吡格列酮对新生大鼠成骨细胞p38和ERK1/2信号通路的影响
2018-07-05刘永立程富礼景小博王来喜
刘永立,程富礼,景小博,王来喜
0 引言
骨质疏松症是常见的人代谢骨疾病,其特征在于进行性和年龄依赖性骨丢失和骨折风险增加。近年来,因药物引起的骨质疏松症得到广泛关注。噻唑烷二酮类(TZDs,曲格列酮、罗格列酮和吡格列酮)是PPARγ的人工合成配体,是用于治疗糖尿病的胰岛素敏化化合物。已有研究表明,TZDs在骨代谢过程中发挥重要作用。临床研究表明,曲格列酮治疗能够降低2型糖尿病患者的骨转换率[1]。A糖尿病结局进行试验(ADOPT)表明,罗格列酮增加了2型糖尿病绝经后妇女骨折的风险[2]。另外,已有研究表明了TZDs对骨生物标志物的影响。Gray等[3]发现,吡格列酮治疗能够降低碱性磷酸酶(ALP)的活性。Berberoglu等[4]发现,罗格列酮治疗降低了ALP和骨钙素的生成。这些研究表明,TZDs对骨骼存在不利作用,存在引起骨质疏松的风险。然而,吡格列酮对骨代谢的影响及相关机制尚不完全清楚。本研究通过体外分离培养新生大鼠颅骨成骨细胞,探究吡格列酮对成骨细胞增殖、凋亡和分化的影响,并初步探讨其可能的机制,以期对TZDs的临床用药提供一定参考。
1 材料与方法
1.1 材料 新生24 h内SD大鼠幼仔(北京生命科学研究所,许可证号:SYXK2015-0003),均为SPF级动物,雌雄及体重不限。吡格列酮购自上海迈瑞尔化学技术有限公司(纯度99%,CAS号:112529-15-4);3-(4,5-二甲基噻唑-2-基)-2,5-二苯基四唑溴化物(MTT)、4-(2-羟乙基)-1-哌嗪乙磺酸(HEPES)、蛋白酶抑制剂混合物购自美国Calbiochem公司;ALP比色测定试剂盒购自赛奥生物科技有限公司;DMEM培养基、胎牛血清(FBS)、青霉素/链霉素购自美国GIBCO公司;抗Bcl-2、Bax、磷酸化(p-)p38、p-细胞外信号调节激酶(ERK)1/2、p-C-Jun N-末端激酶(JNK)、BMP-2、Runx2、OSX抗体购自美国Cell Signaling Technology公司;反转录试剂盒购自日本Takara公司;引物购自上海生工。
1.2 大鼠颅骨成骨细胞的分离、培养 取出生24 h内的SD大鼠8只,处死后置于75%酒精中,消毒3~5 min后,于无菌条件下取出颅骨,除骨膜及结缔组织后,PBS清洗3次。将颅骨剪成1 mm2左右的碎块,用0.25%胰蛋白酶2 ml,37 ℃预消化10 min,用血清培养液10 ml终止消化后,1 000 r/min离心5 min,弃上清。将沉淀物于0.1%Ⅱ型胶原酶10 ml,37 ℃消化100 min后终止消化。收集消化液,1 000 r/min离心5 min,弃上清,取细胞沉淀,PBS漂洗后,重复离心过程。于离心管中加入含10%胎牛血清、100 U/ml青霉素、100 mg/L链霉素的DMEM培养液2 ml,将沉淀细胞吹打均匀,并于200目滤网过滤去除碎骨片,制成细胞悬浊液。将细胞悬浊液接种至细胞培养瓶中静置10 min后,吸收上清液并置于另一培养瓶中,最后将细胞悬浊液接种于培养皿中,于37 ℃、5% CO2、饱和湿度培养,隔日换液。待显微镜下观察可见成骨细胞80%以上融合时,用0.25%胰蛋白酶消化30 s后,进行细胞传代培养。
1.3 成骨细胞分化及鉴定 第3代成骨细胞培养至出现80%融合时,将细胞以5×103个/cm2接种在40 mm培养皿中,并在成骨分化培养基(含10%胎牛血清、10 mmol/L HEPES、50 μg/ml L-抗坏血酸和5 mmol/L β-甘油磷酸酯的DMEM培养基)中培养21 d,每周更换2次培养基。随后用PBS洗涤细胞2次,用4%福尔马林固定15 min,并用水洗涤,然后将细胞与茜素红孵育2 min进行染色。PBS洗涤3次后,用蒸馏水洗涤数次。于镜下观察细胞。
1.4 成骨细胞增殖、凋亡测定 细胞增殖测定:第3代成骨细胞培养至出现80%融合时,将细胞以5×103个/孔接种在96孔培养板中孵育过夜。加入含0、10、20、40 μmol/L吡格列酮的DMEM培养基培养48 h后,每孔加入20 μl MTT,孵育4 h后,弃去上清液,于每孔中加入100 μl二甲基亚砜,振荡混合3 min后,使用酶标仪于490 nm下测量吸光度(OD490)。
细胞凋亡测定:应用Hoechst 33258染色检测细胞凋亡。第3代细胞培养至出现80%融合时,将细胞以4×104个/cm2接种至12孔板中,孵育过夜。根据参考文献[5],于培养基中加入含0、10、20、40 μmol/L吡格列酮的DMEM培养基培养48 h后,PBS洗涤细胞以除去非贴壁细胞,贴壁细胞置于4%多聚甲醛中固定10 min。PBS洗涤后,细胞与0.2 mmol/L Hoechst 33258在黑暗中孵育5 min。于荧光显微镜下观察凋亡成骨细胞数。
1.5 ALP活性检测 第3代成骨细胞培养至出现80%融合时,将细胞以5×103个/cm2接种在40 mm培养皿中,加入含0、10、20、40 μmol/L吡格列酮的DMEM培养基培养48 h,更换为成骨分化培养基培养,每周更换2次培养基。各组细胞培养的第14天,弃培养液,用PBS洗涤细胞2次后,收集细胞,1 000 r/min离心30 min。取上清液,使用ALP活性测定试剂盒检测ALP活性,所有操作均严格按照说明书进行。在酶标仪上于420 nm处读取吸光值(OD420)。
1.6 钙化结节染色鉴定 应用茜素红染色进行钙化结节测定。“1.6”项下各组细胞培养的第21天,弃培养液,用PBS洗涤细胞2次后,用4%福尔马林固定15 min。用水洗涤后,将细胞与0.1%茜素红(pH 8.9)于37 ℃下孵育2 min。PBS洗涤3次后,用蒸馏水洗涤数次后,拍照记录钙化结节情况。
1.7 实时荧光定量PCR(qRT-PCR) 应用qRT-PCR进行成骨分化相关基因的mRNA表达检测。将细胞数调节至1×106/ml,TRIzol试剂(Invitrogen,美国)从细胞提取总RNA,Reverse Transcription Kit(Takara,日本)将RNA逆转录为cDNA,SYBR Green(Takara)进行实时PCR分析。以β-Actin作为内参。利用ABI7500系统(Applied Biosystems)进行qRT-PCR反应。用2-ΔΔCt法计算mRNA相对表达的倍数变化。qRT-PCR反应条件:95 ℃ 30 s,95 ℃15 s,72 ℃ 15 s,共40个循环。引物设计及合成由上海生工完成,引物序列如下:BMP-2,forward 5′-GGTGGAATGACTGGATTG-3′,reverse 5′-GCATCGAGATAGCACTG-3′;Runx2,forward 5′-GGAATTCCATATGGACACTT-3′,reverse 5′-CCGCTCGAGGTTATAGTCCT-3′;OSX,forward 5′-TGTCCGTCGTGGATCTGAC-3′,reverse 5′-CCTGCTTCACCACCTTCTTG-3′;β-actin,forward 5′-CTCCTGGCAAGAATGGAGAT-3′,reverse 5′-AATCCACGA
GCACCCTGA-3′。
1.8 蛋白免疫印迹(Western blot,WB) 使用WB进行蛋白表达检测,补充有PMSF(Riche)的RIPA蛋白提取试剂(Beyotime)裂解细胞,从细胞中提取蛋白质。在4 ℃下12 000 r/min离心10 min,通过10%十二烷基硫酸钠聚丙烯酰胺凝胶电泳(SDS-PAGE)分离上清液,将分离的蛋白质转移到硝酸纤维素膜(Sigma)上。将膜在PBS和5%脱脂乳中封闭1 h,并用抗Bcl-2、Bax、p-p38、p-ERK1/2、p-JNK、BMP-2、Runx2、OSX的一抗于4 ℃孵育过夜。将膜用HRP缀合的二抗(Cell Signaling Technology)在室温下孵育1 h。使用Quantity One软件(Life Technologies)评估相对蛋白表达水平。
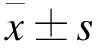
2 结果
2.1 成骨细胞形态观察及鉴定 倒置显微镜下观察发现,大鼠成骨细胞原代培养的第2天,成骨细胞呈圆形,单细胞离散分布(图1A)。在培养的第4天,细胞数量增加,细胞形态多样,呈多角形、三角形、梭形等(图1B)。细胞培养的第14天,细胞迅速增殖,细胞呈梭形或立方形,排列紧密,可多层生长,细胞整合达90%,可进行传代培养(图1C)。第3代成骨细胞培养21 d后,细胞重叠生长,茜素红染色显示,胞浆内可见棕黑色或黑色钙结节沉积(图1D)。提示本研究所培养新生大鼠颅骨成骨细胞具有细胞分化能力。
2.2 吡格列酮对成骨细胞增殖的影响 第3代成骨细胞经0、10、20、40 μmol/L吡格列酮处理48 h后,细胞活性(OD490)分别为0.82±0.03、0.58±0.06、0.38±0.04、0.25±0.03,随着吡格列酮浓度的增加,各组细胞活性逐渐降低,差异有统计学意义(F=282.629,P<0.01)。见图2。
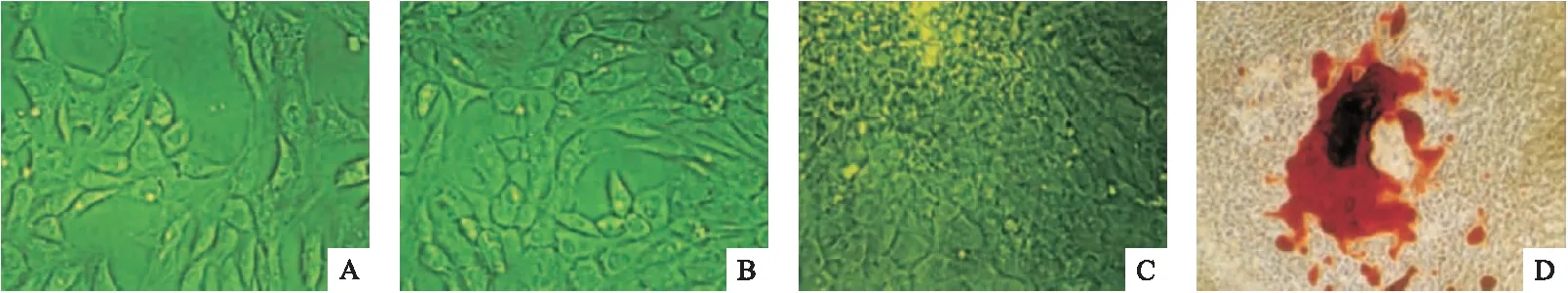
图1 成骨细胞形态观察及鉴定
注:*与空白对照组比较,P<0.05;#与10 μmol/L组比较,P<0.05;△与20 μmol/L组比较,P<0.05
2.3 吡格列酮对成骨细胞凋亡的影响 第3代成骨细胞经0、10、20、40 μmol/L吡格列酮处理48 h后,荧光显微镜下观察成骨细胞Hoechst 33258染色阳性细胞数。与空白对照组相比,经10、20 μmol/L吡格列酮处理组荧光阳性成骨细胞数量显著降低,差异有统计学意义(P<0.05)。与空白对照组及吡格列酮10 μmol/L组、20 μmol/L组相比,吡格列酮40 μmol/L组荧光阳性成骨细胞数量显著增加,差异有统计学意义(P<0.05)。提示低浓度(10、20 μmol/L)吡格列酮抑制成骨细胞凋亡,而高浓度(40 μmol/L)吡格列酮促进成骨细胞凋亡。见图3。

图3 吡格列酮对成骨细胞凋亡的影响(n=8)
2.4 吡格列酮对成骨细胞Bax、Bcl-2蛋白表达的影响 与空白对照组相比,第3代成骨细胞经10、20 μmol/L吡格列酮处理48 h后,细胞内Bax蛋白表达水平显著降低(P<0.05),而Bcl-2蛋白表达水平显著增加(P<0.05)。当吡格列酮浓度为40 μmol/L时,与空白对照组相比,成骨细胞内Bax蛋白表达水平显著增加(P<0.05),而Bcl-2蛋白表达水平显著降低(P<0.05)。见图4。

图4 吡格列酮对成骨细胞Bax、Bcl-2蛋白表达的影响(n=8)
注:*与空白对照组比较,P<0.05;#与10 μmol/L组比较,P<0.05;△与20 μmol/L组比较,P<0.05
2.5 吡格列酮对成骨细胞分化的影响
2.5.1 吡格列酮对ALP活性的影响 第3代成骨细胞经0、10、20、40 μmol/L吡格列酮处理48 h后,ALP活性(OD420)分别为1.07±0.15、0.89±0.04、0.75±0.03、0.53±0.03,随着吡格列酮浓度的增加,各组细胞ALP活性逐渐降低,差异有统计学意义(F=64.247,P<0.01)。见图5。
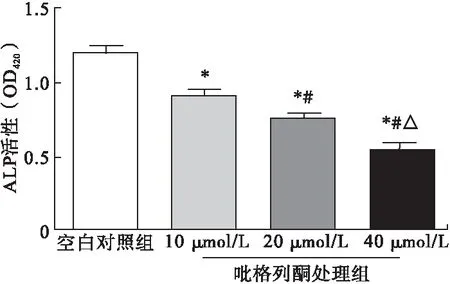
图5 吡格列酮对成骨细胞ALP活性的影响(n=8)
注:*与空白对照组比较,P<0.05;#与10 μmol/L组比较,P<0.05;△与20 μmol/L组比较,P<0.05
2.5.2 吡格列酮对成骨细胞钙化结节形成的影响 与空白对照组相比,吡格列酮处理组的钙化结节数均明显降低(图6A)。以钙化结节在培养皿中所占面积的百分比进行量化,发现随着吡格列酮浓度的增加,钙化结节数量逐渐增加,且差异均有统计学意义(F=672.644,P<0.01)。见图6B。
2.5.3 吡格列酮对成骨细胞BMP-2、Runx2、OSX mRNA表达的影响 经0、10、20、40 μmol/L吡格列酮处理的成骨细胞于成骨分化培养基中培养48 h后,应用qRT-PCR检测吡格列酮对成骨细胞分化相关基因表达的影响。与空白对照组相比,经10、20、40 μmol/L吡格列酮处理后,细胞内BMP-2、Runx2、OSX mRNA表达水平均以剂量依赖形式显著降低(P<0.05)。见图7。

图6 吡格列酮对成骨细胞钙化结节形成的影响(n=8)
注:A.茜素红染色观察到钙化结节,B.钙化结节在培养皿中所占面积百分比。*与空白对照组比较,P<0.05;#与10 μmol/L组比较,P<0.05;△与20 μmol/L组比较,P<0.05

图7 吡格列酮对成骨细胞BMP-2、Runx2、OSX mRNA表达的影响(n=8)
注:*与空白对照组比较,P<0.05;#与10 μmol/L组比较,P<0.05;△与20 μmol/L组比较,P<0.05
2.5.4 吡格列酮对成骨细胞BMP-2、Runx2、OSX蛋白表达的影响 与空白对照组相比,经10、20、40 μmol/L吡格列酮处理后,细胞内BMP-2、Runx2、OSX蛋白表达水平均以剂量依赖形式显著降低,差异均有统计学意义(P<0.05)。见图8。
2.6 吡格列酮对成骨细胞MAPK信号通路的影响 经0、10、20、40 μmol/L吡格列酮处理的成骨细胞于成骨分化培养基中培养48 h后,应用免疫印迹分析,检测吡格列酮对成骨细胞MAPK信号通路的影响,结果发现,与空白对照组比较,经10、20、40 μmol/L吡格列酮处理后,细胞内p-p38 MAPK蛋白表达水平均显著增加(P<0.05),而p-ERK1/2蛋白表达水平显著降低(P<0.05),且均存在剂量依赖性。各组细胞内p-JNK蛋白表达水平差异无统计学意义(P<0.05)。见图9。
图8 吡格列酮对成骨细胞BMP-2、Runx2、OSX蛋白
注:*与空白对照组比较,P<0.05;#与10 μmol/L组比较,P<0.05;△与20 μmol/L组比较,P<0.05

图9 吡格列酮对成骨细胞MAPK信号通路的影响(n=8)
注:*与空白对照组比较,P<0.05;#与10 μmol/L组比较,P<0.05;△与20 μmol/L组比较,P<0.05
3 讨论
本研究采用新生24 h内SD大鼠颅骨组织来源的成骨细胞进行培养,倒置显微镜下观察到成骨细胞生长状态良好,形态与过往学者报道一致。通过茜红素染色可使钙化结节呈现棕黑色或黑色沉积,证实所培养成骨细胞具有分化为钙结节的能力。
研究表明,TZDs对骨代谢过程存在不利作用,能够抑制骨骼间充质干细胞向成骨细胞分化,抑制骨形成,诱导成骨细胞凋亡,使成骨细胞正常功能受损,导致骨量减少,并增加骨折风险[6-7]。研究证实,吡格列酮能够抑制成骨细胞增殖、分裂和DNA合成,且低浓度吡格列酮(≤20 μmol/L)能够抑制成骨细胞凋亡,而较高浓度吡格列酮(30~40 μmol/L)则促进成骨细胞凋亡[5]。与此一致,本研究发现,与空白对照组相比,经10、20、40 μmol/L吡格列酮处理后,成骨细胞活性下降。另外,与空白对照组相比,吡格列酮浓度为10、20 μmol/L时,成骨细胞凋亡被抑制,而当吡格列酮浓度为40 μmol/L时,能够促进成骨细胞凋亡。Bcl-2家族蛋白是调节线粒体损伤的关键分子,Bax是Bcl-2家族中重要的促凋亡蛋白,在细胞凋亡调控中起重要作用。研究证实,Bax表达的增加、Bcl-2表达水平的降低能够导致细胞凋亡[8-9]。研究发现,罗格列酮能够剂量依赖性促进Bax表达增加,从而导致成骨细胞死亡[10]。本研究发现,当吡格列酮浓度为10、20 μmol/L时,成骨细胞内Bax蛋白表达水平降低,而Bcl-2蛋白表达水平显著增加。当吡格列酮浓度为40 μmol/L时,成骨细胞内Bax蛋白表达水平增加,而Bcl-2蛋白表达水平显著降低。提示吡格列酮可通过调节Bcl-2和Bax蛋白表达水平,调控成骨细胞凋亡,且这种调控作用存在剂量依赖性,其机制可能在于成骨细胞自身调节机制可以对抗低浓度吡格列酮诱导的凋亡作用,而随着吡格列酮含量的增加,Bcl-2/Bax平衡超过自身调节范围,无法对抗药物刺激引起的细胞凋亡。
成骨细胞增殖抑制、凋亡增加及分化程度降低,是造成骨质疏松症的重要原因。TZDs对成骨细胞分化影响的研究较少,且吡格列酮对成骨细胞分化的影响及相关机制尚未报道。本研究进一步分析吡格列酮对大鼠成骨细胞分化功能的影响,并探讨潜在机制。
ALP是成骨细胞在分化早期所分泌的酶蛋白,是骨形成的标志性蛋白,特异性较高,可作为鉴定成骨细胞的生化和组织学标志[11]。ALP变化反映了骨转换过程中骨形成程度,其活性与成骨细胞的分化程度呈正相关。有证据表明,ALP活性的微小降低直接影响成骨细胞矿化形成过程。另外,成骨细胞分化的标志性表型是矿化结节的形成。本研究发现,经不同浓度吡格列酮处理后,各组成骨细胞ALP活性降低,且吡格列酮可抑制成骨细胞矿化结节的形成,提示吡格列酮可抑制大鼠成骨细胞的分化,且随着其浓度的增加,抑制作用逐渐增强。BMP-2是促进骨形成、诱导成骨细胞分化的重要细胞外信号分子之一,是最强的成骨因子之一。BMP-2调节因子Runx2是间质细胞分化成前成骨细胞所必需的[12]。Runx2下游的OSX是成骨细胞特异性的成骨细胞分化和骨形成所必需的转录因子[13]。本研究发现,经吡格列酮处理后,成骨细胞内BMP-2、Runx2、OSX mRNA和蛋白表达以剂量依赖的形式降低,由此推测,吡格列酮通过调控成骨细胞内BMP-2/Runx2/OSX信号通路,参与调控大鼠成骨细胞分化功能。
丝裂原活化蛋白激酶(MAPKs)是能够被不同细胞外刺激激活的丝氨酸-苏氨酸蛋白激酶,是信号转导过程中的主要信号分子,参与多种重要的细胞生理病理过程[14-15]。已在哺乳动物细胞中鉴定了MAPKs的3条并行信号通路:p38、细胞外信号调节激酶(ERK)和C-Jun N-末端激酶(JNK),在细胞增殖、分化、凋亡等过程中都发挥重要作用。研究表明,包括成骨细胞在内的多种细胞类型中,ERK参与细胞的增殖、分化和存活,而p38和JNK参与细胞周期控制、细胞凋亡和炎症反应[16]。Tang等[17]发现,BMP-2在成骨细胞中的表达受p38和ERK信号通路调控。ERK1/2组成型活性形式的激活能够诱导成骨细胞的分化[18-19]。p38的磷酸化能诱导smad1活化,从而调控BMP-2/smads信号通路,导致成骨细胞ALP表达和活化[20],且BMP-2在成骨细胞中的表达受MAPK调控[21-23]。另外,在成骨细胞中,JNK和p-38 MAPK能够诱导细胞凋亡,而ERK能够抑制凋亡过程[24]。这些研究表明,MAPK信号通路参与成骨细胞特异性基因表达和功能调控。已有研究报道,吡格列酮通过调控MAPK信号通路,调节细胞炎症反应[25]。体外研究表明,曲格列酮通过激活MAPK信号通路增加成骨细胞的凋亡[26]。然而,吡格列酮在成骨细胞中对MAPK信号通路的影响尚不明确。本研究发现,经吡格列酮处理后,成骨细胞内p-p38蛋白表达水平增加,而p-ERK1/2蛋白表达水平降低,且均呈剂量依赖性。而p-JUN1/2蛋白表达水平无明显变化。推测吡格列酮能够通过活化p38信号通路和抑制ERK1/2信号途径参与成骨细胞凋亡和分化过程。成骨细胞代谢过程十分复杂,作为细胞内信号转导的主要信号分子,MAPK信号通路在吡格列酮影响成骨细胞增殖、分化及凋亡过程中的作用还需深入探究,以期为临床用药提供一定参考依据。
综上所述,吡格列酮以剂量依赖的方式调节大鼠成骨细胞增殖、凋亡和分化,p38和ERK1/2信号途径在这一过程中起重要调控作用。
参考文献:
[1] Okazaki R,Miura M,Toriumi M,et al.Short-term treatment with troglitazone decreases bone turnover in patients with type 2 diabetes mellitus[J].Endocr J,1999,46(6):795-801.
[2] Pfützner A,Forst T.A diabetes outcome and progression trial-die ADOPT studie[J].Der Diabetologe,2007,3(4):261-269.
[3] Grey A,Bolland M,Gamble G,et al.The peroxisome proliferator-activated receptor-gamma agonist rosiglitazone decreases bone formation and bone mineral density in healthy postmenopausal women:a randomized,controlled trial[J].J Clin Endocrinol Metab,2007,92(4):1305-1310.
[4] Berberoglu Z,Gursoy A,Bayraktar N,et al.Rosiglitazone decreases serum bone-specific alkaline phosphatase activity in postmenopausal diabetic women[J].J Clin Endocrinol Metab,2007,92(9):3523-3530.
[5] 李俊岩,董进,牛晓红,等.吡格列酮对成骨细胞Bax、Bcl-2蛋白表达的影响[J].中华内分泌代谢杂志,2011,27(2):118-121.
[6] 李丽华.吡格列酮对大鼠骨髓间充质干细胞向成骨细胞分化后转脂分化的影响[D].华中科技大学,2009.
[7] 侯艳丽,季虹,荣海钦.噻唑烷二酮类药物对骨代谢的影响[J].中华骨质疏松和骨矿盐疾病杂志,2009,2(3):189-193.
[8] Kokawa K,Shikone T,Otani T,et al.Apoptosis and the expression of Bax and Bcl-2 in squamous cell carcinoma and adenocarcinoma of the uterine cervix[J].Cancer,1999,85(8):1799-1809.
[9] Zhang T,Tian F,Wang J,et al.Atherosclerosis-associated endothelial cell apoptosis by MiR-429-mediated down regulation of Bcl-2[J].Cell Physiol Biochem,2015,37(4):1421-1430.
[10]Hsiao FY,Mullins CD.The association between thiazolidinediones and hospitalisation for fracture in type 2 diabetic patients:a Taiwanese population-based nested case-control study[J].Diabetologia,2010,53(3):489-496.
[11]Suzuki A,Takayama T,Suzuki N,et al.Daily low-intensity pulsed ultrasound-mediated osteogenic differentiation in rat osteoblasts[J].Acta Biochim Biophys Sin (Shanghai),2009,41(2):108-115.
[12]Guo CH,Yue ZP,Bai ZK,et al.Runx2 acts downstream of C/EBPβ to regulate the differentiation of uterine stromal cells in mice[J].Cell Tissue Res,2016,366(2):393-401.
[13]Wang L,Mishina Y,Liu F.Osterix-Cre transgene causes craniofacial bone development defect[J].Calcif Tissue Int,2015,96(2):129-137.
[14]高黎,赵志英.MAPK信号转导通路及其意义[J].世界最新医学信息文摘(连续型电子期刊),2015,15(53):25-27.
[15]安秋霞,蒙艳丽,吕丹丹,等.丝裂原活化蛋白激酶信号通路的研究进展[J].黑龙江中医药,2016,45(5):65-66.
[16]Hu Y,Chan E,Wang SX,et al.Activation of p38 mitogen-activated protein kinase is required for osteoblast differentiation[J].Endocrinology,2003,144(5):2068-2074.
[17]Tang CH,Yang RS,Chien MY,et al.Enhancement of bone morphogenetic protein-2 expression and bone formation by coumarin derivatives via p38 and ERK-dependent pathway in osteoblasts[J].Eur J Pharmacol,2008,579(1-3):40-49.
[18]Ge C,Xiao G,Jiang D,et al.Critical role of the extracellular signal-regulated kinase-MAPK pathway in osteoblast differentiation and skeletal development[J].J Cell Biol,2007,176(5):709-718.
[19]廖清船,徐康康,许静,等.p38MAPK与ERK1/2的协同效应及对成骨细胞分化的调控[J].中华骨科杂志,2011,31(9):970-975.
[20]Nöth U,Tuli R,Seghatoleslami R,et al.Activation of p38 and Smads mediates BMP-2 effects on human trabecular bone-derived osteoblasts[J].Exp Cell Res,2003,291(1):201-211.
[21]Tsai HY,Lin HY,Fong YC,et al.Paeonol inhibits RANKL-induced osteoclastogenesis by inhibiting ERK,p38 and NF-kappaB pathway[J].Eur J Pharmacol,2008,588(1):124-133.
[22]赵良功.MAPK家族在成骨细胞分化过程中调控作用的研究[D].兰州大学,2015.
[23]陈学鹏,施洁珺,叶青松,等.p38MAPK信号通路在BMP-2诱导人牙囊细胞成骨分化中的作用[J].中国细胞生物学学报,2013,35(6):816-823.
[24]Guo C,Yang XG,Wang F,et al.IL-1α induces apoptosis and inhibits the osteoblast differentiation of MC3T3-E1 cells through the JNK and p38 MAPK pathways[J].Int J Mol Med,2016,38(1):319-327.
[25]汪姗,叶山东,孙文佳,等.吡格列酮对肾小球系膜细胞ROS和p38MAPK的影响[J].中国药理学通报,2014,30(1):82-85.
[26]Jung JY,Yoo CI,Kim HT,et al.Role of mitogen-activated protein kinase (MAPK) in troglitazone-induced osteoblastic cell death[J].Toxicology,2007,234(1-2):73-82.
