户外环境条件下三角褐指藻生长和积累脂肪酸的条件优化
2018-03-01宋培钦刘鹭魏东
宋培钦,刘鹭,魏东
(华南理工大学食品科学与工程学院,广东广州 510640)
海洋微藻作为食物链的初级生产者,具有种类丰富、产油量较高及繁殖较快等特点,在海洋生态系统的能量流动及物质循环中扮演着重要的角色。海洋微藻含有大量高价值的天然活性物质,如抗生素、类胡萝卜素、虾青素、藻红蛋白,及不饱和脂肪酸EPA(二十碳五烯酸)、DHA(二十二碳六烯酸)等,在水产养殖业中,海洋微藻是鱼虾的重要饵料之一。近年来海洋微藻备受研究者的青睐,已被广泛应用于食品、医药、环境修复和生物能源等多个领域中。
三角褐指藻(Phaeodactylum tricornutum)是一种海洋真核单细胞藻,在分类上隶属于硅藻门、羽纹纲、褐指藻目、褐指藻科。通常情况下,三角褐指藻存在三种形态,梭形、卵形和三出放射形。在不同的条件下,三种形态间可以相互转换,在正常培养条件,多以三放出型和梭型为主[1]。三角褐指藻对环境的适应性较强,并且生长速率较快,易于室内或户外规模化培养。三角褐指藻脂肪酸含量非常丰富,其中EPA可占到总脂肪酸含量的30%以上[2]。近年来,大量研究表明,二十碳五烯酸(eicosapentaenoic acid,EPA)不仅具有抗单核细胞募集、抗炎、抗血栓、调节血脂水平和抗动脉粥样硬化等作用[3],对于乳腺癌的治疗方面也可达到显著的效果[4]。在水产养殖上,通常在动物的饵料中添加这些物质用来强化饵料,提高投喂动物的生长速度和存活率[5]。微藻养殖方法和反应器很多,如:跑道池、光生物反应器和发酵罐等。和其他培养模式相比,跑道池易于排氧及规模化,迄今为止90%的微藻商业化养殖,均采用跑道池[6]。本文选取三角褐指藻为实验对象,探讨在户外跑道池条件下,盐度、pH值和氮源种类对三角褐指藻生长情况及脂肪酸含量的影响,获得优化培养条件,以期为今后三角褐指藻的户外大规模培养提供理论依据。
1 材料与方法
1.1 藻种与培养方法
三角褐指藻(CCMP2561)由中国科学院水生生物研究所胡晗华研究员惠赠,藻种用f/2人工海水培养基进行斜面保种,保存温度4 ℃。
1.2 实验试剂及仪器
标准品十九碳饱和酸(C19:0,纯度>99%)购于美国Sigma公司;正己烷、甲醇为色谱纯试剂;二氯甲烷﹑磷酸二氢钠、硅酸钠等为分析纯试剂;Instant Ocean海盐购自Marineland;盐度计(ATC)购自上海淋誉贸易有限公司;笔式pH计(MT-8062)购自Exact Instrument;可见分光光度计(721)购自上海佑科仪器仪表有限公司;生物显微镜(PH100)购自中国江西凤凰光学集团公司;电子天平(BS224S)购自Sartorius;冷冻干燥机(Modulyod)购自Thermor Eletron Corporation;气相质谱联用仪(6890-5975i);高速离心机(TDL-4013)购自上海安享科学仪器厂;光照强度测定仪(1339R)购自TaiWan。
1.3 实验方法
1.3.1 培养条件

图1 开放池和超滤采收装置Fig.1 open pool and ultrafiltration recovery unit
三角褐指藻的实验室阶段培养主要包括培养基配制、扩种以及在实验室人工光源条件下连续培养。三角褐指藻种子液的培养分三步进行,首先在250 mL的锥形瓶培养至指数期,然后转移到3 L的锥形瓶,最后扩种到18 L的蓝盖瓶中培养。三角褐指藻的培养采用f/2人工海水培养基[7](表1和2)。海水利用人工海盐(Reef Crystals®,Instant Ocean Corp., VA, USA)进行配置。培养温度20±1 ℃,光照强度4000±500 Lux。接种前要对培养基和培养器皿进行高温高压灭菌和消毒处理。当培养至大桶时,需每天保持连续光照并且不断通入空气进行并混合搅拌。
三角褐指藻户外培养阶段主要包括由一级池到三级池的户外培养以及采收工作,一级培养池及采收装置见图1,(各级跑道池的底面积为:一级跑道2.5 m2;二级跑道池10 m2,三级跑道池40 m2,各级跑道池的培养水深均为20 cm)。在户外培养过程中每级跑道池的起始接种密度要达到OD680在0.2~0.3之间,接种时间一般选在早上光照不强的时间,接种以及培养过程中添加的培养基均采用f/2人工海水培养基。户外培养基用水为自来水,配置培养基之前经过0.5 μm微孔滤膜过滤系统进行过滤,并用10-5的二氧化氯消毒。在每次使用培养池之前,使用5×10-5的二氧化氯对培养池进行消毒,每次消毒时间12 h。
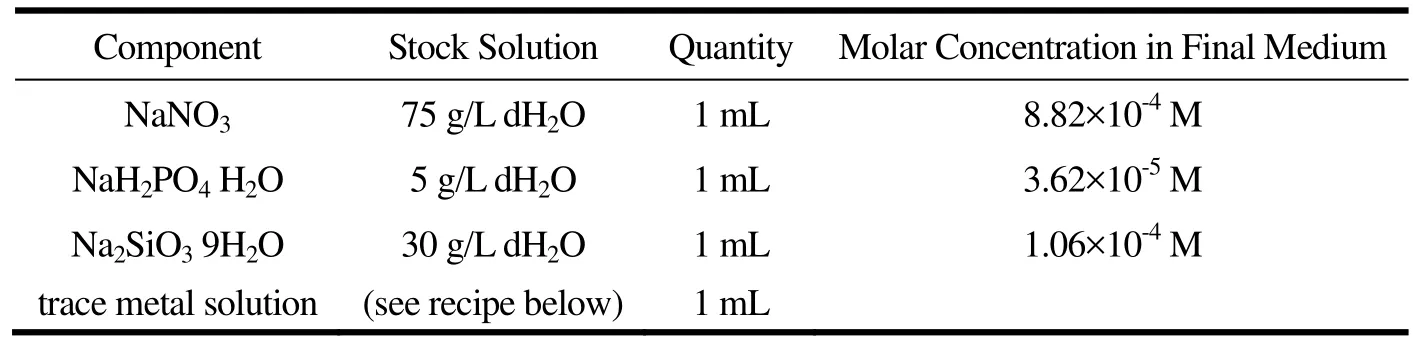
表1 f/2培养基Table 1 f/2 media
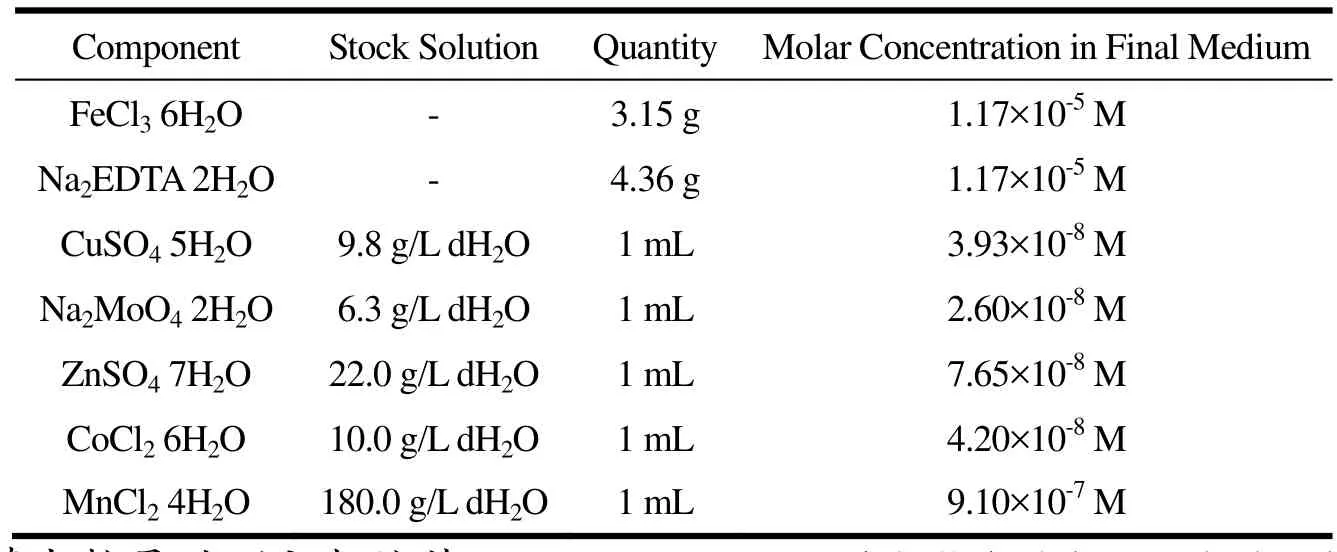
表2 f/2微量元素溶液Table 2 f/2 Trace element Solution
1.3.2 三角褐指藻生物量的测定与收获
三角褐指藻生物量的测定方法采用干重法和血球计数法。干重法:吸取藻液40 mL置于已准确称量的50 mL康宁离心管中,6000 r/min离心10 min,所得藻泥用超纯水进行清洗后继续进行离心,重复两次,得到的藻泥进行冷冻干燥,然后用分析天平称量干燥后藻粉的最终质量并计算差值。血球计数法:在生物显微镜下,通过血球计数板计数获得,采用25×16的血球计数板,计算公式如下:
细胞数/mL=(80小格内的细胞数/80)×400×1000×稀释倍数
采收过程采用二级过滤采收模式,首先藻液通过20 μm的预过滤装置除去藻液中较大的杂质,再经过0.5 μm的微滤膜组件来浓缩藻液,连续循环浓缩后达到预期浓度的藻液灌装后冷藏备用。
1.3.3 实验设计及处理
实验选用盐度、pH值和氮源种类为主要影响因子,浓度设计及氮源种类见表3。
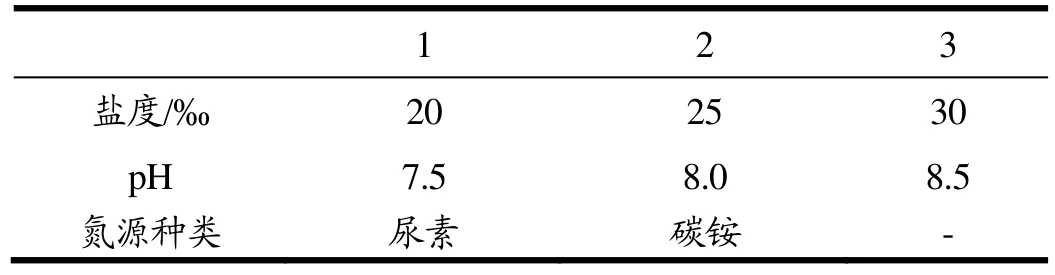
表3 影响因子实验设计Table 3 design of influence factor
盐度优化实验在一级跑道池中进行。培养基采用f/2人工海水培养基(氮、磷含量加倍);碳铵为氮源;pH值8.0(通过通入CO2控制pH恒定);设置A、B、C三池,盐度分别为20‰、25‰和30‰;使用电机搅拌,搅拌速率80 rad/min。保持接种密度相同。
pH优化实验在一级跑道池中进行。设置A、B、C三池,在接种密度相同的条件下,通过通入CO2分别控制培养基pH在7.5、8.0和8.5;培养基采用f/2人工海水培养基(氮、磷含量加倍);碳铵为氮源;选取最优盐度20‰;使用电机搅拌,搅拌速率80 rad/min。
氮源优化实验在一级跑道池中进行。设置A、B两池,采用f/2人工海水培养基(氮、磷含量加倍),分别以尿素和碳铵为氮源,氮源浓度为1.76 mmol/L;选取最优盐度20‰;最优pH值8.0;使用电机搅拌,搅拌速率80 rad/min。
实验过程中,通过跑道池﹑CO2储气罐、pH检测器和通气管道来维持pH值的恒定,通气管道的一端与储气罐相连,另一端伸入跑道池内的藻液中,检测器具有一根浸没于藻液中的pH电极,通过电极检测跑道池内的藻液pH值,然后将测得的pH与检测器所设定的阈值相比较,并形成阈值比较信号;通气管道上具有电子通气控制器能够控制通气管道的气路通断,电子通气控制器与检测器连接并可接收检测器的阈值比较信号,并根据所接收的阈值比较信号控制电子通气控制器,从而实现pH值的稳定。优化周期均为5 d,实验过程中,白天每隔两小时记录一次水温、pH值和光照强度,时刻注意其生长状况。实验结束后,通过对细胞生长速度、细胞密度、细胞干重以及脂肪酸的积累情况进行分析对比,以确定户外高密度培养三角褐指藻的最适宜条件。
1.4 脂肪酸含量的测定
准确称取10 mg藻粉于10 mL螺口试管中,加入100 μL内标液(1 g/L十九烷酸二氯甲烷溶液),加入2 mL质量浓度为40 g/L的KOH-CH3OH溶液,75 ℃水浴加热15 min,冷却至室温后加入2 mL BF3-CH3OH溶液(体积比1:4),再在75 ℃水浴中加热15 min,待冷却后加入1 mL饱和NaCl溶液和2 mL正己烷(色谱纯),涡旋1 min进行混匀,并在4000 r/min下离心8 min,取上清液1 mL过0.22 μm滤膜于进样瓶中。
在Angilent 6890N GC-MS上进行样品的脂肪酸含量测定,其中配备高效毛细管柱(DB-23,30 mm×0.25 mm,0.25 μm)和5975内置型质量探测器。载气为高纯氦气,流速为1 mL/min,样品分流比为2:1,进样量为0.2 μL,质谱的质量扫描范围为33~400 u。程序升温条件为:初始柱温110 ℃,保持2 min,以5 ℃/min升高至220 ℃保持5 min。各峰型的鉴定采用NIST05a谱库自动检索。根据各脂肪酸相对于C19:0内标的峰面积,计算出各脂肪酸组分的绝对含量。
1.5 数据统计
采用Microcal Origin 8.5Software对数据进行处理和统计,实验数据均采用平均值±标准误差(mean±SD)给出(n=3)。采用t检验来确定各试验组之间差异,p<0.05为显著差异。
2 结果与讨论
2.1 培养基盐度对三角褐指藻户外生长情况及脂肪酸含量的影响
盐度优化实验在一级跑道池中进行,培养期间白天平均最高光强为12000 Lux,最低光强为1500 Lux;跑道池藻液温度最高为25 ℃,最低为22 ℃。如图2所示,随着培养时间的变化,细胞密度和干重呈现出不断增长的趋势。随着培养液盐度的增加,细胞密度及干重的增长有显著性差异(p<0.05)。实验进行到第三天时,细胞进入对数生长期,细胞密度增长较快。在第5 d时,20‰盐度下细胞密度及干重最大,分别达到6.01×106/mL,177.78 mg/L;25‰盐度次之,细胞密度为3.85×106/mL,干重为165.93 mg/L;30‰盐度下最低,细胞密度为3.6×106/mL,干重为158.52 mg/L。
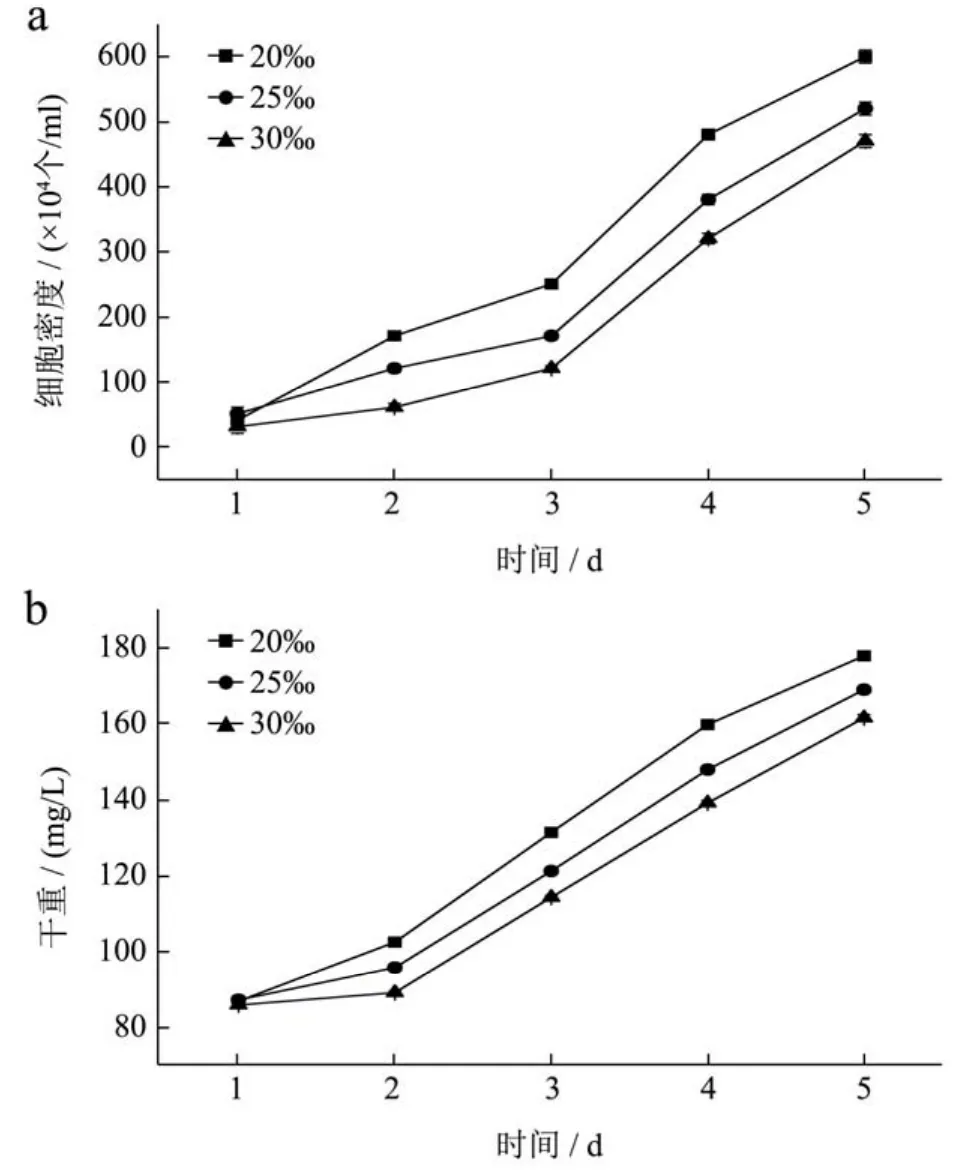
图2 不同盐浓度下细胞密度(a)、生物量浓度(b)的变化Fig.2 Cell concentration(a) and dry biomass concentration (b)of Phaeodactylum trirornutum at different salinities
如图3-a所示,总脂肪酸含量随着培养时间的增加呈现出不断增长的趋势。在培养初期,随着盐度的升高,总脂含量呈现出下降的趋势。在培养至第2 d时,20‰盐度下的总脂含量最大,且不同盐度条件下,总脂肪酸含量有显著性差异(p<0.05)。培养至第5 d时,不同盐度条件下,总脂肪酸含量无显著性差异,在30‰盐度条件下总脂肪酸含量达到最高,为123.99 mg/g。如图3-b所示,在培养初期,随着盐度的升高,EPA所占总脂肪酸比例呈现出先升高后下降的趋势。培养至第5 d时,不同盐度条件下,总脂肪酸组成中EPA含量无显著性差异,分别为17.78%、17.33%和17.09%。
三角褐指藻对盐度的适应范围较广,在9‰~92‰的范围内都能生存,较高和较低浓度下,都不利于三角褐指藻的生长[8]。盐度在一定程度上影响着藻类的渗透压、营养盐的吸收及其悬浮性。当藻细胞所处的海水盐度发生改变时,细胞的渗透压也随之改变,过高或过低的海水盐度都会对藻细胞造成伤害。陈峰等[9]研究报道称脂类对微藻细胞膜的通透性起关键作用,因此盐度的变化能影响许多微藻的脂质含量,种类和组成比例。叶丽等[10]研究表明三角褐指藻紫外诱变株MP-2在盐度范围为10~40‰下生长良好,最佳生长盐度为25‰,高盐条件下(盐度高于35),该藻的生长受到明显的抑制;但高盐有利于该藻总脂的积累。可能由于藻种及培养环境不同,本研究中三角褐指藻最适宜生长盐度20‰。并且在30‰盐度条件,总脂含量相对较高。Qiao H[8]等研究称EPA含量随着盐度的变化,没有显著性差异这与本实验研究的结果相符合。

图3 不同盐度下脂肪酸总量(a)及EPA占总脂肪酸百分比(b)Fig.3 Total fatty acid content (a) and EPA percentage of TFA (b)of Phaeodactylum tricornutum at different salinities
综上所述,盐度为20‰不仅有利于三角褐指藻的生长,更适合于脂肪酸的积累,并且总脂肪酸组成中EPA所占的比例也相对较高。
2.2 培养基pH值对三角褐指藻户外生长情况及脂肪酸含量的影响
pH值优化实验在一级跑道池中进行,培养期间白天平均最高光强为35000 Lux,最低光强为2000 Lux;培养基水温最高为28 ℃,最低为21 ℃。
从图4中可以看出,在起始两天内,不同pH条件下,细胞密度以及干重均呈现增长的趋势。从实验第3 d开始,三种pH值下的细胞密度及干重的增长速率发生了明显的变化,pH值8.0下的细胞密度和干重终细胞密度高达5.8×106/mL,干重为166.39 mg/L,pH值8.5条件下的细胞密度及干重量相对增长缓慢,最终达到4.15×106/mL,干重为114.72 mg/L,此时而pH值为7.5条件下的细胞密度及干重则开始下降,最终降为1.75×106/mL,干重下降到66.11 mg/L。通过图5-a可以看出,pH值8.0以及pH值8.5条件下,随着培养时间的增加,总脂肪酸含量升高。三角褐指藻培养结束后,三种pH条件下,总脂肪酸含量差异较为显著(p<0.05),pH值为8.0时三角褐指藻细胞的脂肪酸含量最高为145.92 mg/g;pH值为8.5的稍低,为134.92 mg/g;pH值7.5的脂肪酸含量最低,为109.76 mg/g。如图5-b所示,不同pH条件下,EPA占总脂肪酸的百分比也有显著性差异。培养至第五天时,pH为8.5时,EPA占总脂肪酸百分比最大,为20.19%。
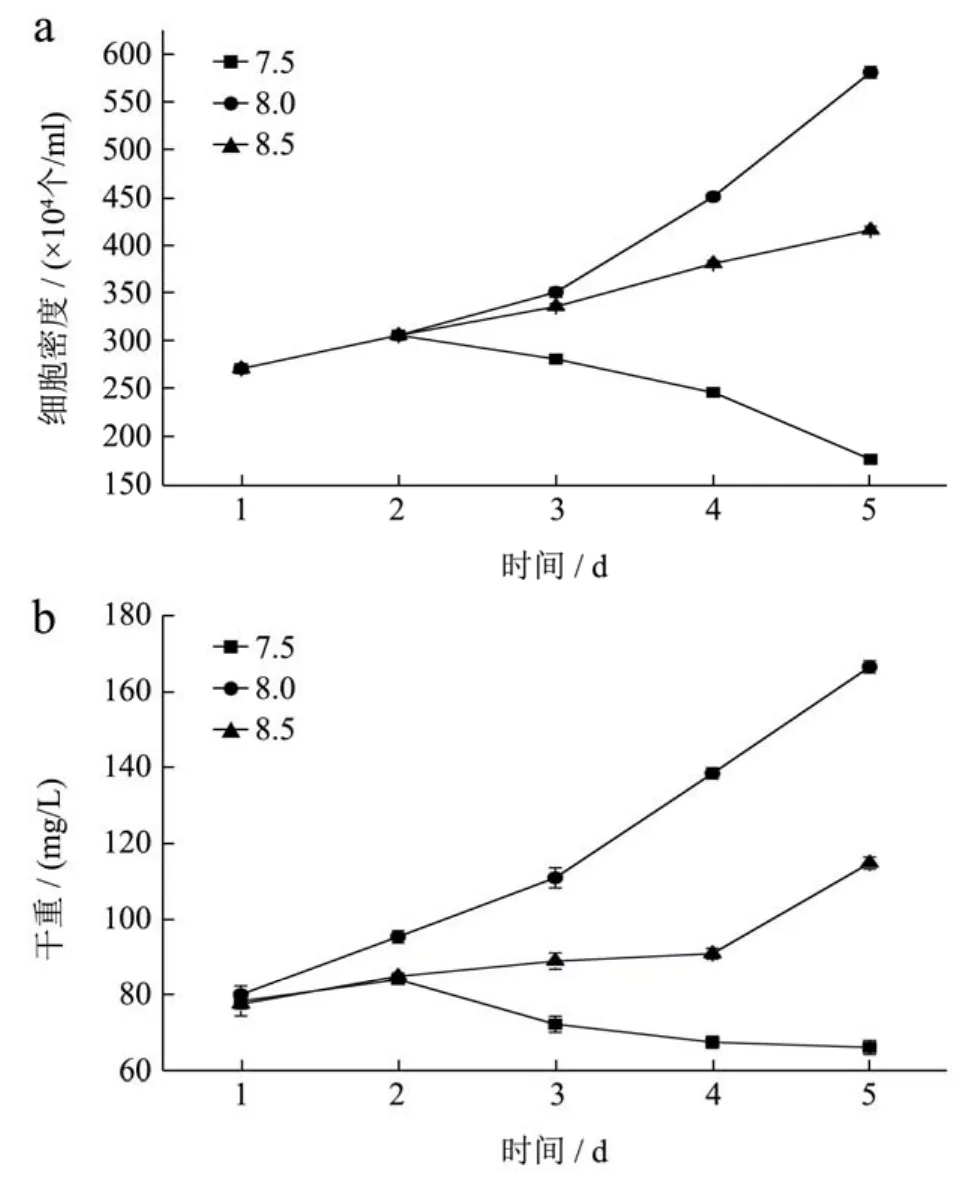
图4 不同pH条件下细胞密度(a)、生物量浓度(b)随时间的变化Fig.4 Cell concentration(a) and dry biomass concentration (b)of Phaeodactylum trirornutum at different pH conditions
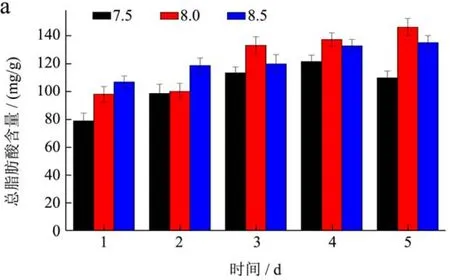

图5 不同pH条件下脂肪酸总量(a)及EPA占总脂肪酸百分比(b)Fig.5 Total fatty acid content (a) and EPA percentage of TFA (b)of Phaeodactylum tricornutum at different pH conditions
海洋微藻的最适pH值与海水相近约为8.0,但也具有藻种差异性。陈峰等[11]研究指出pH值的变化会影响微藻细胞内外离子平衡、相关膜的结构组成、渗透压及藻细胞内相关酶的活性,一旦偏离最适pH值,微藻的生长及体内相关代谢活动会受抑制。吴伟伟等[12]研究发现三角褐指藻的最适pH值是8.5,且与pH值7.5和8.0的差异性显著;此外pH值在6.0~10.0条件下总脂含量呈现出先增加后下降的趋势,在8.0条件下达到最大值;EPA占总脂肪酸的比例在7.5和8.5条件下相对较高,较8.0条件下差异性显著,在8.5条件下达到最大值。Qiao H等[8]研究发现随着培养时间的增加单不饱和脂肪酸占总脂肪的比例逐渐下降,EPA占总脂肪酸的百分比呈现出先升高后下降的趋势,在指数期时增加,在稳定期时开始下降。由于藻种的差异性及培养环境的不同,本实验中三角褐指藻的最适生长pH值为8.0,并且在7.5条件下生长受到抑制;总脂含量及EPA占总脂肪酸的比例均与之前的研究呈现出相同的变化规律。
由此可以看出,pH值为8.0条件下更适合于三角褐指藻的生长;pH值为8.5的条件下更有利于脂肪酸的积累,并且总脂肪酸组成中EPA所占的比例也相对较高。
2.3 培养基中氮源种类对三角褐指藻户外生长情况及脂肪酸含量的影响
氮源种类优化实验在一级跑道池中进行,培养期间白天平均最高光强为30000 Lux,最低光强为2000 Lux;培养基水温最高为29 ℃,最低为23 ℃。不同氮源条件下,起始接种细胞密度分别为尿素1.9×106/mL、碳酸氢铵1.8×106/mL。
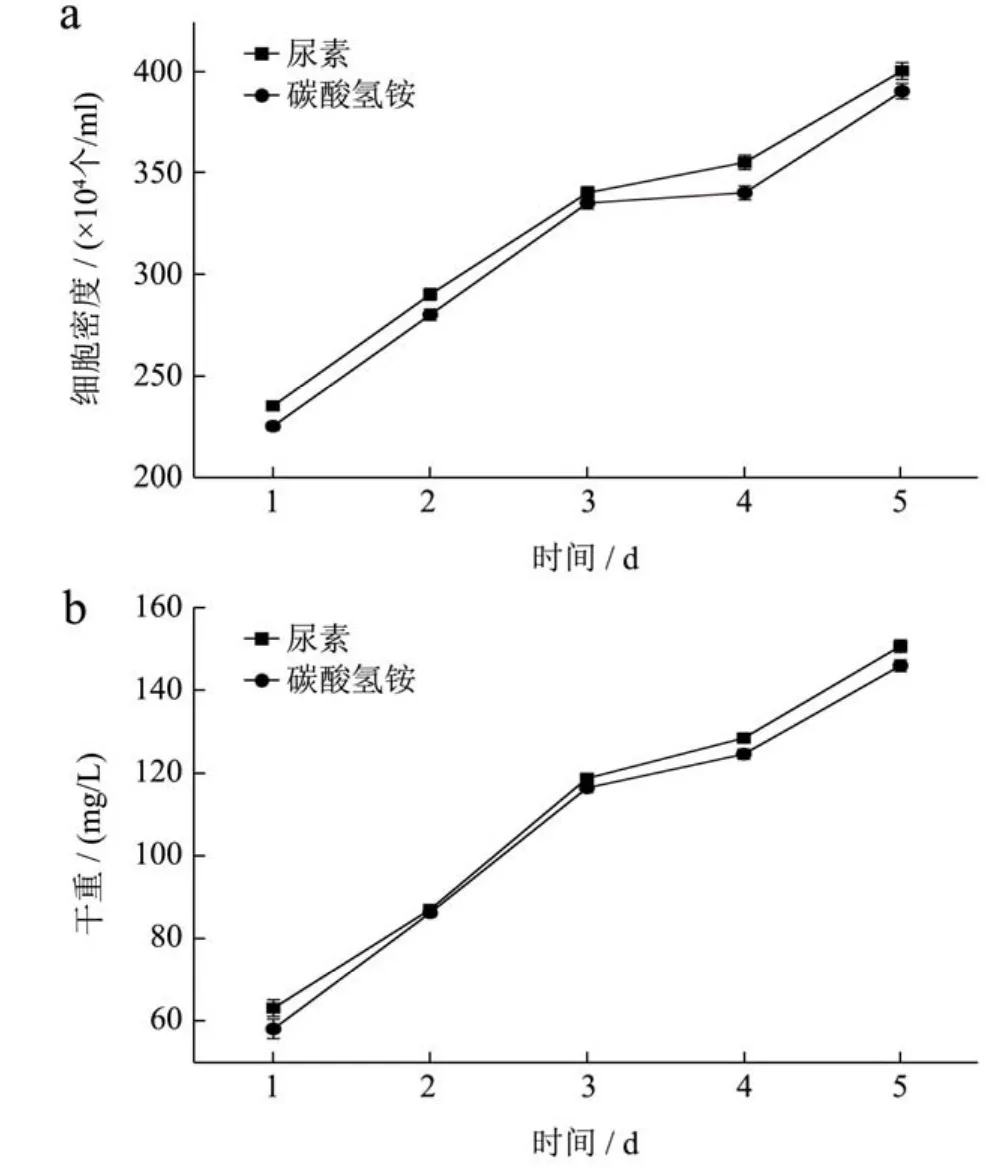
图6 不同氮源条件下细胞密度(a)、生物量浓度(b)随时间的变化Fig.6 Cell concentration(a) and dry biomass concentration (b)of Phaeodactylum trirornutum with different nitrogen sources

图7 不同氮源条件下脂肪酸总量(a)及EPA占总脂肪酸百分比(b)Fig.7 Total fatty acid content (a) and EPA percentage of TFA (b)in Phaeodactylum tricornutum with different nitrogen sources
培养结束后,最终细胞密度分别为4.0×106/mL和3.9×106/mL,生物量分别达到150.83 mg/L和146.11 mg/L。以尿素和碳酸氢铵这两种氮源培养三角褐指藻时,对其生物量的影响差异性显著(p<0.05)。
通过图7-a可以看出,培养结束后,当使用碳酸氢铵作为氮源时三角褐指藻细胞的脂肪酸含量最高为169.72 mg/g,尿素作为氮源时脂肪酸含量则相对较低,为109.76 mg/g。但通过图7-b可以看出,不同氮源条件下,总脂肪酸组成中EPA的含量均呈现出先升高后下降的趋势。在第二天时达到最大值,且不同氮源间无显著性差异;培养至第5 d时,尿素作为氮源其EPA的百分比要显著性高于碳铵,占总脂肪酸的20.43%,碳铵为氮源时,其EPA含量相对较低,为15.58%。
氮是微藻生长所必需的基本元素之一,同时也是构成藻体内蛋白质、核酸及色素的重要组成元素,对藻类的生长发育有着重要的作用。蒋汉明等[13]以不同种类氮源(NH4+、CO(NH2)2、NO3-)、不同氮源浓度(0、0.9、1.8、3.5、7.0 mmol/L)培养三角褐指藻,得出氮源浓度低于1.8 mmol/L时,以NH4+为氮源时比生长速率最高,CO(NH2)2次之;氮浓度高于3.5 mmol/L时,以CO(NH2)2为氮源比生长速率最高,NH4+次之;在氮浓度低于3.5 mmol/L时,以NO3-和CO(NH2)2为氮源时EPA占总脂肪酸的比例相对较高,较NH4+为氮源时差异性显著。张文源等[14]以含氮量为14.36mmol/L的硝酸钠、尿素、碳铵为氮源在不同培养基中培养三角褐指藻,得出尿素较适宜三角褐指藻的生长,且更有利于EPA的积累。Andreas[15]等以KNO3、CO(NH2)2和NH4Cl为氮源培养三角褐指藻,得出尿素为最佳氮源且更有利于EPA的积累。本实验以含氮量1.76 mmol/L的尿素﹑碳铵为氮源,得出尿素较适宜三角褐指藻的生长,且更有利于EPA的积累,但以碳铵为氮源时,更有利于脂肪酸的积累。这与大多数研究结果相符合。由此可以看出,以尿素为氮源时可以明显提高三角褐指藻的生物量及总脂肪酸组成中EPA的所占的比例,以碳铵作为氮源时则更有利于三角褐指藻脂肪酸的积累。
2.4 光暗周期对三角褐指藻脂肪酸组成和含量的影响
通过图8可以看出,一天中不同时刻总脂肪酸含量,及EPA占总脂肪酸的百分比均有较大差异。在上午10时,三角褐指藻中所积累的脂肪酸含量是最高的,为136.15 mg/g。而EPA占总脂肪酸含量的百分比最高的时刻为4时~7时,占总脂肪酸的27.3%以上,10时处EPA所占百分比也相对较高,占总脂肪酸的26.91%。

图8 总脂肪酸含量(a)及EPA占总脂肪酸含量百分比(b)的昼夜变化规律Fig.8 Diurnal variation of total fatty acid content (a) and EPApercentage of TFA (b) in Phaeodactylum tricornutum
由此可以看出,三角褐指藻积累脂肪酸的最佳时间为上午10时左右,并且此时EPA占总脂肪酸的百分比也相对较高,所以上午10时是三角褐指藻户外培养的最佳采收时间。
3 结论
本研究通过优化影响户外跑道池中三角褐指藻生长及脂肪酸含量的环境因素,得到户外培养三角褐指藻的最适条件,并建立了一整套完整的户外培养、采收设备及原位补碳-pH在线控制专利技术。在户外跑道池规模化培养三角褐指藻时,选择20‰盐度、pH值8.0条件下,以尿素为氮源进行培养,不仅可以获得较大的生物量,同时可以得到含量较高的脂肪酸,且更有利于EPA的积累。此外,藻种采集的时间可以选择在上午10时左右进行,以获得最大生物量和脂肪酸含量。在今后的研究中,多个因素之间的交互作用,及交互作用下的最优水平,仍需要更深入的探究。
[1] Kim S M, Jung Y-J, Kwon O-N, et al. A potential commercial source of fucoxanthin extracted from the microalga Phaeodactylum tricornutum [J]. Applied Biochemistry and Biotechnology, 2012, 166(7): 1843-55
[2] Santos-S Nchez N, Valadez-Blanco R, Hern Ndez-Carlos B,et al. Lipids rich in ω-3 polyunsaturated fatty acids from microalgae [J]. Applied Microbiology And Biotechnology,2016, 100(20): 8667-84
[3] Mcmanus S, Aacuten, Tejera N, et al. Differential effects of EPA vs. DHA on postprandial vascular function and the plasma oxylipin profile in men [J]. Journal of Lipid Research,2016, 57(9): 1720-1727
[4] Rendeiro C, Sheriff A, Bhattacharya T K, et al. Long-lasting impairments in adult neurogenesis, spatial learning and memory from a standard chemotherapy regimen used to treat breast cancer [J]. Behavioural Brain Research, 2016, 315:10-22
[5] Wu H, Li T, Wang G, et al. A comparative analysis of fatty acid composition and fucoxanthin content in six Phaeodactylum tricornutum strains from different origins [J].Chinese Journal of Oceanology and Limnology, 2016, 34(2):1-8
[6] Norsker N-H, Barbosa M J, Vermu M H, et al. Microalgal production-a close look at the economics [J]. Biotechnology Advances, 2011, 29(1): 24-7
[7] Gullardr R. Culture of phytoplankton for feeding marine invertebrates [M]. Culture of marine invertebrate animals.Springer, 1975
[8] Qiao H, Cong C, Sun C, et al. Effect of culture conditions on growth, fatty acid composition and DHA/EPA ratio of Phaeodactylum tricornutum [J]. Aquaculture, 2016, 452:311-7
[9] Chen G Q, Jiang Y, Chen F. Salt-Induced alterations in lipid composition of diatom Nitzschia Laevis (Bacillariophyceae)under heterotrophic culture condition(1) [J]. Journal of Phycology, 2008, 44(5): 1309-14
[10] 叶丽,蒋霞敏,毛欣欣,等.温,光,盐对三角褐指藻紫外诱变株生长,总脂及脂肪酸的影响[J].生态学杂志,2015, 34(2):454-62
YE Li, JIANG Xia-min, MAO Xin-xin, et al. Effects of temperature, light intensity and salinity on the growth, total lipid and fatty acid of Phaeodactylum tricornutum mutant [J].Chinese Journal of Ecology, 2015, 34(2): 454-62
[11] 陈峰,姜悦.微藻生物技术[M].北京:中国轻工业出版社,1999
CHEN Feng, JIANG Yue. Thematic technology of microalgae [M]. Beijing: China Light Industry Press, 1999
[12] 吴伟伟.环境因子和抑制剂对三角褐指藻脂肪酸合成的影响[D].南京:南京农业大学,2012
WU Wei-wei. Effects of environmental factors and inhibitors on fatty acid synthesis of Phaeodactylum tricornutum [D].Nanjing: Nanjing Agricultural University, 2012
[13] 蒋汉明,高坤山.氮源及其浓度对三角褐指藻生长和脂肪酸组成的影响[J].水生生物学报,2004,28(5):545-51
JIANG Han-ming, GAO Kun-shan. Effects of nitrogen source and its concentration on growth and fatty acid composition of Phaeodactylum tricornutum [J]. Acta Hydrobiologica Sinica, 2004, 28(5): 545-51
[14] 张文源,高保燕,李爱芬,等.不同培养条件对三角褐指藻生长及其生物活性成分积累的影响[J].海洋科学,2016,40(5):57-65
ZHANG Wen-yuan, GAO Bao-yan, LI Ai-fen, et al. Effects of different culture conditions on the growth and bioactivity constituents of phaeodactylum tricornutum [J]. Marine Sciences, 2016, 40(5): 57-65
[15] Meiser A, Schmid-Staiger U, Trösch W. Optimization of eicosapentaenoic acid production by Phaeodactylum tricornutum in the flat panel airlift (FPA) reactor [J]. Journal of Applied Phycology, 2004, 16(3): 215-225
