寄生于鲫的锚首虫一新种
2018-03-30夏晓勤黄晓丽石米娟程莹寅张婉婷陆承平
夏晓勤 黄晓丽 石米娟 程莹寅 张婉婷 陆承平
(1. 中国科学院水生生物研究所, 武汉 430072; 2. 南京农业大学动物医学院, 南京 210095; 3. 中国科学院大学, 北京 100049)
鲫(Carassius auratus)生命力强, 肉质鲜美, 经遗传改良的品种生长迅速, 是我国的重要经济鱼类。寄生于鲫的单殖吸虫逾二十种, 分别隶属指环虫(Dactylogyridae)、双身虫(Diplozoonidae)和三代虫(Gyrodactylidae)三个科, 但淡水单殖吸虫的另一主要类群锚首虫科(Ancyrocephalidae)尚无报道[1]。作者于1999年5月在鲫鳃部获得两锚首虫标本, 其几丁质结构与寄生于鳜(Siniperca chuatsi)的河鲈锚首虫(Ancyrocephalus mogurndae)很相似, 但尺寸略大, 连接片形状有所不同[2]。其他文献也未见与此虫相似的几丁结构[2—5], 因此鉴定为一新种, 命名为鲫锚首虫(Ancyrocephalus carassii sp. nov.), 模式标本保存于中国科学院水生生物研究所。这是在鲫中首次发现锚首虫科(Ancyrocephalidae)的单殖吸虫寄生。
1 材料与方法
按传统方法进行鱼类病原调查[6]。宿主鲫购于南京市卫岗童卫路市场, 取下鳃片后在解剖镜下观察, 并挑取虫体, 用75%酒精固定之后, 经布氏胶固定封片, 在显微镜下测量其几丁质结构, 绘制特征图。
2 结果
所绘制的虫体后吸器与交配器结构见图 1A,新种量度特征描述如下。
鲫锚首虫的虫体较大, 长1.12 mm, 宽0.32 mm。因收集到标本时, 其死亡已久, 虫体膨胀, 此数据仅作参考。后吸器由7对边缘小钩、2对大小接近的中央大钩和2个联结棒组成。
背中央大钩粗壮, 基部膨大, 无明显分叶, 全长0.073 mm, 钩柄长0.056 mm, 钩尖长0.028 mm, 内突长0.035 mm。背联结棒长0.015 mm, 宽0.1 mm, 其两端稍膨大, 中部两小突起, 似由两直棒小角度相交愈合而成。
腹中央大钩全长0.059 mm, 钩柄长0.047 mm,钩尖长0.02 mm, 内突长0.03 mm, 外突长0.012 mm。腹联结棒略呈弧形, 长0.014 mm, 宽0.078 mm。
边缘小钩雏形, 全长0.025 mm, 钩尖长0.007 mm,钩柄长0.016 mm。交接管极细长, 并回旋盘曲, 管长约0.256 mm, 基部呈球状膨大, 直径约0.012 mm,其支持器短小, 长0.033 mm, 有心形突起。阴道管长约0.119 mm, 盘曲成圈, 阴道口呈喇叭状, 与支持器相靠, 叶形的支持器两端向翘向同一侧, 长约0.45 mm。
鲫锚首虫的分类结构与河鲈锚首虫(图 1B)较为相似, 其中央大钩、边缘小钩, 以及交配器均属同一类型, 形态很接近, 主要差异在鲫锚首虫的背联结棒上有两个突起; 且背中央大钩基部较后者更粗壮, 虽无外突, 但在着生着生的位置成一尖角, 而河鲈锚首虫在相应部位为平削状; 鲫锚首虫的一对中央大钩在近钩顶处有一明显的小凹, 此结构未见于河鲈锚首虫。此外, 两虫在量度上的差别也较明显, 鲫锚首虫的各结构均较河鲈锚首虫大(表 1)。
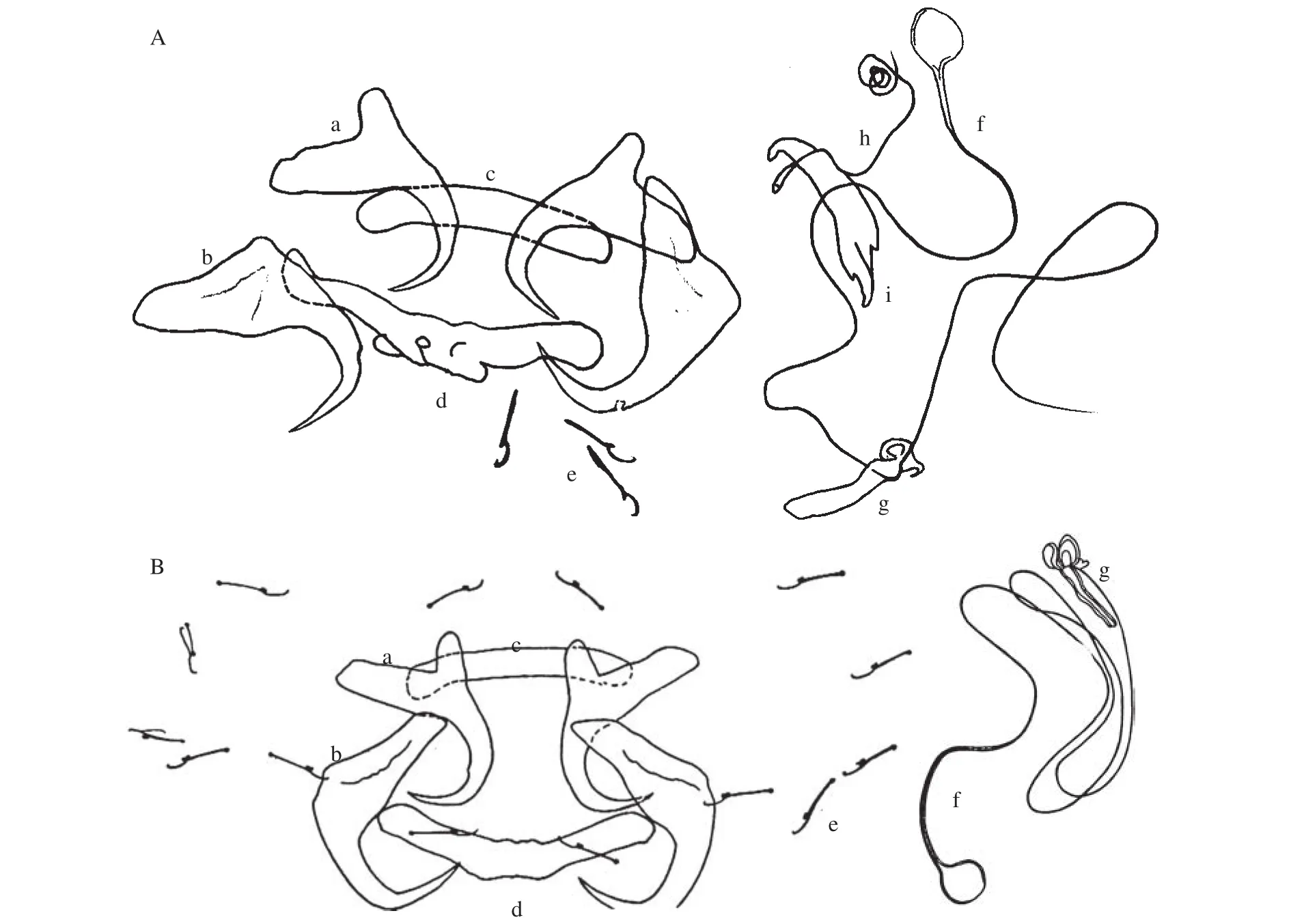
图 1 鲫锚首虫(A)与河鲈锚首虫(B, 引自文献[2])的几丁质结构Fig. 1 The chitinous structures of Ancyrocephalus carassii sp. nov. (A) and Ancyrocephalus mogurndae (B)
3 讨论
寄生于鲫属鱼类的单殖吸虫种类较多, 共计有21种之多, 其中包括13种指环虫, 5种三代虫和3 种双身虫, 没有一种锚首虫据统计[1,7—9], 故此例为鲫鱼中首次发现锚首虫的寄生。鲫鱼在分类中属于鲤科(Cyprinidae), 鲤亚科(Cyprininae), 鲫属(Carassius)。中国是鲤科鱼类的发祥地, 该科鱼类种类繁多, 但寄生于鲤科鱼类的锚首虫科单殖吸虫仅29种, 且大多分布于逗亚科中[10], 鲤亚科中仅在鲤(Cyprinus carpio)中报道过宿主特异性较弱的两种锚首虫: 近相等锚首虫(Ancyrocephalus subaequlis Achmerow, 1952)和奇异似鲶盘虫[Silurodiscoides exima (Gussev et Strelkow, 1960) Ling,1973]。前者可寄生于逗亚科(Gobioninae)的花逕(Hemibarbus maculatus Bleeker)、唇逕(Hemibarbus labeo)和似逗未定种(Pseudogobio sp.), 以及鲃亚科(Barbinae)的瓣结鱼(Tor (Folifer) brevifilis Wu,1977); 后者则发现于鲶科(Siluridae)的鲶(Silurus asotus Linnaeus)和大口鲶(Silurus soldatovi meridionalis Chen), 以及鲤科襁亚科(Danioninae)的宽鳍鲱(Zacco platypus Temminck et Schlegel)、鲌亚科(Culterinae)的华鳊和逗亚科的花逕; 鲫锚首虫则是首先发现于鲤亚科鱼类的首例锚首虫科单殖吸虫。
在我国内陆水体的单殖吸虫中, 锚首虫科的种类数有100余种, 占比近20%, 仅次于指环虫科[1,11]。相较于360种指环虫科(Dactylogyridae)单殖吸虫仅分布于8科宿主鱼中, 其锚首虫的宿主分布范围很广, 达19科。尽管如此, 锚首虫的宿主特异性很高,近65%的种类仅寄生于单一的宿主[10]。通过对鲫锚首虫和河鲈锚首虫的比较, 发现两者在整体上很相似, 但在结构细节和量度上又有实质性的差异,两者宿主差异也大, 说明两者虽非同种, 但亲缘关系很近, 可能是通过宿主转移(Host-switching)分化而成。鱼类寄生虫调查中鲫鱼是一种很常见的调查对象, 既然以前从未发现过鲫锚首虫, 说明该虫很罕见; 相比之下, 而河鲈锚首虫较为常见。因此我们猜测更有可能是后者通过偶然的机会转移到鲫上寄生, 进而分化形成新种。目前所报道河鲈锚首虫的宿主均为肉食性的鲈形目(Perciformes)鱼类,包括逇科(Serranidae)的鳜(Siniperca chuatsi Basilewsky)、大眼鳜(Siniperca kneri Garman)、斑鳜(Siniperca scherzeri Steindachner)和长身鳜(Coreosiniperca roulei Wu), 以及塘鳢科(Eleotridae)的沙塘鳢(Odontobutis obscurus Temminck et Schlegel)和鳅虎鱼科(Gobiidae)的条纹长颌鳅虎(Chaenogobius sp.)。宿主切换是如何在这些宿主与鲫之间发生,到底是通过纤毛幼虫感染, 还是通过成虫在宿主发生接触时直接接触转移? 尚不得而知。
参考文献:
[1]Wu B H, Lang S. Wang W J, et al. Fauna China, Platyhelminthes, Monogenoidea [M]. Beijing: Science Press.2000, 305—313 [吴宝华, 郎所, 王伟俊, 等. 中国动物志,扁形动物门, 单殖吸虫纲. 北京: 科学出版社. 2000,305—313]
[2]Hubei Institute of Hydrobiology. Hubei Fish Disease Pathogen Flora Disposition [M]. Beijing Science Press.1973, 57—93 [湖北省水生生物研究所, 湖北省鱼病病原区系图志. 北京:科学出版社. 1973, 57—93]
[3]Gussev A V. Key to the Parasites of Freshwater Fish Fauna of the USSR [M]. Leningred: Science Publishing Company. 1985, 1—425 (in Russian)
[4]Wu B H, Sun X D, Song C C. Fauna of Zhejiang (Trematoda) [M]. Hangzhou: Science Technology Press. 1991,34—276 [吴宝华, 孙希达, 宋昌存. 浙江动物志(吸虫类). 杭州: 浙江科学技术出版社. 1991, 34—276]
[5]Yamaguti S. Systema Helminthum. Vol. IV. Monogenea and Aspidocotylea. John Wiley and Sons [M]. New York:Interscience Publishers. 1963, 1—699
[6]The Fish Disease Laboratory of Institute of Hydrobiology, Chinese Academy of Sciences. The Handbook of Fish Disease Examination, Second edition [M]. Shanghai:Science Technology Press. 1981, 1—93 [中国科学院水生生物研究所鱼病学研究室. 鱼病调查手册. 2版. 上海:上海科学技术出版社. 1981, 1—93]
[7]Xia X Q. Ecological studies on Dactylogyrus spp. and the distribution feature of Monogenea in inland water of China [D]. Thesis for Doctor of Science. Institute of Hydrobiology, Chinese Academy of Sciences, Wuhan. 1997[夏晓勤. 指环虫的生态学及我国内陆水体单殖吸虫的分布特征. 博士学位论文, 中国科学院水生生物研究所,武汉. 1997]
[8]Ma X R, Wang G T, Zou H, et al. Community structure of the Dactylogyrus species on the gills of wild and cultured crucian carp (Carassius auratus) [J]. Acta Hydrobiologica Sinica, 2014, 38(1): 137—141 [马幸荣, 王桂堂,邹红, 等. 野生与人工养殖鲫鳃部寄生指环虫的群落结构. 水生生物学报, 2014, 38(1): 137—141]
[9]Li R R, Li W X, Wu X D, et al. Identification of Gyrodactylus species in goldfish (Carassius auratus) through morphological study and the analysis of the rDNA ITS sequence [J]. Acta Hydrobiologica Sinica, 2014, 38(5):903—909 [李冉冉, 李文祥, 吴旭东, 等. 金鱼寄生三代虫的形态学及基于rDNA ITS 序列的分子鉴定. 水生生物学报, 2014, 38(5): 903—909]
[10]Xia X Q, Wang W J, Yao W J. The host-specificity of monogeneans in inland waters of China [J]. Acta Ecolo-gica Sinica, 2000, 20(4): 594—597 [夏晓勤, 王伟俊, 姚卫建. 我国内陆水体单殖吸虫的宿主特异性. 生态学报,2000, 20(4): 594—597]Xia X Q, Wang W J, Yao W J. The composition of mono-
[11]genea in inland waters of China [J]. Journal of Hunan Agricultural University, 1999, 25(1): 57—61 [夏晓勤, 王伟俊, 姚卫建. 中国内陆水体单殖吸虫的种类组成. 湖南农业大学学报, 1999, 25(1): 57—61]
