miR-495在骨肉瘤细胞中表达变化、对MG-63细胞增殖的影响及其机制
2018-08-03王美凤邢国英宫锦蕾孙英华夏海王复超张兆卿
王美凤,邢国英,宫锦蕾,孙英华,夏海,王复超,张兆卿
(1潍坊医学院附属益都中心医院,山东潍坊262500;2大连医科大学附属第一医院)
骨肉瘤是儿童和青少年最常见的原发性骨恶性肿瘤,其来源于间叶组织,多发生于长骨干骺端,易早期发生转移,恶性程度极高,预后较差,其发病机制目前仍不清楚。近年来,骨肉瘤患者的生存率得到一定提高,但化疗的耐药性及毒性反应较大,因此,探索骨肉瘤的发生发展机制,寻找有效的基因治疗靶点意义重大[1]。微小RNA(miRNA)是高度保守的内源性非编码小RNA,能够在转录后水平调控靶基因表达,参与肿瘤的发生、发展过程[2,3]。研究发现miR-495与恶性肿瘤的关系密切,在胃癌[4]、食管癌[5]、前列腺癌、非小细胞肺癌[6]、膀胱癌[7]等多种实体肿瘤中发挥着抑癌基因的作用。目前有关miR-495在骨肉瘤中表达情况的研究甚少,并且作用机制尚不明确。高迁移率族蛋白A2(HMGA2)是高迁移率族蛋白超家族重要成员之一,近年来被公认作为一种新的癌基因参与恶性肿瘤的发生发展[8]。本研究观察了miR-495在骨肉瘤细胞中的表达变化,及其对骨肉瘤MG-63细胞增殖的影响,并探讨可能的分子机制。
1 材料与方法
1.1 细胞与主要实验材料 人骨肉瘤细胞系Saos-2、U20S、MG-63及人成骨细胞系hFOB1.19均购自美国菌种保藏中心。Saos-2、U20S、MG-63细胞用含有10%胎牛血清的DMEM培养液培养。hFOB1.19细胞用含10%胎牛血清的DMEM/F12培养液培养。所有细胞均在37 ℃、5% CO2恒温培养箱中培养,定期换液,细胞融合至75%~80%时常规传代。miR-495 mimics、阴性对照mimics-NC、HMGA2表达质粒、miR-495及内参U6引物均由上海吉玛制药技术有限公司设计合成。TRIzol总RNA提取试剂盒及Lipofectamine2000转染试剂盒购自美国Invitrogen公司。逆转录试剂盒及real-time PCR试剂盒购自大连宝生物公司。real-time PCR仪购自瑞士Roche公司。双荧光素酶报告检测试剂盒购自美国Promega公司。CCK-8试剂盒购自上海碧云天生物技术有限公司。HMGA2单克隆抗体及辣根过氧化物酶标记的二抗均购自美国Santa Cruz公司。
1.2 不同骨肉瘤细胞系中miR-495检测 采用real-time PCR法。按照试剂盒说明书用TRIzol提取细胞总RNA,分光光度仪检测RNA的纯度及浓度,按逆转录试剂盒说明书逆转录得到cDNA。miR-495上游引物序列为5′-TCCGATTCTTCACGTGGTAC-3′,下游引物序列为5′-GTGCAGGGTCCGAGGT-3′;内参U6上游引物序列为5′-TCCGATCGTGAAGCGTTC-3′,下游引物序列为5′-GTGCAGGGTCCGAGGT-3′。以cDNA为模板,参照real-time PCR试剂盒说明书配置PCR反应体系。反应条件:94 ℃ 2 min,94 ℃ 20 s,60 ℃ 30 s,共40个循环。miR-495相对表达量以2-ΔΔCt表示。
1.3 miR-495对MG-63细胞增殖的影响观察
1.3.1 细胞分组及miR-495 mimics转染 采用脂质体介导法,转染前1 d将处于对数生长期的MG-63细胞消化、离心,以2×105/孔接种于6孔细胞培养板,待细胞融合度为50%~70%时进行转染。将MG-63细胞分为阴性对照组、miR-495 mimics组、miR-495 mimics+HMGA2组,分别转染mimics-NC、miR-495 mimics、miR-495 mimics+HMGA2表达质粒。
1.3.2 细胞中miR-495检测 采用real-time PCR法,具体操作参考“1.2”。转染后24h,miR-495 mimics组、miR-495 mimics+HMGA2组和阴性对照组细胞中miR-495的相对表达量分别为43.63±3.84、41.08±4.20、1.01±0.07。miR-495 mimics组和miR-495 mimics+HMGA2组细胞中miR-495相对表达量高于阴性对照组﹙P均<0.01﹚,证明转染miR-495 mimics能够明显提高MG-63细胞中miR-495的表达。
1.3.3 细胞增殖能力检测 采用CCK-8法。取转染后的各组细胞,加入胰酶消化后重悬细胞制备单细胞悬液,调整细胞密度为1×105/L。分别取200 μL的各组细胞悬液接种于96孔板,各组设5个复孔,于24、48、72、96 h后参照CCK-8试剂盒操作说明书向每孔加入10 μL的CCK-8溶液,置于37 ℃、5% CO2的培养箱中避光孵育2 h,采用酶标仪测定450 nm波长处的OD值,表示细胞增殖能力。
1.4 miR-495靶基因预测及对HMGA2的靶向调控作用观察
1.4.1 miR-495靶基因预测 应用靶基因在线预测数据库miRanda(www.microrna.org)并结合基因的功能分析进行miR-495的靶基因预测。
1.4.2 miR-495与靶基因的结合位点分析 采用双荧光素酶报告实验验证miR-495与靶基因的结合位点。将HMGA2的3′UTR的PCR扩增产物连接到pGL3质粒XbalⅠ酶切位点,构建野生型pGL3-HMGA2-wtUTR报告基因质粒。对miR-495与HMGA2的3′UTR结合处的碱基进行定点突变,突变后得到的新的质粒为突变型pGL3-HMGA2-mutUTR报告基因质粒。取MG-63细胞制备单细胞悬液,接种于96孔板,用Lipofectamine2000试剂进行转染,转染后分为HMGA2野生型质粒对照组(转染野生型pGL3-HMGA2-wtUTR报告基因质粒和mimics-NC)、HMGA2野生型质粒实验组(转染野生型pGL3-HMGA2-wtUTR报告基因质粒和miR-495 mimics)、HMGA2突变型质粒对照组(转染突变型pGL3-HMGA2-mutUTR报告基因质粒和mimics-NC)、HMGA2突变型质粒实验组(转染突变型pGL3-HMGA2-mutUTR报告基因质粒和miR-495 mimics)。四组转染24 h后裂解细胞,按照双荧光素酶活性检测试剂盒说明书操作,计算相对荧光酶活性。相对荧光酶活性=萤火虫荧光素酶活性值/海肾荧光素酶活性值。
1.4.3 细胞中HMGA2蛋白检测 采用Western blotting法。细胞分组与转染方法参照“1.3.1”,收集转染后的细胞,PBS洗涤细胞3遍,加入RIPA细胞裂解液冰上裂解30 min,离心收集上清液,BCA法测定蛋白浓度。每上样孔加入20 μg蛋白,进行10% SDS-PAGE电泳,将电泳分离后的蛋白质电转移至硝酸纤维素膜上,5%脱脂奶粉室温封闭2 h后,加入适当浓度的HMGA2一抗和β-actin一抗,4 ℃摇床孵育过夜,TBST洗膜3次后,再加入适当浓度的二抗室温孵育2 h,洗膜,加入ECL显影液显影、定影。蛋白的相对表达量以目的蛋白条带灰度值与内参条带灰度值的比值表示。
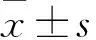
2 结果
2.1 不同骨肉瘤细胞系中miR-495表达比较 Saos-2、MG-63、U20S细胞及hFOB1.19细胞中miR-495的相对表达量分别为0.43±0.06、0.22±0.08、0.36±0.06、1.01±0.02。Saos-2、MG-63、U20S细胞中miR-495的相对表达量低于hFOB1.19细胞(P均<0.01)。
2.2 miR-495对MG-63细胞增殖的影响 转染后48、72、96 h后miR-495 mimics组MG-63细胞OD值低于阴性对照组,miR-495 mimics+HMGA2组MG-63细胞OD值高于miR-495 mimics组(P均<0.01)。详见表1。
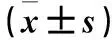
表1 各组细胞不同时间OD值比较
2.3 miR-495的靶基因及结合位点 HMGA2的3′UTR区存在与miR-495的种子序列理论上互补的结合位点,HMGA2可能是miR-495的直接调控靶基因。双荧光素酶报告实验结果显示,HMGA2野生型质粒实验组、HMGA2野生型质粒对照组、HMGA2突变型质粒实验组和HMGA2突变型质粒对照组细胞荧光素酶活性分别为0.31±0.09、1.01±0.03、0.98±0.04、1.02±0.04,HMGA2野生型质粒实验组细胞荧光素酶活性低于HMGA2野生型质粒对照组(P<0.01),而HMGA2突变型质粒实验组与HMGA2突变型质粒对照组的细胞荧光素酶活性差异无统计学意义(P>0.05)。这证实miR-495能够特异性结合HMGA2的3′UTR区,使荧光蛋白表达水平明显降低,而HMGA2的3′UTR区突变后则无此作用,提示miR-495可与HMGA2的3′UTR区特异性结合。
2.4 miR-495表达变化对MG-63细胞中HMGA2蛋白表达的影响 miR-495 mimics组、miR-495 mimics+HMGA2组和阴性对照组细胞中HMGA2蛋白相对表达量分别为0.23±0.10、0.94±0.06、1.01±0.04。miR-495 mimics组MG-63细胞中HMGA2蛋白的相对表达量低于阴性对照组,miR-495 mimics+HMGA2组MG-63细胞中HMGA2蛋白的相对表达量高于miR-495 mimics组(P均<0.01)。
3 讨论
近年来随着表观遗传学的发展,miRNA在疾病中作用越来越受到重视。研究显示,在多数恶性肿瘤中存在miRNA的异常表达,miRNA在恶性肿瘤的发生发展过程中发挥着重要作用[9]。miR-495基因定位于染色体14q32.31,属于近期发现的miRNA重要家族成员之一,其在机体细胞增殖、凋亡及机体免疫、炎症反应方面起着重要的作用。研究发现miR-495表达异常与肿瘤细胞的增殖、凋亡及肿瘤细胞化疗耐药关系密切。miR-495在大部分实体肿瘤中发挥着抑癌基因的作用,但在肝细胞癌、乳腺癌等少数实体肿瘤中表达升高,起着癌基因的作用[10]。本研究中检测了骨肉瘤细胞Saos-2、U20S、MG-63及人成骨细胞hFOB1.19中的miR-495,发现miR-495在骨肉瘤细胞中表达均明显下调,提示miR-495在骨肉瘤中可能起着抑癌基因的作用。为了研究miR-495对骨肉瘤细胞增殖的影响,我们通过脂质体介导法将miR-495 mimics成功转染入骨肉瘤MG-63细胞,CCK-8实验显示上调miR-495能够明显抑制MG-63细胞的增殖,证实了上述推论。
目前miR-495对恶性肿瘤调控的具体分子机制尚不明确。为了研究miR-495调控骨肉瘤细胞增殖的机制,我们应用靶基因预测数据库miRanda,并结合基因的功能分析,预测到HMGA2为miR-495的可能靶基因,并通过双荧光素酶报告实验证实miR-495能够特异性的结合HMGA2的3′UTR区,使荧光蛋白活性明显降低。Western blotting检测HMGA2蛋白,结果显示上调miR-495表达后骨肉瘤细胞中HMGA2蛋白表达量明显降低,充分证实了miR-495对HMGA2存在靶向负调控作用,HMGA2是miR-495直接调控的靶基因之一。
HMGA2是一种非组蛋白染色体蛋白,是高迁移率族蛋白超家族成员之一。HMGA2基因被认为是一种新的癌基因,在甲状腺癌[11]、卵巢癌[12]、前列腺癌[13]、胆囊腺癌[14]、食管癌[15]、膀胱癌[16]、非小细胞肺癌[17]和胃癌[18]等实体肿瘤组织中高表达。HMGA2可上调细胞周期蛋白的表达,加速G2/M期转化,促使肿瘤发生、发展[19]。Morishita等[20]发现HMGA2可激活TGF-β信号通路促进EMT,增强肿瘤细胞的侵袭和转移能力。目前研究已经证实下调骨肉瘤细胞中HMGA2的表达能够明显抑制骨肉瘤细胞的增殖[21]。本研究结果显示miR-495 mimics+HMGA2组MG-63细胞中HMGA2蛋白相对表达量高于miR-495 mimics组,成功逆转了miR-495对HMGA2蛋白的抑制作用,并且进一步通过CCK-8实验证实转染HMGA2表达质粒能够明显逆转miR-495对MG-63细胞增殖的抑制作用,提示miR-495可能通过调控HMGA2的表达从而抑制骨肉瘤细胞的增殖能力。
通过上述实验,我们发现骨肉瘤细胞中miR-495的表达水平低于正常成骨细胞,上调miR-495表达后骨肉瘤细胞增殖受到抑制,且miR-495可能通过靶向调控HMGA2表达从而抑制骨肉瘤细胞的增殖。
