Chemerin、肥胖与运动研究进展
2018-03-23宋威巍柏友萍
宋威巍, 柏友萍
(安徽师范大学 体育学院,安徽 芜湖 241003)
脂肪组织不仅是能量储存仓库还是一个内分泌器官,能够分泌多种具有生物学活性的脂肪细胞因子,在肥胖和肥胖相关疾病中发挥着重要作用[1,2]。Chemerin是2007年发现的一种新的脂肪细胞因子,参与调节脂肪细胞的分化、体内的物质能量代谢、肥胖和胰岛素抵抗等的病理生理过程[3-5]。本文主要对Chemerin、肥胖与运动研究进展进行分析综述。
1 Chemerin的概述
1.1 Chemerin的发现
Chemerin也被称作视黄酸受体反应蛋白2和他扎罗汀诱导基因2,是1997年Nagpal[6]等用维甲酸类药它扎罗汀治疗银屑病时克隆出来,最初被视为一种在银屑病中的新型的视网膜下腔应答基因,维甲酸类药物可调节其基因表达。2003年,Wittamer[7]等从卵巢癌患者的腹水中提取出Chemerin,并证明是G蛋白偶联。
受体ChemR23的天然配体,因巨噬细胞和未成熟树突状细胞对其有趋化作用,命名为趋化素。Chemerin受体ChemR23的发现早于Chemerin,ChemR23是1996年Owman[8]等从B淋巴母细胞c-DNA文库中鉴定出的新的cDNA序列,G蛋白偶联受体(G-protein coupled receptor,GPCRs)家族与其编码蛋白质高度同源,将其命名为ChemR23,主要在白细胞、脂肪细胞、内皮细胞、上皮细胞、破骨细胞以及血管平滑肌细胞中表达。直到2007年,Bozaoglo[9]等采用信号序列捕捉技术确定Chemerin是一种新的脂肪细胞因子,在脂肪组织和肝脏组织中都有表达,该表达与脂肪细胞分化高度相关。Goralski[10]等发现,Chemerin在小鼠的肝脏和胎盘中表达较高,尤其是在小鼠附睾的白色脂肪组织细胞中的表达,该表达量是间质血管上皮细胞中表达量的两倍。因此,Chemerin表达的主要作用靶点可能是白色脂肪组织。Chemerin和ChemR23结合后,ChemR23发生内吞噬作用促使细胞内钙离子释放,通过结合G蛋白偶联的异源三聚体来抑制环磷酸腺苷(cyclic adenosine monophosphate,cAMP)的积聚,从而调控各种代谢过程和炎性反应[11]。G蛋白偶联感受器(G-protein coupled receptor,GPR)1和趋化因子类受体(chemokine receptor-like,CCRL)2也是Chemerin的两个受体,与ChemR23的共同作用是促进脂肪细胞的分化及葡萄糖的转运[12,13]。
1.2 Chemerin的分子结构与理化特性
人类 Chemerin 的相对分子量为18kD,编码基因全长3289bp,是由163 个氨基酸组成,位于染色体7p36上,其mRNA长度为734bp,由5个内显子和6个外显子组成[14]。在血循环中,Chemerin是以收缩为3nm的前体pro-Chemerin形式存在的趋化蛋白,它为前体形式分泌的蛋白质。人类Chemerin的前体 pro-Chemerin是由20个疏水肽和163个氨基酸组成的复合体,成熟的 pro-Chemerin 是一个由 143 个氨基酸组成的多肽(Chem21-163),有较低的趋化活性[15]。为增加其生物活性,在C-末端结构域,pro-Chemerin需要被位于外胱抑素状褶皱的蛋白水解酶处理[16]。Chemerin的基因结构与内含子的数目和位置显著相关[17]。Chemerin在白色脂肪组织、肝脏和胎盘中表达相对较高,而在肺、棕色脂肪组织、心脏、肾脏和骨骼肌中表达相对较低[18]。Chemerin是G蛋白偶联受体的趋化因子受体(chemokine-like receptor,CMKLR)1的天然配体和趋化信号,它也可与GPR1结合。CMKLR1在巨噬细胞、未成熟的树突细胞和白色脂肪组织中表达较高,在肺、心脏、脑和胎盘的表达相对较低,而GPR1主要在脂肪组织中表达[19]。Chemerin的第三种配体趋化因子受体CCRL2与Chemerin的结合是非内在化的,不具有受体信号的作用[20]。CCRL2在体内与Chemerin的结合是局部的,增加局部Chemerin的浓度,有利于CMKLR1和潜在GPR1介导过程的发生(见图1)。
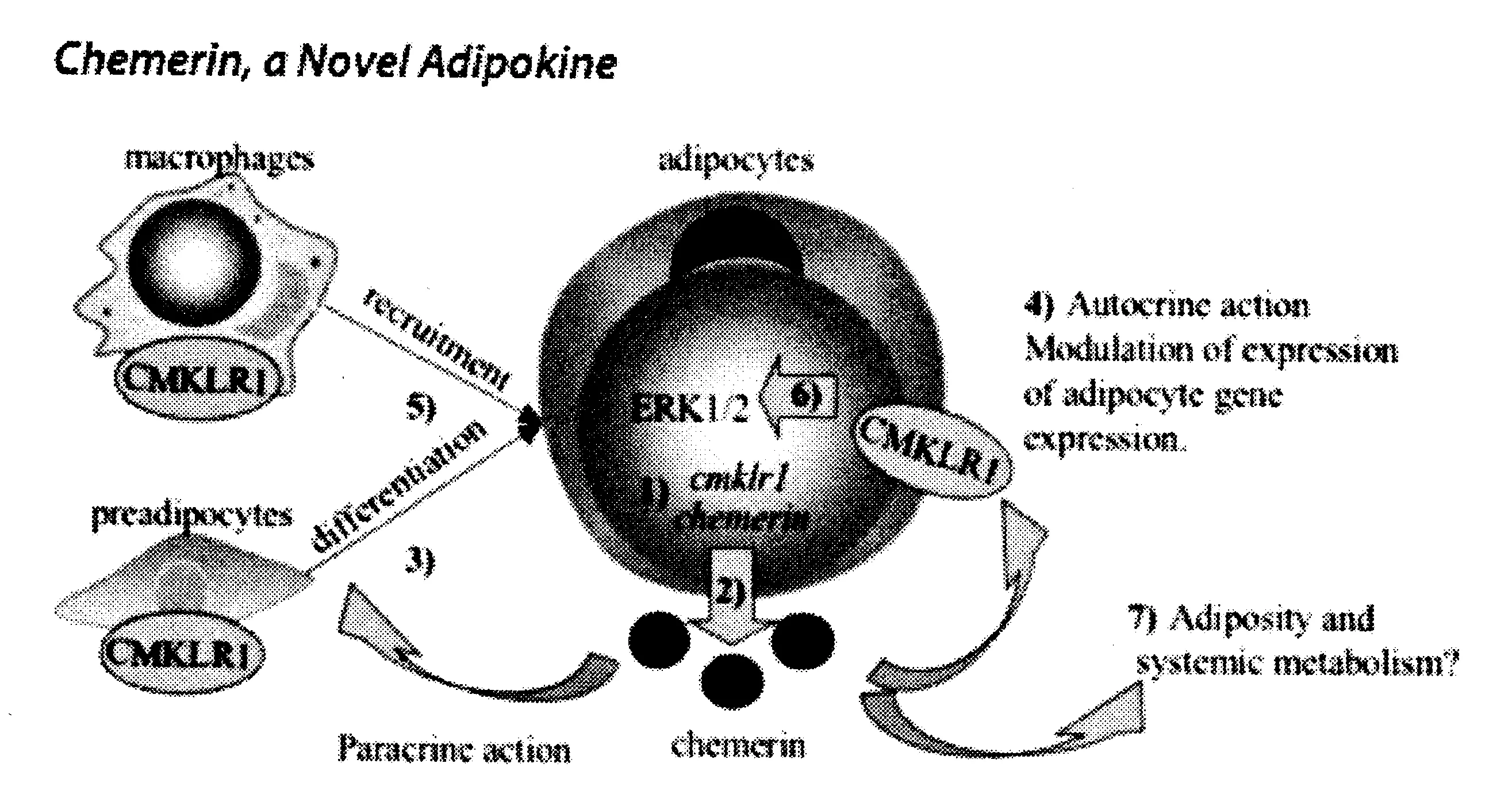
图1 Chemerin,一个新的脂肪细胞因子(Kerry B,Goralski et al,2007)
2 Chemerin与肥胖相关的疾病
肥胖一般是指体内白色脂肪组织中的脂肪细胞数目增多或体积增大,导致脂肪过度堆积而引起代谢紊乱。高脂、高热量的膳食和缺乏运动是导致单纯性肥胖的主要原因,现代人肥胖的产生大多是因为身体活动的减少和久坐行为的增加及高热能的膳食[21]。陆维等[22]的流行病学研究证明肥胖容易并发胰岛素抵抗、血脂异常、高血压等,并且是它们的独立危险因素。Polednak对肥胖与癌症的关系的临床研究发现,肥胖人群患乳腺癌、腺癌、肠癌、甲状腺癌等恶性肿瘤的发病率比非肥胖健康人群明显升高,可能原因为肥胖引起体内部分脂肪细胞因子的升高,增加恶性肿瘤的发病率[23]。Bozaoglu等[24]对Chemerin研究发现,肥胖人群中常伴随高胰岛素血症,且血清Chemerin水平也偏高。余岚等[25]研究发现,肥胖青少年中血清Chemerin水平明显增加,肥胖青少年的代谢紊乱症候群与Chemerin水平升高密切相关。另外,Rima[26]等通过对肥胖小鼠的研究发现,肥胖小鼠Chemerin水平较对照组显著升高。因此,肥胖可能参与上调Chemerin的表达水平。
2.1 Chemerin与胰岛素抵抗及2型糖尿病
胰岛素抵抗是肥胖和2型糖尿病共同的病理生理基础,胰岛素抵抗伴随2型糖尿病发生、发展的整个过程[13]。肥胖人群的脂肪组织对胰岛素敏感性下降,而产生胰岛素抵抗,使胰岛β细胞代偿性的分泌更多的胰岛素,但当胰岛β细胞功能缺陷不代偿时,血糖水平就会升高,逐渐演变为2型糖尿病[27]。白色脂肪组织过多积累,分泌脂肪细胞因子Chemerin参与胰岛素抵抗、高血压、糖尿病以及代谢综合症等肥胖相关疾病的发生(见图2)[28]。Takahashi等[29]采用体外3T3-L1细胞培养的方法发现,Chemerin在脂肪细胞中能够通过激活胰岛素信号转导通路和某些氧化应激,调节脂肪组织对胰岛素的敏感性而提高葡萄糖摄取。但Sell等[30]采用体外骨骼肌细胞培养的方法发现,Chemerin在骨骼肌细胞中通过增加蛋白激酶上游胰岛素底物受体-1丝氨酸磷酸化水平,降低下游信号分子的活性,减少胰岛素刺激下的葡萄糖摄取,降低骨骼肌对胰岛素的敏感性,产生胰岛素抵抗。临床研究发现[31],胰岛素抵抗人群中血清Chemerin水平显著高于正常人群,在肥胖2型糖尿病患者体内,血清Chemerin水平明显增高。Lloyd[32]等通过大鼠实验发现,高脂膳食提高脂肪和肌肉中的Chemerin基因和受体的表达,但对胰腺没有显著影响。Chemerin是近年来新发现的脂肪细胞因子,在肥胖、胰岛素抵抗及2型糖尿病中的水平均有改变,但大多数是从横断面进行研究,所以我们需要设计良好的实验和临床研究验证Chemerin在肥胖和肥胖相关疾病中扮演着协同还是拮抗的角色。
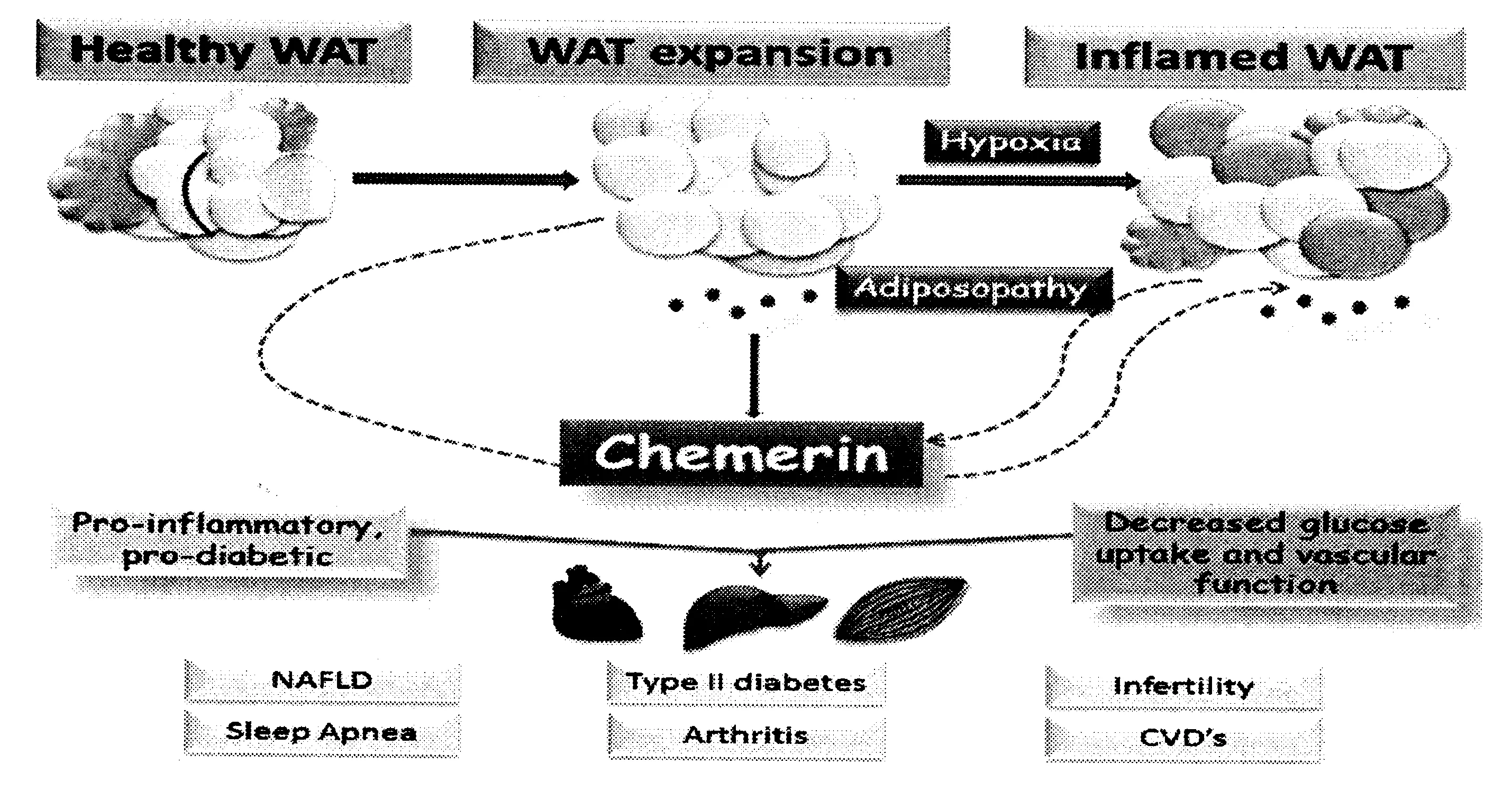
图2 Chemerin与肥胖相关疾病的作用机制(Fatima SS,et al.2014)
2.2 Chemerin与炎症
Chemerin与炎症的关系主要是通过CMKLR1发挥作用,Chemerin通过CMKLR1提升未成熟的树突细胞和巨噬细胞的趋化[33]。Chemerin及其受体CMKLR1影响免疫细胞的损伤部位,并可能参与炎症的发生、发展过程[18]。肥胖病人的血清Chemerin水平与代谢综合征的各个方面都呈正相关,并且血清Chemerin水平与促炎细胞因子(proinflammatory cytokine)、肿瘤坏死因子(tumor necrosis factor,TNF)-α、白细胞介素(interleukin,IL)-6和C反应蛋白(C-reactive protein)水平相关[34]。在病理学上,Chemerin在银屑病皮损中表达,先天免疫系统的几个效应细胞,如树突细胞、单核细胞、巨噬细胞和自然杀伤细胞(natural killer cells,NK)与银屑病发病机制相关,Chemerin表达被认为是早期银屑病病变的标志,能够提升类浆细胞和树突细胞的活化[35]。相关研究发现,Chemerin及其受体的表达在炎症时会有升高,如ChemR23在吸烟诱发的肺细胞炎症中表达较高[36],Chemerin表达水平在慢性阻塞型肺部疾病患者中显著升高[37]。Ozcan[38]等研究牙周炎疾病时发现,牙周炎患者唾液中Chemerin浓度显著升高。由此可以猜测,Chemerin表达水平提高可能成为很多炎症状态的预测因子,但要应用于临床还要进一步研究。
3 Chemerin与运动
肥胖是一种疾病,同时是心脑血管病、糖尿病、某些恶性肿瘤等的危险因素之一[39]。减肥是促进肥胖人群身体健康,提高生活质量的有效手段,而有规律性的运动是抑制脂肪累积的重要手段。正常的体重取决于热能的平衡,通过运动和减少能量的摄入可以减少初始体重的9%-10%[40]。运动减肥既可达到减肥的效果,又能促进身体健康,在减肥方式中应用广泛。研究表明[41],运动能够减少肥胖鼠和瘦鼠的脂肪量和obmRNA水平,运动增加能量消耗,使ob基因减少对能量消耗增加的反应性表达,使体重维持在一个相对稳定的水平。下丘脑神经肽Y(neuropeptide Y,NPY)是一种很强的增食欲激素,主要由下丘脑弓状核神经元分泌和释放,在摄食行为和能量消耗的整合中起重要作用[42],长期的有氧运动导致体重下降的重要原因,可能是其能够抑制NPY的基因表达,使得NPY蛋白合成减少[43]。柏友萍[44]等的研究表明,对超重肥胖大学生进行8周运动强度为F.C.的60%-70%的持续运动干预,超重肥胖大学生的肥胖指数、体脂百分比、腰臀比、体内脂肪重量都发生较为显著的变化。李娟[45]等对肥胖青少年进行6周的有氧运动干预发现,有氧运动能够有效降低肥胖青少年人群的体重,改善其体成分,调节脂肪代谢及其提高胰岛素敏感性。关于运动对肥胖的影响研究已取得一系列成果,但作用机制还不完善,有待进一步研究。
以胰岛素抵抗为发病基础的代谢性疾病与机体慢性炎性反应有密切关系,而科学的、有规律的运动对缓解和预防胰岛素抵抗及心血管系统疾病具有良好的作用。有研究表明,有氧运动可以调节一些脂肪细胞因子和炎症因子的水平,来影响肝脏胰岛素抵抗的形成[46]。刘燕[27]等的研究发现,高脂膳食可诱导肥胖的发生,并升高血清Chemerin的表达,且Chemerin的表达升高与血糖血脂代谢紊乱和胰岛素抵抗有着紧密联系。Chemerin作为一种新的脂肪细胞因子,运动对其影响研究近年有所报道。韩晓菲[20]等给予实验大鼠游泳运动和药物干预,发现大鼠体重减轻、空腹血糖、胰岛素水平下降、胰岛素抵抗降低,血清Chemerin水平也显著下降。丁锐[47]对高脂膳食大鼠运动与安静的不同状态下肝脏组织Chemerin /CMKLR1-mRNA表达水平的变化进行研究发现,有氧运动可以使高脂膳食大鼠肝脏CMKLR mRNA/Chemerin表达水平显著降低。Mehrdad[48]等对24只小鼠进行8周跑台运动干预发现,小鼠血清Chemerin水平显著降低。由以上可推测,运动可能通过减少体重来降低Chemerin的表达。
综上所述,Chemerin是一种新的脂肪细胞因子,明确了Chemerin的结构、基因表达、生物学功能。研究发现Chemerin参与肥胖、胰岛素抵抗、2型糖尿病、炎症的发生与发展,Chemerin表达水平降低能够改善糖耐量增加葡萄糖的利用,是否能作为判断肥胖导致相关疾病的重要指标有待进一步研究。从运动对Chemerin的干预研究发现有一定的影响,但处于初始阶段,运动是否通过对Chemerin影响而预防肥胖相关疾病及作用的机制,还需要大量实验和临床研究证明。
[1] 吴孟水,宁翠丽,刘宽芝.Chemerin的研究进展[J].医学综述,2016,22(11):2102-2106.
[2] H,JAMES HARWOOD JR.The adipocyte as an endocrine organ in the regulation of metabolic homeostasis[J]. Neuropharmacology,2012,63(1):57-75.
[3] 陈晨,柏友萍.脂肪因子Chemerin与运动[J].安徽师范大学学报(自然科学版),2015,38(3):301-306.
[4] PAULINA KULIG, TOMASZ KANTYKA, BRIAN A ZABLE, et al. Chemerin is an Antimicrobial Agent in Human Epidermis[J]. PLOS One,2013,8(3):1-8.
[5] KERRY B GORALSKI, TANYA C MCCARTHY, ELYISHA A HANNIMAN et al. Chemerin, a Novel Adipokine That Regulates Adipogenesis and Adipocyte Metabolism[J]. THE JOURNAL OF BIOLOGICAL CHEMISTRY, 2007,282(38):28175-28188.
[6] MATTHEW C, ERNST AND CHRISTOPHER J SINAL. Chemerin: At the crossroads inflammation and obesity[J]. Cell, 2010,21(11):660-667.
[7] WITTAMER V, FRANSSEN JD, VULCANO M, et al. Specific recruitment of antigen-presenting cells by chemerin,a novel processed ligand from human inflammatory fluids[J]. J Exp Med,2003,198(7):977-985.
[8] OWMNC,NILSSON,LOLAIT SJ.Cloning of cDNA encoding a putative chemoattractant receptor[J].Genomics,1996,37(2):187-194.
[9] ROURKE J L, DRANSE H J, SINAL C J. Towards an integrative approach to understanding the role of chemerin in human health and disease[J].Obesity Reviews,2013,14(3):245-62.
[10] GORALSKI K B, MC CARTHY T C, HANNIMAN E A, et al. Chemerin,a novel adipokine that regulates adipogenesis and adipocyte metabolism.J Biol Chem,282(38):28175-28188.
[11] PENG L, YU Y, LIU J, et al.The chemerin receptor CMKLR1 is a functional receptor for amyloid-beta peptide[J].J Alzheimers Dis,2015,43(1):227-242.
[12] RHEE E J.Chemerin: a novel link between inflammation and atherosclerosis? Diabetes Metab[J]. Diabetes Metab, 2011,35(3):216-218.
[13] BONDUE B,WITTAMER V,PARMENTIER M.Chemerin and its receptors in leukocyte trafficking, inflammation and metabolism[J].Cytokine Growth Factor Rev,2011,22(5-6):331-338.
[14] 王婷,焦凯.Chemerin与胰岛素抵抗的关系[J].陕西医学杂志,2014,43(5):629-631.
[15] 卢建刚,彭清,范忠才.脂肪因子Chemerin的研究进展[J].山东医药,2012,52(11):96-98.
[16] EWA STELMANSKA, TOMASZ SLEDZINSKI, JACEK TURYN, et al.Chemerin gene expression is regulated by food restriction and food restriction-refeeding in rat adipose tissue but not in liver[J].Regulatory Peptides,2013,181(1):22-29.
[17] BANAS M, ZABIEGLO K, KASETTY G, et al. Correction:Chemerin Is an Antimicrobial Agent in Human Epidermis[J].Plos One,2013,8(3):e58709.
[18] TAKAHASHI M, OKIMURA Y, IGUCHI G, et al. Chemerin regulates beta-cell function in mice[J].Sci Rep,2011(1):123.
[19] GORALSKI KB,MCCARTHY TC,HANNIMAN EA,et al,Chemerin,a novel adipokine that regulates adipogenesis and adipocyte metabolism[J].J Biol Chem,2007,282(38):28175-28188.
[20] 韩晓菲,田慧.脂肪因子Chemerin 的研究进展[J].解放军医学杂志,2016,41(5):430-436.
[21] 陈旭.高胰岛素水平导致肥胖相关机制研究[D].郑州:郑州大学,2012.
[22] 陆维,王燕萍.体质指数对血压、血糖、血脂的影响.中华医院感染学杂志[J].2010,20(23):3715-3716.
[23] POLEDNAK AP.Trends in incidence rates for obesity-associated cancers in the US[J]. Cancer Detection and Prevention,2003,27(6):415-421.
[24] BOZAOGLU K,BOLTON K,MCMILLAN J,et al.Chemerin is a novel adipokine associated with obesity and metabolic syndrome[J].Endocrinology,2007,148(10):4687-4694.
[25] 余岚,马向华,沈捷,等.单纯性肥胖青少年Chemerin水平与代谢综合征的相关性[J].江苏医药,2011,37(12):1442-1445.
[26] RIMA CHAKAROUN,MATTHIAS RASCHPICHLER, NORA KLOTING,et al.Effects of weight loss and exercise on Chemerin serum concentrations and adipose tissue expression in human obesity[J].Metabolism-clinical&Experimental,2012,61(5):706-714.
[27] 刘燕.游泳运动及二甲双胍对肥胖大鼠血清chemerin水平的影响[J].安徽医科大学学报,2011,46(2):146-150.
[28] FATIMA SS,REHMAN R,BAIG M,et al. New roles of the multidimensional adipokine: Chemerin[J]. Peptides, 2014,62(2):15-20.
[29] TAKAHASHI M, TAKAHASHI Y, TAKAHASHIK, et al.Chemerin enhances insulin signaling and potentiates insulin-stimulated glucose uptake in 3T3-L1 adipocytes[J].FEBS Lett,2008,582(5):573-578.
[30] SELL H, LAURENCIKIENE J, TAUBE A, et al.Chemerin is a novel adipocyte-derived factor inducing insulin resistance in primary human skeletal muscle cells[J].Diabetes,2009,58(12):2731-2740.
[31] 郭启熠,吴从愿.肥胖致胰岛素抵抗的机制[J].国外医学内科学分册,2001,28(12):518-526.
[32] LLOYD J W, OWUSU P, SIEBURG M, et al.The good,the bad and the ugly? The role of exercise on obesity-induced Chemerin expression[J].The FASEB Journal,2012(26):869.
[33] ZABEL BA,AMANDA M,SILVERIO,et al.Butcher Chemokine-like receptor 1 expression and chemerin-directed chemotaxis distinguish palsmacytoid from myeloid dendritic cells in human blood[J].Immunol,2005,174(1):244-251.
[34] WEIGERT J.Systemic chemerin is related to inflammation rather than obesity in type 2 diabetes[J].Clin Endocrinol,2010,72(3):342-348.
[35] 李雪梅,瞿丽东.脂肪因子Chemerin在炎症和代谢综合征中的双重效应[J].中国组织工程研究,2015,19(7):1094-1098.
[36] DEMOR T,BRACKE KR,DUPOND LL,et al.The role of ChemR23 in the induction and resolution of igarette smoke-induced inflammation[J]. J Immunol.2011,186(9):5457-5467.
[37] BOYUK B,GUZEL EC,ATALAY H,et al.Relationship between plasma chemerin levels and disease severity in COPD patients[J].Clin Respiratory Journal ,2014,9(4):468-474.
[38] OZCAN E,SAYGUN NI,SERDAR MA,et al.Evaluation of the salivary levels of visfatin,chemerin,and progranulin in periodontal inflammation[J].Clin Oral Investig.2015,19(4):921-928.
[39] 姜勇,赵文华.成人肥胖的评价方法、指标及标准在公共卫生中应用的研究进展[J].卫生研究,2013,42(4):701-705.
[40] NACIFF JM,HESS K A,OVERMANN G J,et al.Gene expression changes induced in the testis by transplacental exposure to high and low doses of 17{alpha}-ethynyl estradiol,genistein,or bisphenol A[J].Toxicol Sci,2005,86(2):396-416.
[41] 刘一平,王有乐.肥胖与运动减肥研究进展[J].河北体育学院学报,2007,21(1):67-70.
[42] 杨新玲,李杰,卢素玉,等.瘦素-下丘脑神经肽Y轴在2型糖尿病大鼠发病过程中的变化[J].中国全科医学,2014,17(36):4329-4332.
[43] 张缨.有氧运动训练对大鼠下丘脑食欲调节肽的影响[J].中国运动医学杂志,2002,21(1):19-22.
[44] 柏友萍,张晶,江双双,等.减肥运动处方对超重肥胖大学生体脂、血糖与抵抗素的影响[J].卫生研究,2013,42(4):538-549.
[45] 李娟,唐东辉,陈巍.有氧运动结合抗阻训练对男性肥胖青少年心血管功能的改善及可能机制[J].体育科学,2013,33(8):37-42.
[46] 夏莉.二甲双弧和有氧运动对肥胖大鼠血清及不同组织Chemerin蛋白表达的影响[D].合肥:安徽医科大学,2011.
[47] 丁锐.Chemerin/CMKLR1-mRNA 在高脂膳食大鼠肝脏中的表达及有氧运动干预[D].苏州:苏州大学,2014.
[48] MEHRDAD FATHI,ALIREZA DELPASAND1,MORTEZA NASTARAN.Serum Chemerin and insulin sensitivity alterations due to exercise training below and above lactate thresholds in streptozocin-induced diabetic rats[J].Sport Sci Health,2015,11(2):211-215.
