5-羟色胺诱导冠状动脉收缩的机制研究*
2018-05-17邓春玉邝素娟徐劲松
王 昊, 邓春玉, 饶 芳, 邝素娟, 杨 慧, 刘 林, 吴 琪, 徐劲松△
(1南昌大学第二附属医院, 江西 南昌 330006; 2广东省人民医院医学研究部,广东省医学科学院, 广东省心血管病研究所, 广东 广州 510080)
5-羟色胺(5-hydroxytryptamine,5-HT)作为神经递质,主要分布于松果体和下丘脑,但其亦可作为自体活性物质,储存于肠嗜铬细胞颗粒内。在刺激因素作用下,5-HT释放进入血液,被血小板摄取和储存,并可促进血小板聚集、血管平滑肌细胞(vascular smooth muscle cells,VSMCs)增殖和血管收缩。因此,5-HT可能在动脉粥样硬化、高血压和糖尿病等疾病的病理过程中发挥作用,进而促进心血管事件的发生[1]。
近年来的研究发现,5-HT参与诱导许多重要动脉的收缩。如有研究者发现,5-HT可通过激活三磷酸肌醇(inositol triphosphate,IP3)-Ca2+信号通路和二酰甘油(diacylglycerol,DAG)-蛋白激酶C(protein kinase C,PKC)信号通路,诱导胸主动脉收缩[2],并可以通过激活RhoA/Rho激酶和Src激酶刺激肠系膜动脉收缩[3]。此外,5-HT亦可诱导基底动脉收缩,其机制可能与线粒体单胺氧化酶(monoamine oxidase,MAO)介导过氧化氢生成增加有关[4],5-HT还可通过磷脂酶C(phospholipase C,PLC)/Orai1信号通路诱导冠状动脉收缩[5],以上研究结果提示5-HT通过参与多种信号通路诱导血管收缩,且机制不尽相同,而5-HT诱导冠状动脉收缩的机制研究较少,有待进一步阐明。
因此,本研究通过给予不同信号通路的抑制剂,观察5-HT诱导大鼠离体冠状动脉血管张力的变化,以研究5-HT诱导冠状动脉收缩的可能机制。
材 料 和 方 法
1 动物
SPF级雄性Wistar成年大鼠,体重300~350 g,由南方医科大学实验动物中心提供,许可证号为SCXK(粤)2016-0041。
2 主要试剂和仪器
5-HT、沙格雷酯(sarpogrelate)、U73122、硝苯地平(nifedipine)、SKF96365,2-氨基乙氧基苯硼酸(2-aminoethoxydiphenyl borate,2-APB)和卡马拉素(rottlerin)均购自Sigma;其余试剂均为国产分析纯。其中,nifedipine溶于DMSO,其余均溶于水。Krebs-Henseleit (K-H) 溶液(mmol/L): NaCl 119、NaHCO325、MgCl2·6H2O 1、KCl 4.7、KH2PO41.2、CaCl22.5和D-Glucose 11.1;高钾K-H溶液(mmol/L):NaCl 63.7、NaHCO325、MgCl2·6H2O 1、KCl 60、KH2PO41.2、CaCl22.5和D-Glucose 11.1,所用溶液配好通混合气(含95%O2+5% CO2)充分饱和,调pH约为7.4。
610M型多通道血管张力测定仪(DMT);Power Lab 8/30生物信号采集处理系统(AD);DK-8D型电热恒温水槽(上海医用恒温设备厂);Stemi DV4型体视显微镜(ZEISS)。
3 主要方法
3.1大鼠离体冠状动脉环的制备 采用CO2处死大鼠,用胶带固定在木板上,打开胸腔充分暴露心脏,取下大鼠的心脏。将取出的心脏立即放入4 ℃ 预冷,经混合气体(95% O2+ 5% CO2)饱和的K-H液中,并将心脏固定在玻璃皿中的硅胶板上。在冰浴条件下显微操作分离冠状动脉,小心去除冠脉周围的心肌组织,尽量避免钳夹、牵拉冠脉,以防损伤血管平滑肌,从而影响血管张力。将游离的冠状动脉剪成长度为1.8~2.0 mm的血管环。将血管环穿上2根直径40 μm的不锈钢丝,并平行固定在张力仪浴槽内的2个钳夹上,1个钳夹连接到螺旋测微器,用于调整冠脉的周长,另1个钳夹连接到内置的高灵敏度张力传感器,观察和测定血管环张力的变化。浴槽内预先放置5 mL 37 ℃ 预温的K-H液。对于冠状动脉,设定并调节基础张力至1.5 mN,平衡60 min,期间每隔15 min换液1次,并使张力维持在1.5 mN。在加入不同药物后,血管张力信号通过张力传感器,再经数模转换器输入计算机,完成张力记录。实验过程中槽内温度控制在(37±0.5) ℃,并往槽内持续通入95% O2和5% CO2的混合气体[6-7]。
3.2血管反应性及内皮完整性的测定 首先用高钾溶液刺激血管,检测血管的反应性。把浴槽内的K-H 液换成高钾K-H 液,使冠脉产生张力,维持5 min 后,充分冲洗(冲洗4次,每次间隔5 min)至基线,静息30 min 后,重复上述实验。前后2次高钾刺激后,血管收缩的幅度相差不超过10%可用于下一步实验。为检测血管内皮的完整性,重新平衡血管环30 min,先用血管收缩剂100 nmol/L U46619 引起血管持续性收缩,再加入血管舒张剂1 μmol/L 乙酰胆碱(acetylcholine,ACh),若血管的舒张幅度大于60%,视为内皮完整,若ACh不能使预先收缩的血管舒张,或者舒张程度小于10%,说明内皮去除完全。
4 统计学处理
以2次高钾诱发收缩张力的均值作为标准值(100%),各血管收缩剂诱发的张力变化的幅度用百分比来表示; 血管舒张剂是以收缩剂诱发的最大稳定收缩幅度定为标准值(100%),血管舒张/收缩的幅度以占标准值的百分比来表示,半数有效浓度(median effect concentration,EC50)表示产生 50% 最大效应时的药物浓度, pEC50= -lg(EC50)。采用 Prism 5.0软件对数据进行曲线拟合,绘制收缩及舒张曲线,求得 lgEC50和最大收缩或舒张百分比的变化值(Emax)。用SPSS 11.0统计软件分析,数据均采用均数±标准误(mean±SEM)表示。两组之间的显著性差异应用双侧t检验进行分析,多组间均数比较用单因素方差分析,组间均数的两两比较使用LSD法和SNK法。以P<0.05为差异有统计学意义。
结 果
1 5-HT2A受体阻断剂和PLC抑制剂对5-HT诱导的冠状动脉收缩反应的影响
给予单剂量(2 μmol/L) 5-HT使冠状动脉收缩,达到平台后,用5-HT2A受体阻断剂sarpogrelate (1 μmol/L)可完全阻断5-HT引起的冠状动脉收缩(P<0.01),见图1A、B。
采用累积给药法加入5-HT(0.01~10 μmol/L),可诱导冠状动脉呈浓度依赖性收缩;K-H液洗4次后,加入PLC特异性抑制剂U73122 (10 μmol/L和50 μmol/L),孵育30 min,结果发现2个浓度的U73122均可阻断5-HT引起的冠状动脉收缩(P<0.01),见图1C、D及表1。
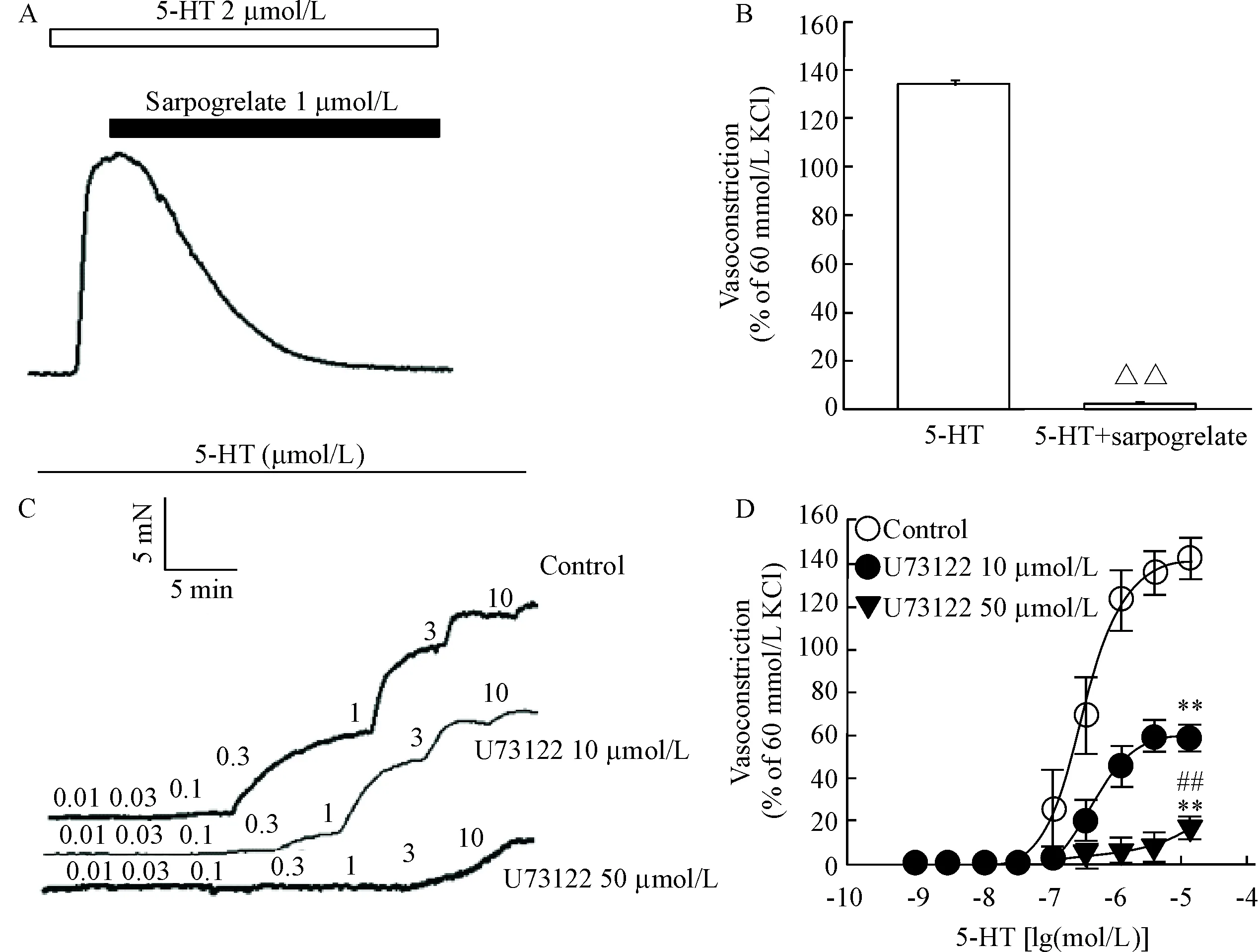
Figure 1. The effect of sarpogrelate and U73122 on vasoconstriction induced by 5-HT in endothelium-denuded coronary arterial rings from Wistar rats. A: representative recordings of vasoconstriction induced by 5-HT, then upon addition of sarpogrelate; B: sarpogrelate at 1 μmol/L significantly inhibited the contraction induced by 5-HT; C: representative recordings of concentration-dependent vasoconstriction induced by 5-HT; D: U73122 at 10 μmol/L and 50 μmol/L significantly inhibited the contraction induced by 5-HT in a dose-dependent manner. Mean±SEM.n= 4.△△P<0.01vs5-HT group;**P<0.01vscontrol group;##P<0.01vsU73122 10 μmol/L group.
图15-HT2A受体阻断剂sarpogrelate和PLC抑制剂U73122对5-HT诱导的冠脉收缩的影响
2 Rottlerin 和Y-27632对5-HT诱导的冠状动脉收缩反应的影响
2.1Rottlerin可抑制5-HT诱导的冠状动脉收缩反应 用上述方法记录5-HT累积给药后冠状动脉张力变化,再加入PKC抑制剂rottlerin 3 μmol/L和10 μmol/L孵育30 min。结果发现10 μmol/L的rottlerin可阻断5-HT引起的冠状动脉收缩(P<0.01),见图2A、B及表1。
2.2Y-27632可抑制5-HT诱导的冠状动脉环的收缩反应 同法记录5-HT累积给药后冠状动脉张力变化,再加入Rho相关蛋白激酶抑制剂Y-27632 3 μmol/L和10 μmol/L孵育30 min。结果发现2个浓度的Y-27632均可明显阻断5-HT引起的冠状动脉收缩(P<0.01),见图2C、D及表1。
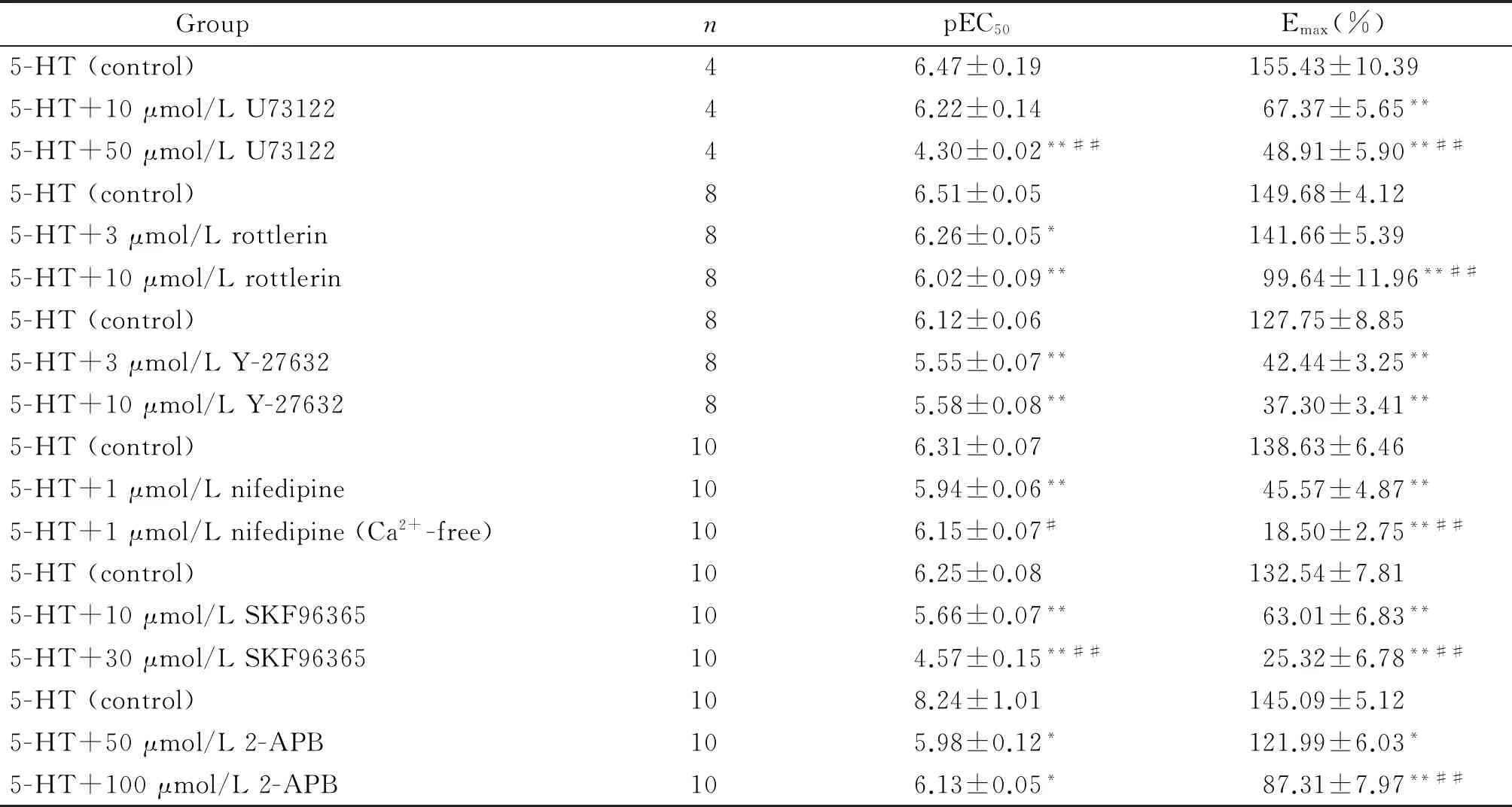
表1 5-HT诱导的冠状动脉收缩的pEC50和Emax (%)
*P<0.05,**P<0.01vscontrol group;#P<0.05,##P<0.01vslow concentration group.
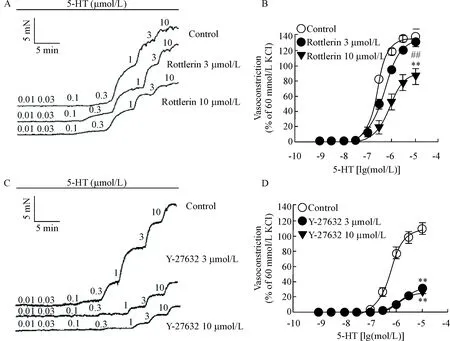
Figure 2. The effect of rottlerin and Y-27632 on vasoconstriction induced by 5-HT in endothelium-denuded coronary arterial rings from Wistar rats. A: representative recordings of concentration-dependent vasoconstriction induced by 5-HT in the presence of rottlerin; B: rottlerin at 10 μmol/L significantly inhibited contraction induced by 5-HT; C: representative recordings of concentration-dependent vasoconstriction induced by 5-HT in the presence of Y-27632; D: Y-27632 at 3 μmol/L and 10 μmol/L significantly inhibited contraction induced by 5-HT. Mean±SEM.n=8.**P<0.01vscontrol group;##P<0.01vsrottlerin 3 μmol/L group.
图2PKC抑制剂rottlerin和Rho相关蛋白激酶抑制剂Y-27632对5-HT诱导的冠脉收缩的影响
3 Nifedipine可抑制5-HT诱导的冠状动脉环收缩反应
同法记录5-HT累积给药冠状动脉张力变化,再加入L-型钙通道阻断剂nifedipine 1 μmol/L孵育30 min,结果发现nifedipine可明显阻断5-HT引起的冠状动脉收缩,见图3、表1。
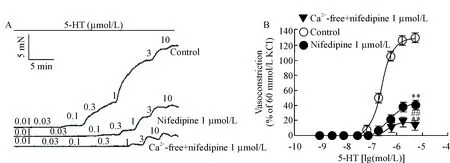
Figure 3. The effect of nifedipine on vasoconstriction induced by 5-HT in endothelium-denuded coronary arterial rings from Wistar rats. A: representative recordings of concentration-dependent vasoconstriction induced by 5-HT; B: nifedipine at 1 μmol/L significantly inhibited contraction induced by 5-HT. Mean±SEM.n=10.**P<0.01vscontrol group;##P<0.01vsnifedipine 1 μmol/L group.
图3L-型钙通道阻断剂nifedipine对5-HT诱导的冠脉收缩的影响
4 SKF96365和2-APB对5-HT诱导的冠状动脉收缩反应的影响
4.1SKF96365可抑制5-HT诱导的冠状动脉环的收缩反应 同法记录5-HT累积给药后冠状动脉张力变化,再加入钙库操纵性钙内流(store-operated Ca2+entry,SOCE)抑制剂SKF96365 10 μmol/L和30 μmol/L孵育30 min。结果发现2个浓度的SKF96365均可阻断5-HT引起的冠状动脉收缩,见图4A、B及表1。
4.22-APB可抑制5-HT诱导的冠状动脉环的收缩反应 同法记录5-HT累积给药后血管张力的变化,再加入SOCE抑制剂2-APB 50 μmol/L和100 μmol/L孵育30 min。结果发现给予的2个浓度2-APB均可阻断5-HT引起的冠状动脉收缩,见图4C、D及表1。
讨 论
本研究发现5-HT2A受体阻断剂sarpogrelate可完全消除5-HT引起的冠状动脉收缩;PLC抑制剂U73122、Rho相关蛋白激酶抑制剂Y-27632和PKC抑制剂rottlerin均可明显抑制5-HT引起的冠状动脉收缩;L型钙通道(Cav1.2)阻断剂nifedipine及SOCE抑制剂SKF96365和2-APB孵育后,5-HT诱导血管收缩张力较未处理组明显下降;同时,在含nifedipine的无钙K-H液中,发现5-HT仍可诱导血管收缩。
Sarpogrelate为5-HT2A受体阻断剂,具有抑制血小板聚集的作用,已应用于多种心血管疾病的治疗,如糖尿病、冠心病和动脉粥样硬化等。5-HT受体包括7个类型(5-HT1-7),除了5-HT3受体家族属于配体门控阳离子通道,其它5-HT受体家族均属于跨膜G蛋白偶联受体。而参与冠状动脉收缩的相关受体主要有5-HT1B和5-HT2A[8-9],本实验给予sarpogrelate干预后发现可完全消除5-HT引起的冠状动脉收缩,提示冠状动脉中是5-HT2A受体介导5-HT效应。
研究发现,5-HT可与VSMCs的细胞膜相应受体结合,激活PLC,使磷脂酰肌醇二膦酸(phosphatidylinositol bisphosphate,PIP2)分解为IP3和DAG。IP3与受体结合,使内质网上Ca2+排空,激活Ca2+内流通道,从而介导外钙内流,诱导血管收缩[10]。DAG激活PKC,使PKC增强性抑制蛋白17磷酸化,抑制肌球蛋白轻链磷酸酶活性,使其不易去磷酸化,导致平滑肌产生收缩增强[11]。U73122为PLC抑制剂,通过抑制PIP2水解为IP3,使细胞内Ca2 +的下降;Y-27632为Rho相关蛋白激酶特异性抑制剂,可靶向结合ROCK-1和ROCK-2的催化位点,从而抑制其活性[12]。本实验分别给予PLC/PKC/Rho激酶通路的抑制剂干预,发现均可明显抑制5-HT引起的冠状动脉环收缩,提示冠状动脉中5-HT2A受体介导冠脉收缩可能与PLC/PKC/Rho激酶信号通路有关。
此外,Ca2+作为第二信使,在平滑肌细胞中参与细胞收缩过程[13]。细胞内Ca2+通过与钙调蛋白结合后激活肌球蛋白轻链激酶,使肌球蛋白轻链磷酸化,导致肌动蛋白和肌球蛋白相互作用引起血管收缩。细胞膜上参与细胞内外钙离子交换的主要通道有:电压操纵性钙通道(voltage-gated calcium channel, VGCC)、SOC和受体操纵性钙通道(receptor operated calcium channel, ROCC)[14]。Sung等[15]发现在大鼠肠系膜动脉中,硝苯地平可显著降低5-HT诱导的动脉收缩。细胞内Ca2+浓度紊乱可引起冠状动脉收缩功能障碍。为研究冠状动脉VSMCs外钙离子交换的主要通道,本实验给予钙通道阻滞剂nifedipine,可阻断L-型钙通道抑制钙内流进入VSMCs;SKF96365和2-APB为SOCE抑制剂,可阻断SOCE从而降低胞质Ca2+,减少细胞外钙的进入[16]。本研究发现nifedipine、SKF96365和2-APB孵育后,5-HT诱导血管收缩张力较未处理组均明显下降;同时,在含nifedipine的无钙K-H液中,发现5-HT仍可诱导血管收缩,说明Cav1.2、SOC和内质网Ca2+释放可能与5-HT介导的冠状动脉收缩有关。
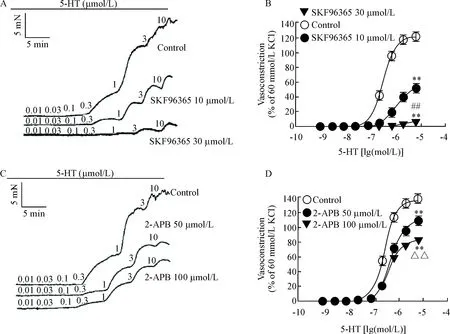
Figure 4. The effect of SKF96365 and 2-APB on vasoconstriction induced by 5-HT in endothelium-denuded coronary arterial rings from Wistar rats. A: representative recordings of concentration-dependent vasoconstriction induced by 5-HT in the presence of SKF96365; B: SKF96365 at 10 μmol/L and 30 μmol/L significantly inhibited contraction induced by 5-HT in a dose-dependent manner; C: representative recordings of concentration-dependent vasoconstriction induced by 5-HT in the presence of 2-APB; D: 2-APB at 50 μmol/L and 100 μmol/L significantly inhibited contraction induced by 5-HT in a dose-dependent manner. Mean±SEM.n=10.**P<0.01vscontrol group;##P<0.01vsSKF96365 10 μmol/L group;△△P<0.01vs2-APB 50 μmol/L group.
图4SOCE抑制剂SKF96365和2-APB对5-HT诱导的冠脉收缩的冠脉收缩的影响
本实验通过对大鼠冠状动脉张力的测定,发现5-HT通过激活5-HT2A受体诱导冠状动脉收缩,可能与PLC/PKC/Rho信号通路,以及L-型钙通道和SOC通道相关的钙调控有关,为相关疾病的治疗用药带来更多选择。但本实验仅对离体大鼠冠状动脉进行了分析,并未在细胞学水平对可能参与的信号通路进行深入研究,以进一步确定其上下游关系;且大鼠与人的冠状动脉存在种属间的差异,是其局限性。
[参 考 文 献]
[1] Vikenes K, Farstad M, Nordrehaug JE. Serotonin is associated with coronary artery disease and cardiac events[J]. Circulation, 1999, 100(5):483-489.
[2] 王 微, 宋二飞, 戚晓静, 等. 地西泮对5-羟色胺诱导的成年大鼠离体胸主动脉收缩的减弱作用及其机制研究[J]. 中西医结合心脑血管病杂志, 2010, 8(4): 446-448.
[3] Matsumoto T, Kobayashi T, Ishida K, et al. Enhancement of mesenteric artery contraction to 5-HT depends on Rho kinase and Src kinase pathways in theob/obmouse model of type 2 diabetes[J]. Br J Pharmacol, 2010, 160(5): 1092-1104.
[4] Poon CC, Seto SW, Au AL, et al. Mitochondrial monoamine oxidase-A-mediated hydrogen peroxide generation enhances 5-hydroxytryptamine-induced contraction of rat basilar artery[J]. Br J Pharmacol, 2010, 161(5):1086-1098.
[5] Deng CY, Yang H, Kuang SJ, et al. Upregulation of 5-hydroxytryptamine receptor signaling in coronary arteries after organ culture[J]. PLoS One, 2014, 9(9):e107128.
[6] Yang H, Kuang SJ, Rao F, et al. Species-specific differences in the role of L-type Ca2+channels in the regulation of coronary arterial smooth muscle contraction[J]. Naunyn Schmiedebergs Arch Pharmacol, 2016, 389(2):151-157.
[7] 邝素娟, 邓春玉, 杨 慧, 等. 离体大鼠冠状动脉的分离培养及张力测定技术[J]. 中国药理学通报, 2014, 30(3):137-141.
[8] Watts SW, Davis RP. 5-Hydroxytryptamine receptors in systemic hypertension: an arterial focus[J]. Cardiovasc Ther, 2011, 29(1):54-67.
[9] Nakamura E, Tanaka N, Kuwabara M, et al. Relative contributions of 5-hydroxytryptamine (5-HT) receptor subtypes in 5-HT-induced vasoconstriction of the distended human saphenous vein as a coronary artery bypass graft[J]. Biol Pharm Bull, 2011, 34(1):82-86.
[10] Raote I, Bhattacharya A, Panicker MM. Serotonin 2A (5-HT2A) receptor function: ligand-dependent mechanisms and pathways[M]∥ Chattopadhyay A. Serotonin Receptors in neurobiology. Boca Raton (FL): CRC Press/Taylor & Francis, 2007:1-17.
[11] 汪 燕, 周慧轩, 王 莉. 血管平滑肌钙动员和钙敏感机制在高血压中的改变[J]. 中国药理学通报, 2014, 30(2):287-291.
[12] Uehata M, Ishizaki T, Satoh H, et al. Calcium sensitization of smooth muscle mediated by a Rho-associated protein kinase in hypertension[J]. Nature, 1997, 389(6654):990-994.
[13] Cahalan MD. STIMulating store-operated Ca2+entry[J]. Nat Cell Biol, 2009, 11(6):669-677.
[14] Ambudkar IS, de Souza LB, Ong HL. TRPC1, Orai1, and STIM1 in SOCE: friends in tight spaces[J]. Cell Calcium, 2017, 63:33-39.
[15] Sung DJ, Noh HJ, Kim JG, et al. Serotonin contracts the rat mesenteric artery by inhibiting 4-aminopyridine-sensitive Kv channels via the 5-HT2A receptor and Src tyrosine kinase[J]. Exp Mol Med, 2013, 45(12):e67.
[16] Chen T, Zhu J, Zhang C, et al. Protective effects of SKF-96365, a non-specific inhibitor of SOCE, against MPP+-induced cytotoxicity in PC12 cells: potential role of Homer1[J]. PLoS One, 2013, 8(1):e55601.
