内蒙古羊屠宰场致泻性大肠杆菌和沙门氏菌污染状况分析
2018-06-05杨永青姚一萍狄彩霞李秀萍栗艳芳
杨永青,姚一萍,史 培,赵 格,狄彩霞,莎 娜,李秀萍,栗艳芳,骆 洪
(1.内蒙古自治区农牧业科学院,农业部农产品质量安全风险评估实验室(呼和浩特),内蒙古呼和浩特 010031;2. 内蒙古大学生命科学学院,内蒙古呼和浩特 010021;3.中国动物卫生与流行病学中心,农业部畜禽产品质量安全风险评估实验室(青岛),山东青岛 266032)
以上的沙门氏菌感染都由动物性食品引起[4-5]。现有报道多集中于鸡、鸭和生猪屠宰环节食源性致病菌的污染种类、水平、耐药性等,而针对不同规模羊屠宰场中各屠宰环节的相关研究较少,而研究屠宰环节羊肉产品沙门氏菌和致病性大肠埃希氏菌的携带状况,比较不同规模屠宰场的携带水平,能够为羊肉产品食源性病原微生物的防控提供基础数据。
1 材料与方法
1.1 材料
1.1.1 样品来源 羊肉胴体/屠宰环境拭子:用一次性无菌棉签在羊肉、环境表面(>30 cm2)擦拭,然后将其放入运输培养基中,36 h内冷藏运送至实验室。同时采集2份棉拭子,分别用于分离致泻性大肠杆菌和沙门氏菌。2017年4—11月,选取内蒙古6个不同规模屠宰场(日屠宰量>200只为大中型屠宰场,日屠宰量<200只为小型屠宰场),在各屠宰环节(刚宰杀、预冷间和屠宰环境)采样(图1),共采集349份棉拭子样品。其中,屠宰环境包括地面、墙面、工作人员手套、台面和托盘。
1.1.2 主要试剂 一次性无菌棉拭子、运输培养基、营养肉汤、肠道增菌肉汤、麦康凯培养基、伊红美蓝培养基、缓冲蛋白胨水(BPW)、四硫磺酸钠煌绿增菌液(TTB)、沙门氏菌显色培养基和革兰氏阴性杆菌鉴定系统(HK-MID-64,GN-ID A):购自广东环凯微生物科技有限公司;DBI-05沙门氏菌干制生化鉴定试剂盒:购自北京陆桥技术有限责任公司。其他试剂包括细菌基因组提取试剂盒、GoTaq Green Master Mix 、DNA Marker和ddH2O等。
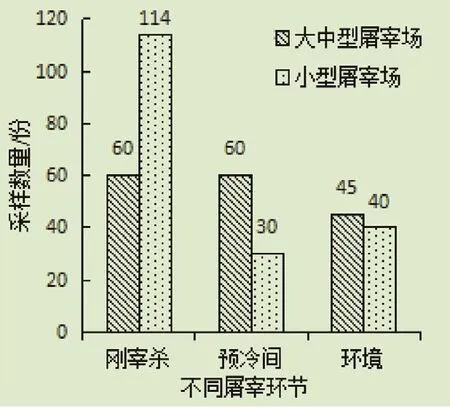
图1 不同规模屠宰场各屠宰环节采样数量
1.2 方法
1.2.1 致泻性大肠杆菌分离与生化鉴定 致泻性大肠杆菌的富集和分离参照《食品安全国家标准食品卫生微生物学检验致泻大肠埃希氏菌检验》(GB/T 4789.6—2016)[6]进行。从麦康凯琼脂(MAC)上挑取表面光滑的红色或桃红色单菌落,划线接种于营养琼脂平板纯培养[7]。挑取少量纯化后的单菌落,用无菌水制成0.5麦氏浊度的菌悬液,用HK-MID-64,GN-ID A鉴定系统进行生化鉴定。大肠埃希氏菌(Escherichia coli)ATCC 25922为质控菌株。
1.2.2 沙门氏菌分离与生化鉴定 沙门氏菌的富集和分离参照《食品安全国家标准食品卫生微生物学检验沙门氏菌检验》(GB/T 4789.4—2016)[8]方法进行。从沙门氏菌显色培养基上挑取紫色或淡紫色的单菌落,接种胰蛋白大豆胨平板纯培养。挑取少量纯化后的单菌落,用无菌水制成0.5麦氏浊度的菌悬液,用DBI-05沙门氏菌干制生化鉴定试剂盒进行生化鉴定。
1.2.3 病原菌毒力基因PCR扩增 将符合生化现象的分离株接种于营养肉汤中,37 ℃培养12~14 h,按照细菌基因组提取试剂盒说明书提取DNA。分别扩增致泻性大肠杆菌的6种毒力基因、沙门氏菌invA基因。特异性引物(表1)由上海生工生物工程技术有限公司合成。致泻性大肠杆菌多重PCR[9]、沙门氏菌PCR反应体系(25 µL):GoTaq Green Master Mix 12.5 µL,Primer Mix 1 µL,DNA模板1 µL,ddH2O 10.5 µL。PCR扩增条件见表2。PCR产物经2.0%琼脂糖凝胶电泳。

表1 病原菌毒力基因特异性引物序列
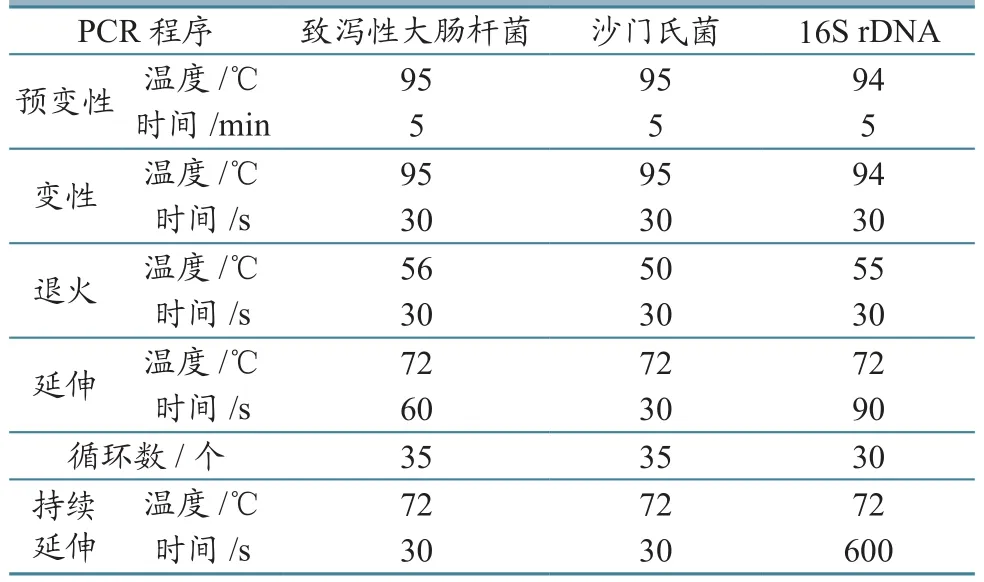
表2 致泻性大肠杆菌和沙门氏菌PCR扩增程序
1.2.4 致泻性大肠杆菌16S rDNA鉴定及系统发育树构建 以携带毒力基因的致泻性大肠杆菌DNA为模板,扩增16S rDNA序列。PCR扩增条件[10]见表2。细菌16S rDNA通用引物:27 F:5´AGAGTTTGATCCTGGCTCAG 3´;1492 R:5´TACGGTTACCTTGTTACGACTT3´。PCR 体 系(50 µL):GoTaq Master Mix 25.0 µL,27 F 1 µL,1492 R 1 µL,DNA 1 µL,ddH2O 22.0 µL。PCR产物委托上海生工生物测序。16S rDNA序列用MEGA5.0软件,以Neighbor-Joining法构建系统发育树。
2 结果
2.1 致泻性大肠杆菌毒力基因PCR扩增
经生化鉴定筛选出101株符合大肠杆菌生化反应结果的分离株,多重PCR扩增其毒力基因并由电泳结果(图2)得出,携带致泻性大肠杆菌毒力基因的共12株,其中1株(36号)携带毒力基因eae、elt,其余11株携带毒力基因elt,均属于能够引起动物肠道疾病的产肠毒素大肠埃希氏菌(ETEC)。

图2 致泻性大肠杆菌毒力基因多重PCR产物电泳图
2.2 沙门氏菌毒力基因PCR扩增
经生化鉴定筛选出5株符合沙门氏菌生化现象的分离株。PCR扩增其invA基因电泳结果见图3。编号为149的样品出现与阳性对照大小一致的目的片段,是携带invA基因的沙门氏菌。

图3 沙门氏菌invA基因PCR产物电泳图
2.3 致泻性大肠杆菌16S rDNA鉴定及系统发育树
扩增12株致泻性大肠杆菌的16S rDNA,均得到大小约为1 500 bp的单一条带,与目的片段大小一致,无明显非特异性扩增现象。部分阳性菌株PCR产物电泳结果见图4。由16S rDNA序列构建系统发育树(图5)得出,5株阳性菌株18、26、31、36、132与Genebank中鸡源(EF620922.1)、鸭 源(EF620921.1) 和 羊 源(EF025911.1、EF025907.1)等动物源大肠杆菌处于同一分支,亲缘关系较近,其余7株致泻性大肠杆菌处于另一独立分支,原因可能与地区差异有关。
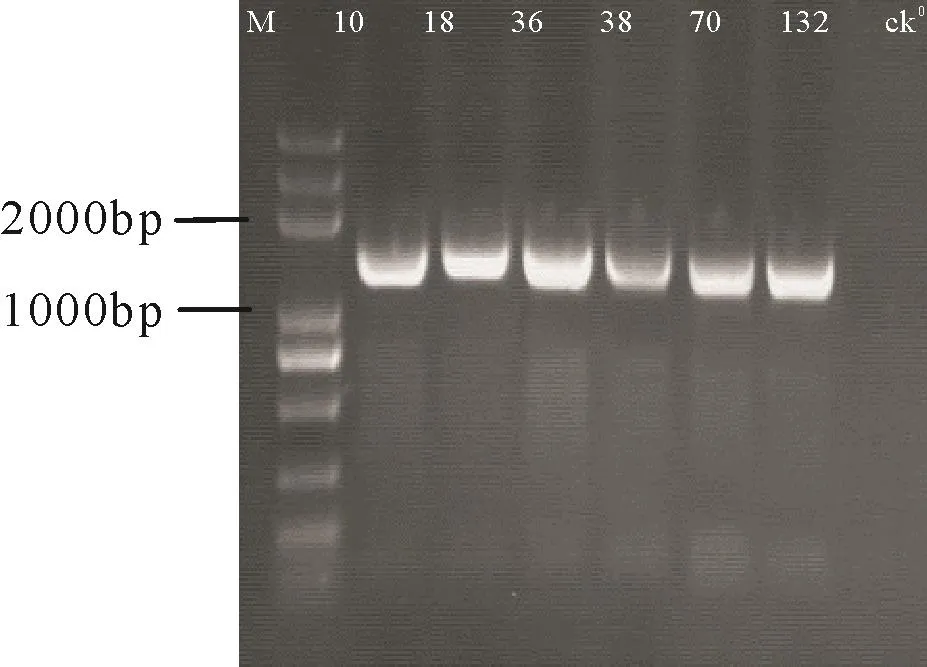
图4 部分致泻性大肠杆菌16S rDNA PCR产物电泳图
2.4 致泻性大肠杆菌和沙门氏菌污染状况分析
2.4.1 大中型屠宰场阳性率低于小型屠宰场 通过试验得出大中型屠宰场致泻性大肠杆菌阳性率为1.21%,小型屠宰场阳性率5.43%,大中型屠宰场的阳性率低于小型屠宰场(表3)。
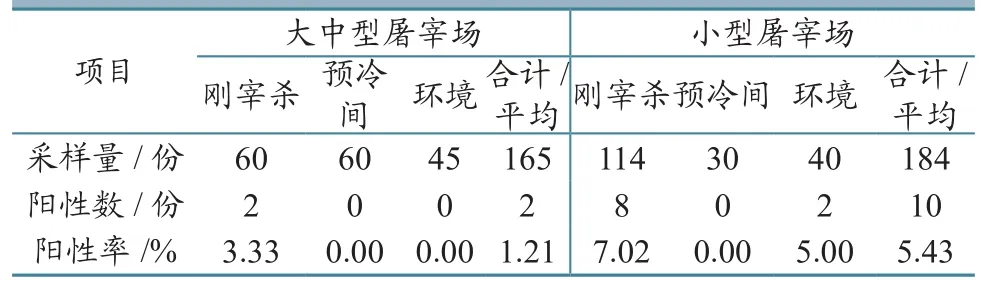
表3 不同规模屠宰场致泻性大肠杆菌试验结果
2.4.2 不同规模屠宰场的预冷间均未检出致泻性大肠杆菌 在174份刚宰杀的羊胴体棉拭子样品中检出10份致泻性大肠杆菌,阳性率为5.75%,而预冷间的样品中均未检出致泻性大肠杆菌。
2.4.3 沙门氏菌阳性率低,在羊屠宰环节的污染风险较小 349份棉拭子样品中,仅检出1株沙门氏菌,且来源于小型屠宰场环境样品,可见在目前调查的屠宰场中沙门氏菌的携带水平较低。
3 讨论

图5 致泻性大肠杆菌基于16S rDNA基因序列构建的系统发育树
3.1 刚宰杀的羊胴体表面是致泻性大肠杆菌的主要来源
本研究发现屠宰场中致泻性大肠杆菌的主要来源均是刚宰杀的羊胴体表面,阳性率分别为3.33%和7.02%。阳性菌株均为携带elt毒力基因的产肠毒素大肠杆菌。该菌可引起婴幼儿和旅游者腹泻,一般呈轻度水样腹泻,也可呈严重霍乱样症状,低热或不发热。腹泻常为自限性,一般2~3 d即可自愈[6]。张佳等[11]研究表明肉牛屠宰过程中,初始剥皮和去内脏是肉牛屠宰工序中造成胴体微生物污染的两个关键工序。由于大中型屠宰场在屠宰过程中工序分工严格、机械化程度较高和卫生消毒彻底等因素,其环境样品的致泻性大肠杆菌阳性率为0,而小型屠宰场为5.00%。
3.2 预冷间羊胴体表面未发现致泻性大肠杆菌和沙门氏菌污染
在349份棉拭子样品中,不同规模屠宰场的预冷间羊胴体表面均未检出致泻性大肠杆菌和沙门氏菌,在一定程度上说明屠宰过程中对羊肉胴体进行高压冲洗和排酸预冷工艺,有效地降低了屠宰环节羊肉产品致泻性大肠杆菌的携带水平。屠宰工艺中的预冷排酸是指在0~4 ℃、相对湿度 90%的冷藏条件下,将屠宰后的羊胴体预冷24 h。这一过程中胴体pH 值从活体的7.0~7.2 下降到5.5~6.5,表面形成油膜层,可抑制大多数微生物的生长繁殖,同时提高肉的品质[12-13]。
3.3 羊屠宰场各屠宰环节中沙门氏菌污染风险较小
本试验中仅检出1株沙门氏菌,污染风险较低。一项针对乌鲁木齐市屠宰场中3种食源性细菌的调查研究亦显示,羊屠宰场中未检出沙门氏菌[14],与本研究结果一致,说明在羊屠宰场沙门氏菌污染风险较小。食源性病原菌是引起食物中毒的主要原因,羊屠宰场是保证羊肉产品质量安全的关键环节。本试验中虽然对致泻性大肠杆菌和沙门氏菌的污染状况进行了分析,但有研究报道在羊屠宰场检出了单增李斯特菌[15],进口牛肉和屠宰场羊肉携带产志贺毒素大肠埃希杆菌O157:H7[15-16],因此在后续对内蒙古羊屠宰场的研究中,可进行相关试验的补充。
屠宰环节的畜禽产品安全与食品安全密切相关。食源性病原菌一旦污染肉品,在后续的速冻、储藏、运输和销售等环节仍能存活。若食用前处理不当或生熟区分不严格,很可能对公共卫生构成威胁[17]。因此,了解羊屠宰环节致泻性大肠杆菌和沙门氏菌的携带状况,不仅能及时掌握屠宰场病原菌的分布及水平,而且通过对羊源致泻性大肠杆菌的分离鉴定、毒力基因扩增和系统发育树分析,还能为羊源致泻性大肠杆菌的防控提供研究基础。
[1] 李鹏,王栋,孙晓东,等. 我国屠宰场动物疫病风险评估指标体系的构建[J]. 畜牧与兽医,2017,49(9):132-139.
[2] NAIM D A,YILMAZ E G,IRFAN E. Prevalence and molecular characterization of sorbitol fermenting and non-fermenting Escherichia coli O157:H7+/H7– isolated from cattle at slaughter house and slaughter house waste water[J]. International journal of food microbiology,2014(174):31-38.
[3] 刘建军,靳烨. 微生物与畜产品安全及控制措施[J].肉类工业,2007,317(9):45-47.
[4] 丁燕霞,柳增善. 市售鲜肉中细菌污染的安全性评价和耐药性分析[D]. 长春:吉林大学,2015.
[5] 谢建华,周莉,侯亚丽,等. 重庆市畜禽产品中沙门氏菌污染状况调查分析[J]. 中国动物检疫,2016,33(8):27-30.
[6] 卫生部. 食品安全国家标准食品卫生微生物学检验致泻大肠埃希氏菌检验:GB/T 4789.6—2016[S]. 北京:中国标准出版社,2016.
[7] 沈萍,陈向东. 微生物学实验[M]. 北京:高等教育出版社,2007:31-32.
[8] 卫生部. 食品安全国家标准食品卫生微生物学检验沙门氏菌检验:GB/T 4789.4—2016[S]. 北京:中国标准出版社,2016.
[9] 刘鲜鲜,王君玮,王娟,等. 山东部分生猪屠宰场屠宰环节沙门氏菌污染状况分析[J]. 中国动物检疫,2015,32(5):82-86.
[10] 王筱梦,孙芝兰,诸永志,等. 羊肉屠宰加工场主要污染菌的分布及其对熟制羊肉的致腐能力[J].食品科学,2017,38(16):261-267.
[11] 张佳,徐艳,霍晓伟,等. 肉牛屠宰工序微生物污染状况分析和喷淋减菌技术[J]. 食品与发酵工业,2011,37(10):209-213.
[12] 杨文婷,柏霜,罗瑞明,等. 排酸方式对成熟过程中滩羊肉品质和水分变化的影响[J]. 食品工业科技,2017,38(19):40-44.
[13] 贾雪晖,王小斌,靳烨,等. 不同排酸方式对巴美肉羊肉品质的影响[J]. 肉类研究,2012,26(5):14-16.
[14] 蔡扩军,齐新林,欧秀玲,等. 乌鲁木齐市屠宰场3种食源性细菌的污染情况调查[J]. 甘肃畜牧兽医,2016,46(21):91-92.
[15] ABUELHASSAN N N,MUTALIB S A,GIMBA F I,et al. Molecular characterization and phylogeny of Shiga toxin-producing E. coli(STEC)from imported beef meat in Malaysia[J]. Environmental science & pollution research international,2016,23(17):1-10.
[16] 李增魁,马兰,种伟雄. 西宁市售动物性食品中肠出血型大肠杆菌O157:H7 的检测[J]. 中国动物检疫,2011,28(7):56-59.
[17] 王娟,郑增忍,王玉东,等. 市售禽肉产品中沙门氏菌污染状况调查[J]. 中国动物检疫,2010,27(7):50.
