食管鳞癌细胞中LSD1通过与Notch靶基因启动子结合调控Notch通路相关蛋白表达*
2017-12-06张幸丽侯桂琴刘襄艳白一汝鲁照明
张幸丽,侯桂琴,凡 丞,刘襄艳,高 盼,田 菲,白一汝,赵 琦,鲁照明
食管鳞癌细胞中LSD1通过与Notch靶基因启动子结合调控Notch通路相关蛋白表达*
张幸丽,侯桂琴,凡 丞,刘襄艳,高 盼,田 菲,白一汝,赵 琦,鲁照明#
食管鳞癌;LSD1;Notch;Hes1
目的:探讨食管鳞癌细胞中LSD1对Notch信号通路的调控作用及调控机制。方法Western blot法检测5种食管鳞癌细胞KYSE450、KYSE790、Eca109、EC9706和TE-1中LSD1及Notch通路相关蛋白的表达情况。用LSD1抑制剂TCP 处理KYSE450、KYSE790和Eca109细胞或用LSD1 siRNA处理Eca109细胞,Western blot检测处理前后细胞中LSD1、组蛋白H3K4me2和Notch信号通路相关蛋白的表达情况。结果5种食管鳞癌细胞中均存在LSD1及Notch信号通路相关蛋白的表达,且表达水平不同;TCP处理后,与未处理组相比,KYSE450、KYSE790和Eca109细胞中LSD1的表达水平均降低,组蛋白H3K4me2表达升高,Notch信号通路相关蛋白的表达均降低(P<0.05);LSD1 siRNA同样使Eca109细胞中LSD1与Notch信号通路相关蛋白表达受到抑制(P<0.05)。结论食管鳞癌细胞中LSD1对Notch信号通路有调节作用,且LSD1可能是通过与Notch靶基因Hes1的启动子区域结合实现的。
食管癌是常见的消化道肿瘤之一,根据其病理学特征分为食管鳞癌和食管腺癌两种类型[1]。在中国常见的是食管鳞癌,其发病率在河南省最高。其侵袭性强、预后差、多伴有淋巴结转移且易复发的特点促使人们从分子水平上了解食管鳞癌的发生机制,以寻找有效的分子治疗靶点。赖氨酸特异性去甲基化酶1(lysine specific demethylase 1,LSD1)是一个黄素腺嘌呤二核苷酸依赖性胺氧化酶,能够特异性去除组蛋白H3第4位赖氨酸(H3K4)和第9位赖氨酸(H3K9)上的单双甲基基团[2]。研究[3]表明,LSD1参与调控肿瘤细胞增殖、迁移和侵袭等,并且在多种肿瘤的发生及发展中发挥重要作用。Notch信号通路是通过细胞间相互作用来调节生物体生长发育的一个十分保守的信号通路[4]。Notch信号的产生是通过相邻细胞的Notch配体与受体相互作用,Notch蛋白经过3次剪切,由胞内区(NICD)释放进入细胞质,并进入细胞核与转录因子CSL结合,形成NICD/CSL转录激活复合体,从而调控下游Hes1、DTX1等靶蛋白发挥生物学作用。Notch的异常表达常与肿瘤细胞的侵袭、转移和凋亡等过程密切相关[5]。该研究从分子水平探讨食管鳞癌中LSD1对Notch信号通路的调控作用及调控机制,为进一步了解食管鳞癌发生的分子机制提供实验基础和理论依据。
1 材料与方法
1.1细胞株与材料食管鳞癌细胞株Eca109、EC9706和TE-1购自上海中科院细胞库,KYSE450、KYSE790由郑州大学第一附属医院病理科李晟磊博士惠赠;RPMI 1640培养基与胎牛血清均购自以色列BI公司,LSD1抑制剂反苯环丙胺硫酸盐(TCP)购自美国Medchemexpress公司,LSD1 siRNA和siRNA-Mate均购自上海吉玛公司,抗体LSD1[普通和染色质免疫沉淀(ChIP)级别]和Hes1均购自美国Abcam公司,Notch3、Notch1、H3K4me2购自美国CST公司,DTX1购自美国GeneTex公司, ChIP检测试剂盒购自德国Millipore公司,real-time PCR试剂盒购自美国Roche公司,real-time PCR引物均由上海生工生物工程技术服务有限公司合成。内参GAPDH引物:上游5’-TACTAGCGGTTTTACGGGCG-3’,下游5’-TCGAA CAGGAGGAGCAGAGAGCGA-3’。Hes1基因不同区域的扩增引物共4对。引物1:上游5’-AGGTCACCCA GAGTCAGGAA-3’,下游5’-CAAGCGTCTTGTTTGAT GTG-3’;引物2:上游5’-CGTGTCTCCTCCTCCCATT-3’,下游5’-GAGAGGTAGACGGGGGATTC-3’;引物3:上游5’-TCAACACAGCACCGGATAAA-3’,下游5’-TCAGCT GGCTCAGACTTTCA-3’;引物4:上游5’-GGCTTTTG GTGGAATTTGAA-3’,下游5’-TCATGGAGGATTGGT GAAAAG-3’。
1.2细胞培养食管癌细胞株均用含体积分数10%胎牛血清、100 U/mL青霉素、100 mg/L链霉素的RPMI 1640培养基,于37 ℃、体积分数5%CO2培养箱中培养,取对数生长期的细胞用于实验。
1.3不同处理方式食管鳞癌细胞中LSD1和Notch信号通路相关蛋白的Westernblot检测收集培养至对数生长期的5种食管鳞癌细胞,提取细胞总蛋白;并且分别用0、10、50 μmol/L TCP处理KYSE450、KYSE790和Eca109细胞48 h,提取细胞总蛋白和组蛋白,Western blot法检测5种食管鳞癌细胞中LSD1与Notch信号通路相关蛋白的表达情况及TCP对上述各蛋白及组蛋白的影响。首先将蛋白用SDS-PAGE分离胶分离并湿转至硝酸纤维素膜,将膜用50 g/L脱脂牛奶室温封闭2 h后,分别与一抗(LSD1、Notch1、DTX1、Hes1 和H3K4me2均按11 000稀释)或β-actin、H3 内参抗体4 ℃孵育过夜,PBST洗涤后再用相应的二抗室温孵育2 h,PBST洗涤后加ECL发光液孵育1 min,暗室显影曝光。实验重复3次,用Image J软件对条带进行灰度分析。
1.4LSD1siRNA对Eca109细胞中Notch信号通路相关蛋白表达的影响Eca109细胞以1.5×105个/孔接种至6孔板,培养至融合度达30%~50%时,用LSD1 siRNA转染细胞,具体步骤如下:将LSD1 siRNA或阴性对照siRNA溶液加入200 μL无血清培养基中,使其终浓度为150 nmol/L,轻轻混匀后室温放置5 min,然后加入5 μL siRNA-Mate充分混匀,室温放置10 min后逐滴加入细胞培养孔,轻轻混匀后继续培养,6 h后荧光拍照,72 h后收集细胞并提取总蛋白,Western blot检测LSD1与Notch3、Notch1、DTX1和Hes1的表达。实验重复3次。
1.5ChIP法检测Eca109细胞中LSD1与Hes1基因启动子结合情况用100 nmol/L的TCP处理Eca109细胞,48 h后采用ChIP法检测LSD1与Hes1基因启动子结合情况。主要步骤如下。①用体积分数1%的甲醛溶液对细胞中蛋白-DNA进行交联,然后用SDS裂解缓冲液裂解细胞。②用细胞超声破碎仪超声断裂细胞染色质,然后4 ℃离心取上清。③免疫共沉淀。上清中加入蛋白G-琼脂糖小珠,4 ℃振摇1 h后离心,取体积分数1%蛋白裂解液上清作为对照,在剩余的上清中分别加入免疫共沉淀抗体,包括阳性对照抗体、阴性对照抗体和LSD1抗体(ChIP级别),4 ℃摇床孵育过夜后,向溶液中加入蛋白G-琼脂糖小珠以吸附免疫沉淀复合物,离心去上清,依次用低盐、高盐、氯化锂和TE缓冲液清洗蛋白G-琼脂糖小珠。④用洗脱缓冲液洗脱免疫沉淀复合物,同时在装有对照蛋白裂解液的管中加入相同体积的洗脱缓冲液。⑤依次用5 mol/L NaCl、RNase A、0.5 mol/L EDTA、1 mol/L Tis-HCl和蛋白酶K处理上述洗脱缓冲液,以使蛋白-DNA结合物解交联,并用DNA纯化柱回收DNA。⑥real-time PCR分析LSD1蛋白与Notch靶基因Hes1启动子区域的结合情况。PCR反应体系:10 μL Master Mix,上下游引物各1 μL,DNA模板2 μL,用灭菌双蒸水补足20 μL。用三步法PCR反应程序(预变性:95 ℃ 10 min;PCR反应:95 ℃ 10 s,60 ℃ 10 s,72 ℃ 10 s,共45个循环;溶解:95 ℃ 10 s,65 ℃ 60 s,97 ℃ 1 s)进行扩增,扩增结果用ΔΔCt值进行相对定量,分别计算Eca109细胞中Hes1基因不同结合区域扩增片段的富集效率(代表特异性抗体所沉淀的目的蛋白水平),分析LSD1与Hes1基因启动子不同区域的结合情况。实验重复3次。
1.6统计学处理采用SPSS 21.0进行数据分析。多组间LSD1及Notch信号通路相关蛋白表达的比较均采用单因素方差分析,两两比较采用LSD-t检验;阴性对照组和LSD1 siRNA组LSD1及Notch信号通路相关蛋白表达的比较,TCP处理前后Eca109细胞中Hes1基因不同结合区域扩增片段的富集效率的比较,均采用两独立样本的t检验。检验水准α=0.05。
2 结果
2.1 5种食管鳞癌细胞中LSD1与Notch信号通路相关蛋白的表达情况结果见图1。由图1可以看出,Notch信号通路相关蛋白在不同食管鳞癌细胞中表达不同,LSD1在5种食管鳞癌细胞中表达均较高,尤其在Eca109和EC9706细胞中表达最高。

1~5:分别为KYSE450、KYSE790、Eca109、EC9706、TE-1细胞。图1 5种食管鳞癌细胞中LSD1与Notch信号通路相关蛋白的表达情况
2.2TCP对KYSE450、KYSE790和Eca109细胞中LSD1、H3K4me2和Notch信号通路相关蛋白表达的影响结果见图2、3和表1~4。可知,不同浓度TCP处理3种细胞后,LSD1的表达均不同程度降低,组蛋白H3K4me2表达升高,Notch信号通路相关蛋白DTX1、Hes1的表达受到抑制。
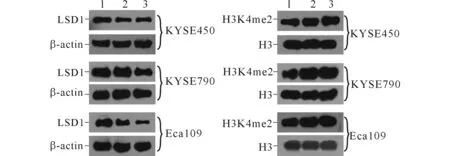
1~3:分别为0、10、50 μmol/L TCP处理后细胞。图2 TCP对食管鳞癌细胞中LSD1与组蛋白H3K4me2表达的影响
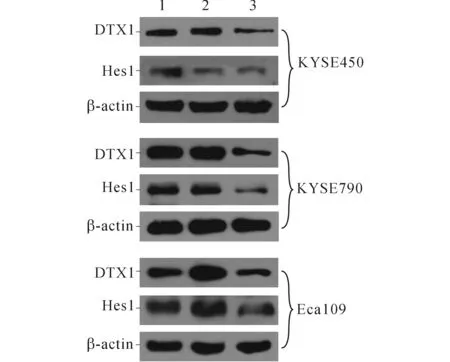
1~3:分别为0、10、50 μmol/L TCP处理后细胞。图3 TCP对食管鳞癌细胞中Notch信号通路相关蛋白表达的影响

c(TCP)/(μmol·L-1)nKYSE450KYSE790Eca109031.457±0.0811.328±0.1021.278±0.0531030.689±0.140∗0.956±0.0310.816±0.0825030.601±0.034∗0.631±0.152∗0.657±0.093∗F13.33734.95412.770P0.006<0.0010.007
*:与0 μmol/L TCP组比较,P<0.05。
*:与0 μmol/L TCP组比较,P<0.05。
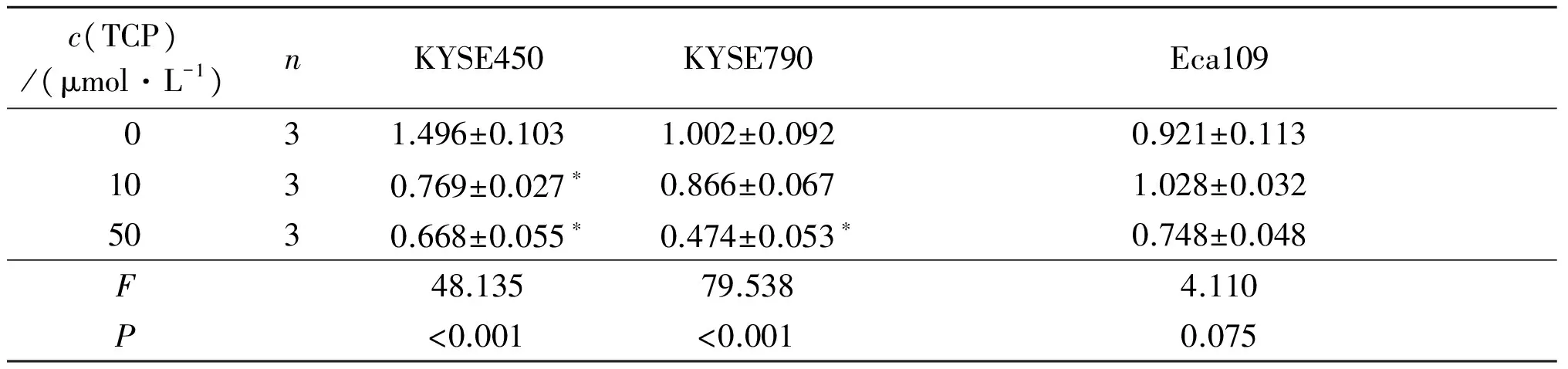
表3 不同浓度TCP对KYSE450、KYSE790和Eca109细胞中DTX1蛋白表达的影响
*:与0 μmol/L TCP组比较,P<0.05。
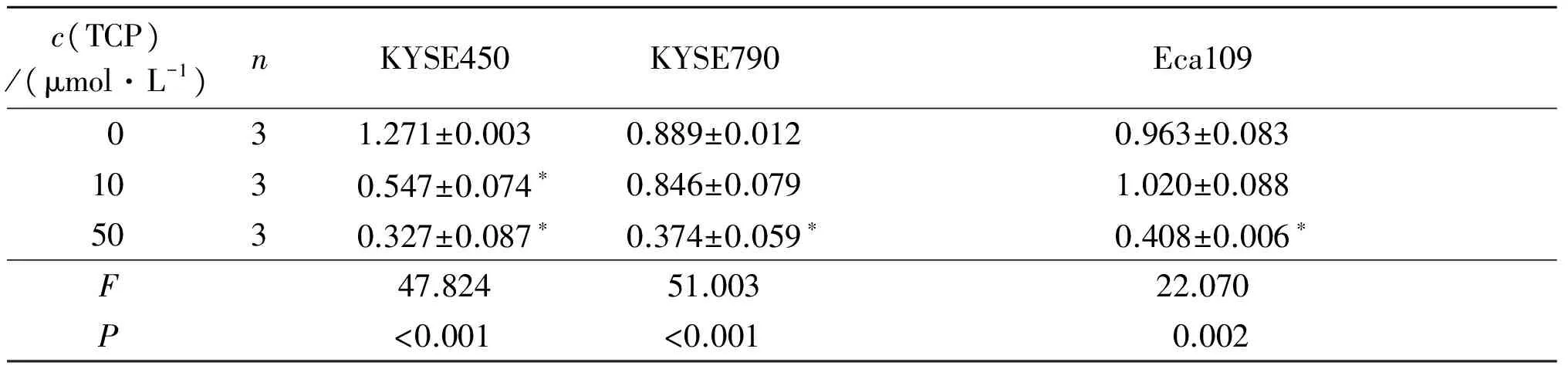
表4 不同浓度TCP对KYSE450、KYSE790和Eca109细胞中Hes1蛋白表达的影响
*:与0 μmol/L TCP组比较,P<0.05。
2.3LSD1siRNA对Eca109细胞中Notch信号通路相关蛋白表达的影响结果(图4、表5)显示,与阴性对照组相比,LSD1 siRNA组细胞中LSD1表达降低,而Notch信号通路相关蛋白Notch3、Notch1、DTX1、Hes1表达也降低。

1:阴性对照组;2:LSD1 siRNA组。图4 LSD1 siRNA对Eca109细胞中LSD1与Notch信号通路相关蛋白表达的影响
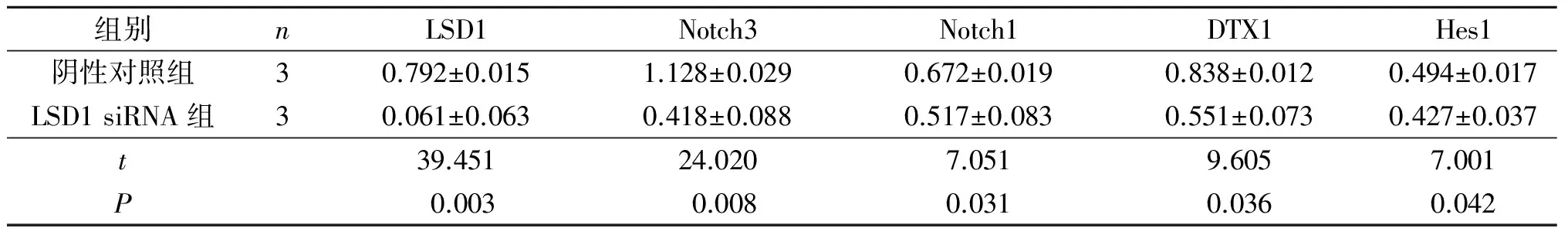
组别nLSD1Notch3Notch1DTX1Hes1阴性对照组30.792±0.0151.128±0.0290.672±0.0190.838±0.0120.494±0.017LSD1siRNA组30.061±0.0630.418±0.0880.517±0.0830.551±0.0730.427±0.037t39.45124.0207.0519.6057.001P0.0030.0080.0310.0360.042
2.4LSD1蛋白在Notch靶基因Hes1基因启动子区域结合情况引物序列扩增片段在Hes1基因的位置见图5。Eca109细胞中Hes1基因不同结合区域扩增片段的富集效率见表6。从表6可以看出,与未处理细胞相比,TCP处理后细胞中LSD1蛋白在Hes1基因1和4区域扩增片段的富集效率明显降低。

TSS:转录起始位点。图5 引物序列扩增片段在Hes1基因的位置

c(TCP)/(μmol·L-1)nHes1基因结合区域1区2区3区4区030.011±0.0010.007±0.0010.006±0.0010.014±0.00110030.004±0.0010.008±0.0010.007±0.0130.011±0.001t593.2512.4191.61521.208P<0.0010.1950.2730.010
3 讨论
肿瘤的发生发展不仅取决于遗传因素,同时也受到表观遗传修饰如DNA甲基化的影响[6]。现代研究[7]认为,组蛋白的甲基化失衡和肿瘤发生存在着密切联系。LSD1是第一个被确定的组蛋白去甲基化酶,它催化H3K4me1/2和H3K9me1/2的去甲基化[8-9]。LSD1定位于细胞核内,调控基因转录的激活和抑制,被誉为细胞深处的基因“开关”,在胚胎发育和肿瘤发生的过程中起着重要的作用。Notch信号通路在细胞的分化发育、肿瘤的发生发展中有着重要作用[10-11]。
LSD1与Notch信号通路在肿瘤发生中均起到非常重要的作用,二者是否互相影响、共同调控肿瘤的发生发展,目前此方面的研究报道甚少。有报道[6]显示:在果蝇发育中SIRT1和LSD1直接相互作用,并在H4K16脱乙酰化和H3K4脱甲基化中发挥保守和协同作用,以抑制由Notch信号通路调节的基因。在T细胞急性淋巴细胞白血病中证实,组蛋白修饰即LSD1对组蛋白的去甲基化在Notch1信号通路中起重要作用[12]。并且LSD1能和其他蛋白形成复合体,调节Notch1信号通路,从而调节细胞的增殖、侵袭和转移。另外,与LSD1形成复合体的蛋白不同,LSD1发挥的功能也不同,即LSD1在Notch1信号通路中可能发挥双重作用。由于食管鳞癌是我国最常见的恶性肿瘤之一,尤其是河南省食管鳞癌的发生率和病死率均很高[13-14],因此,探讨食管鳞癌中LSD1是否对Notch通路具有调控作用具有重要意义。该研究结果显示:在食管鳞癌细胞中,用LSD1抑制剂TCP或LSD1 siRNA干预均能抑制LSD1的表达,并影响其功能;而LSD1功能受抑制后,Notch信号通路也受到抑制,表明LSD1可能调控Notch信号通路。ChIP结果显示,LSD1能与Notch通路靶基因Hes1的启动子区域结合。
综上所述,在食管鳞癌细胞中LSD1可能通过与Notch通路靶基因Hes1的启动子区域结合,从而调控Notch通路。
[1] ENZINGER PC,MAYER RJ.Esophageal cancer[J].N Engl J Med,2003,349(23):2241
[2] SHI Y,LAN F,MATSON C,et al.Histone demethylation mediated by the nuclear amine oxidase homolog LSD1[J].Cell,2004,119(7):941
[3] 马佳萍,韩剑,张金龙.组蛋白去甲基化酶的研究进展[J].中国医药指南,2017,15(12):46
[4] KESSLER M,HOFFMANN K,BRINKMANN V,et al.The Notch and Wnt pathways regulate stemness and differentiation in human fallopian tube organoids[J].Nat Commun,2015,6(4):231
[5] GARCIA A,KANDEL JJ.Notch: a key regulator of tumor angiogenesis and metastasis[J].Histol Histopathol,2012,27(2):151
[6] MULLIGAN P,YANG FJ,DI STEFANO L,et al.A SIRT1-LSD1 corepressor complex regulates Notch target gene expression and development[J].Mol Cell,2011,42(5):689
[7] MAIQUES-DIAZ A,SOMERVAILLE TC.LSD1: biologic roles and therapeutic targeting[J].Epigenomics,2016,8(8):1103
[8] HIRANO K,NAMIHIRA M.LSD1 mediates neuronal differentiation of human fetal neural stem cells by controlling the expression of a novel target gene,HEYL[J].Stem Cells,2016,34(7):1872
[9] CHEN Y,KIM J,ZHANG R,et al.Histone demethylase LSD1 promotes adipocyte differentiation through repressing Wnt signaling[J].Cell Chem Biol,2016,23(10):1228
[10]HALES EC,TAUB JW,MATHERLY LH.New insights into Notch1 regulation of the PI3K-AKT-mTOR1 signaling axis:targeted therapy of γ-secretase inhibitor resistant T-cell acute lymphoblastic leukemia[J].Cell Signal,2014,26(1):149
[11]LOPEZ CI,SAUD KE,AGUILAR R,et al.The chromatin modifying complex CoREST/LSD1 negatively regulates notch pathway during cerebral cortex development[J].Dev Neurobiol,2016,76(12):1360
[12]YATIM A,BENNE C,SOBHIAN BA,et al.NOTCH1 nuclear interactome reveals key regulators of its transcriptional activity and oncogenic function[J].Mol Cell,2012,48(3):445
[13]PARKIN DM,PISANI P,FERLAY J.Estimates of the worldwide incidence of 25 major cancers in 1990[J].Int J Cancer,1999,80(6):827
[14]LE BRAS GF,FAROOQ MH,FALK GW,et al.Esophageal cancer:the latest on chemoprevention and state of the art therapies[J].Pharmacol Res,2016,113(Pt A):236
(2016-12-08收稿 责任编辑姜春霞)
LSD1 regulates expression of related proteins of Notch pathway through combining promotor of target gene of Notch in esophageal squamous cell carcinoma cells
ZHANGXingli,HOUGuiqin,FANCheng,LIUXiangyan,GAOPan,TIANFei,BAIYiru,ZHAOQi,LUZhaoming
CollegeofPharmaceuticalSciences,ZhengzhouUniversity,Zhengzhou450001
esophageal squamous cell carcinoma;LSD1;Notch;Hes1
Aim: To explore the regulation effects and mechanism of LSD1 on Notch signaling pathway in esophageal squamous cell carcinoma(ESCC) cells.MethodsThe expressions of LSD1 and Notch signaling pathway-related proteins in five ESCC cell lines KYSE450, KYSE790, Eca109, EC9706 and TE-1 were investigated by Western blot. After ESCC cells were treated with TCP or LSD1 siRNA, respectively, the expressions of LSD1, histone H3K4me2 and the related proteins of Notch signaling pathway were detected by Western blot.ResultsThere were different expression levels of LSD1 and Notch signaling pathway-related proteins in five ESCC cell lines. After KYSE450, KYSE790 and Eca109 cells were treated with TCP, the expression of LSD1 was down-regulated while H3K4me2 was up-regulated, and those of Notch signaling pathway-related proteins were decreased(P<0.05). Similarly, LSD1 siRNA also down-regulated the expressions of LSD1 and Notch signaling pathway-related proteins(P<0.05).ConclusionLSD1 could regulate the Notch signaling pathway in ESCC by binding with the Hes1 promoter of Notch target gene.
10.13705/j.issn.1671-6825.2017.06.003
郑州大学药学院 郑州 450001
#通信作者,男,1966 年11月生,博士,副教授,研究方向:肿瘤细胞分子生物学,E-mail:lzm310@zzu.edu.cn
R735.1
*国家自然科学基金资助重点项目 81430085;河南省基础与前沿技术研究计划项目 162300410122
