空白对照组及空载体组(均P<0.05),空白对照组及空载体组SiHa细胞的增殖力比较差异无统计学意义(P>0.05),提示RAGE蛋白表达上调后促进了SiHa细胞的增殖,见表1。

表1 RAGE蛋白表达增高对SiHa细胞增殖的影响
2.3 RAGE蛋白表达增高对SiHa细胞凋亡的影响 流式细胞仪检测发现,过表达组SiHa细胞的凋亡率为(7.10±2.67)%,明显低于空白对照组的(18.33±2.06)%及空载体组的(19.81±2.49)%(均P<0.05),空白对照组及空载体组SiHa细胞的凋亡率比较差异无统计学意义(P>0.05),提示RAGE上调后抑制了SiHa细胞的凋亡,见图2。
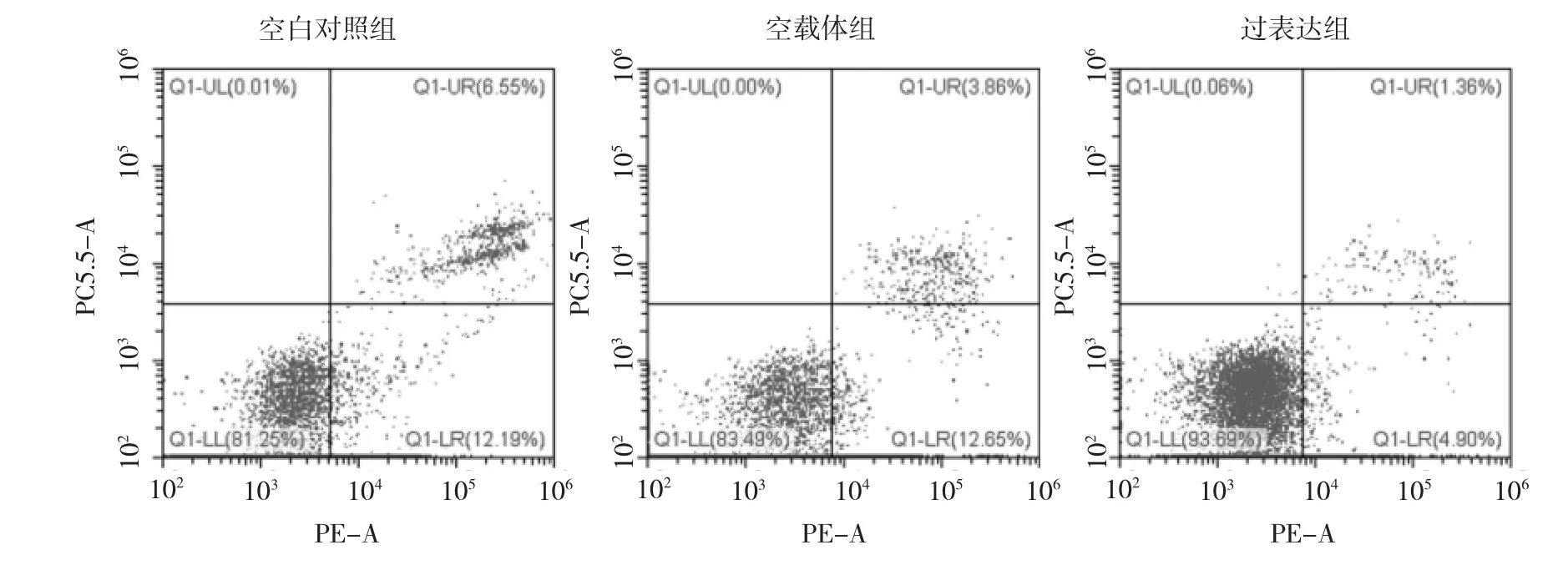
图2 3组SiHa细胞的流式细胞图
2.4 RAGE蛋白表达增高对SiHa细胞中增殖及凋亡相关蛋白表达的影响 Western blot法检测发现,过表达组PCNA及Bcl-2蛋白表达水平均明显高于空白对照组及空载体组(均P<0.05),而Bax/Bcl-2表达水平低于均明显低于空白对照组及空载体组(均P<0.05),3组Bax及Bim蛋白表达水平比较差异均无统计学意义(均P>0.05),见表2及图3。
2.5 RAGE蛋白表达增高对裸鼠宫颈癌皮下成瘤的影响 过表达组裸鼠皮下瘤体体积为(1 599.47±247.97)mm3,瘤体质量为(1.67±0.35)g,显著高于空载体组的体 积(629.92±140.59)mm3和瘤体质量(0.74±0.42)g,差异
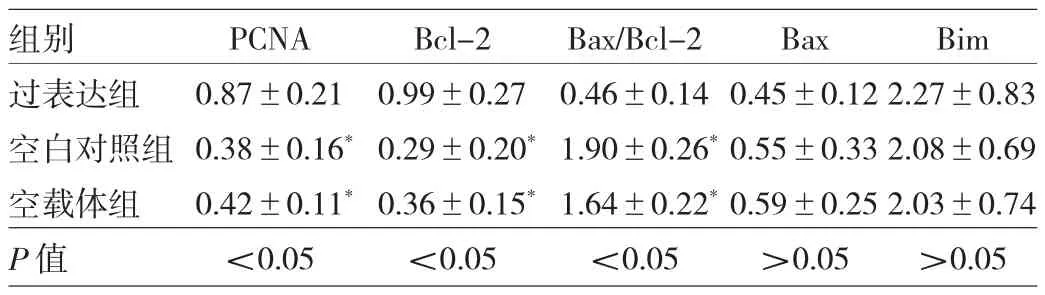
表2 RAGE蛋白表达增高对SiHa细胞中增殖及凋亡相关蛋白表达的影响
均有统计学意义(均P<0.05),见图4-5。
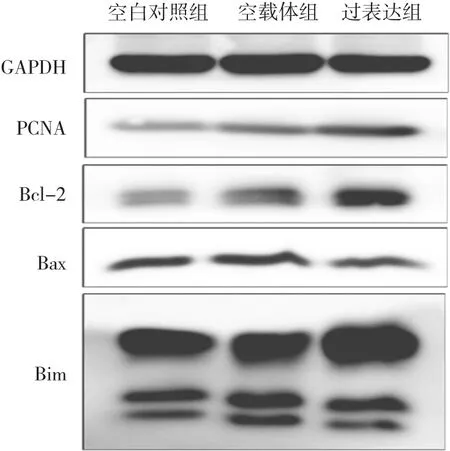
图3 RAGE蛋白表达增高后SiHa细胞中增殖及凋亡相关蛋白表达的电泳图

图4 不同RAGE蛋白表达2组裸鼠宫颈皮下瘤体体积的比较(a:空载体组瘤体图;b:过表达组瘤体图)

图5 不同RAGE蛋白表达2组裸鼠宫颈皮下瘤体体积的比较(与空载体组比较,*P<0.05)
3 讨论
宫颈癌是女性常见的恶性肿瘤,最新数据显示,2017年全美宫颈癌新发病例高达12 820例,死亡病例达4 210例[7]。Chang等[8]研究表明宫颈癌患病呈现年轻化趋势,宫颈癌和癌前病变的发病率呈上升趋势,但宫颈癌的具体发生、发展机制尚未明确。
RAGE蛋白是细胞表面分子免疫球蛋白超家族成员之一,可与晚期糖基化终末产物、高迁移率蛋白B1、溶血性磷脂酸、S-100/钙粒蛋白和β淀粉样肽等配体相互作用,激活细胞内相关信号通路,导致炎症和癌症的发生、发展[9-10]。Iotzova-Weiss等[11]利用慢病毒包裹的RAGE特异shRNA转染原代角质细胞降低RAGE基因的表达量,发现RAGE基因低表达组的细胞生长速率较阴性对照组下降了80%,提示RAGE基因水平下调后可抑制角质细胞的增殖能力,表明了RAGE基因与鳞状上皮细胞的增殖相关。Jin等[12]通过免疫共沉淀实验验证了S-100A14与RAGE在食管鳞癌细胞中相互结合并促进食管鳞癌细胞的增殖能力。Radia等[13]下调乳腺癌细胞中RAGE蛋白表达量后,发现转录因子NF-κB p65、细胞增殖标志物PCNA和G1/S-特异性周期蛋白-D1(Cyclin D1)的表达量也都随之下降,说明RAGE对乳腺癌细胞的增殖具有一定的促进作用。但关于RAGE对宫颈鳞癌细胞生物学行为影响的研究尚未见报道。
本研究应用慢病毒转染技术,使SiHa细胞中RAGE蛋白稳定过表达后,发现过表达组SiHa细胞的增殖能力较空白对照组显著升高,并检测到增殖相关蛋白PCNA的表达量升高,提示RAGE基因可促进宫颈鳞癌细胞增殖。同时,裸鼠皮下成瘤实验也发现RAGE基因具有促进宫颈鳞癌肿瘤生长的能力。
Lata等[14]利用17α乙炔基雌二醇作用乳腺癌细胞系MCF-7后,提高了细胞内雌激素受体相关受体γ的表达,随后增强了细胞的氧化应激,激活了转录因子和NF-κB,最终导致了RAGE蛋白表达明显增高。进一步研究发现,RAGE基因可激活细胞周期蛋白Cyclin D1的表达而促进细胞的增殖,同时RAGE基因可促进Akt磷酸化及抗凋亡蛋白Bcl-2的表达,不仅参与了细胞的增殖,也参与了抑制细胞的凋亡。Malik等[15]也曾提出RAGE基因可以通过激活细胞周期蛋白Cyclin D1、抗凋亡蛋白Bcl-2及自噬相关蛋白的表达,抑制肿瘤细胞的凋亡,从而提高肿瘤细胞的存活率。本研究发现RAGE具有促进宫颈癌细胞凋亡的能力,且该作用与抗凋亡蛋白Bcl-2相关,而与Bax和Bim无关。
