短花针茅(Stipa breviflora)叶片δ13C对载畜率的响应
2017-06-15王亚婷赵天启王玺古琛陈万杰杜宇凡赵萌莉
王亚婷,赵天启,王玺,古琛,陈万杰,杜宇凡,赵萌莉
内蒙古农业大学草原与资源环境学院,内蒙古 呼和浩特 010019
短花针茅(Stipa breviflora)叶片δ13C对载畜率的响应
王亚婷,赵天启,王玺,古琛,陈万杰,杜宇凡,赵萌莉*
内蒙古农业大学草原与资源环境学院,内蒙古 呼和浩特 010019
叶片δ13C值、养分含量等特征体现了植物为获取最大碳收获所采取的生存适应策略,同时δ13C值还可以指示植物的长期水分利用效率,了解荒漠草原短花针植物的水分及资源利用策略有助于人们掌握其生长机制,对分析短花针茅应对干扰的生态对策有重要意义。采用稳定性碳同位素技术,通过测定荒漠草原短花针茅(Stipa breviflora)叶片稳定性同位素值(δ13C),结合植物叶片碳(C)、氮(N)、碳氮比(C/N)、水分含量(LWC)等生理指标,探讨不同载畜率下短花针茅叶片的适应机理。结果表明,短花针茅叶片δ13C值随放牧强度的增大而显著减小(P=0.041),其中对照处理为-26.8‰,轻度放牧处理为-27.2‰,中度和重度放牧处理分别为-27.4‰和-27.5‰,表明随着放牧强度的增大,植物的水分利用效率降低。随着放牧强度的增大,短花针茅叶片N质量分数增大(P=0.003),从对照到重度放牧处理依次为2.1%、2.3%、2.5%和2.7%;C质量分数减小,但不显著(P=0.076),从对照到重度放牧处理依次为46.3%、46.1%、46.1%和45.3%;C/N显著降低(P=0.004),对照到重度放牧处理依次为26.1%、20.7%、19.6%、18.5%,表明随着放牧干扰的增强,植物积累有机物质及资源利用能力下降。相关分析发现短花针茅叶片δ13C值与N质量分数呈显著负相关(r=-0.690,P<0.05),与C质量分数、C/N呈显著正相关(r=0.565,r=0.668;P<0.01);叶片δ13C值可指示植物有机物质的积累和资源利用能力。
稳定性碳同位素;荒漠草原;放牧强度;生理指标;水分利用效率
稳定性碳同位素技术可以揭示与植物生理生态过程相联系的一系列气候环境信息,是了解植物养分循环和生理代谢相互作用的可靠途径(Luo et al.,2006)。植物叶片的碳同位素(δ13C值)是植物组织固碳过程中内部生理特征和外部影响光合气体交换的环境因子的整合(Farquhar et al.,1984;Ponton et al.,2002),可以指示植物碳代谢的基本类型,评估植物与环境的基本关系和自身的生理机能(冯虎元等,2000;Gebrekirstos et al.,2011),是间接指示植物长期水分利用效率(WUE)的有效指标(王玉涛等,2008)。植物叶片是植物反馈环境变化较为敏感的营养器官,也是植物主要光合器官,其特征能体现植物对环境的适应或环境因子变化对植物的影响,叶片δ13C值、养分含量等特征体现了植物为获取最大碳收获所采取的生存适应策略,具有重要的生态学和生物进化意义(Cordell et al.,2001;Zhou et al.,2016)。
近年来,沿大尺度样带利用稳定性同位素技术研究植物叶片δ13C值对环境因子的响应(赵艳艳等,2016;王庆伟等,2010)逐渐成为热点,主要集中于对亚热带、内蒙古、东北等地植物及森林(郑璐嘉等,2015;熊鑫等,2016;刘艳杰等,2016)的研究,普遍认为WUE与温度、光照强度呈负相关(刘贤赵等,2011;林植芳等,1995),与大气CO2浓度、海拔呈正相关(陈拓等,2000;林光辉,2010)。植物δ13C值对环境的响应模式是生物和非生物因子共同作用的结果,单一研究植物δ13C值同环境变量的相互关系不能给予有效的解释,因此,研究植物δ13C值同其他生理指标之间的相互关系有助于我们了解植物为适应异质环境的资源节约和生长平衡等重要生态对策(Isabel et al.,2014)。
目前,对于大区域复杂自然环境内植物生理指标对碳同位素分馏作用影响机制的研究比较缺乏,结果也不尽统一。有研究者认为植物δ13C值与营养元素含量呈正相关,与叶片含水量呈负相关(马剑英等,2008);但也有研究者发现叶片δ13C值与叶片C、N、P含量呈负相关,这可能与不同植物叶片养分指标对叶片δ13C值影响机制不同有关(邱权等,2013)。
荒漠草原是亚洲中部特有的旱生性最强的草地类型,生境脆弱,对自然和人类活动干扰较为敏感,与其他草原类型相比稳定性较差(刘钟龄等,1985)。放牧作为一种典型采食作用(古琛等,2015),是该类型草地生态系统的主要利用方式,是塑造草原群落结构、功能、生物组成的关键干扰,因超越其他生态因子而成为影响荒漠草原生态系统的主导因子(李永宏,1988)。短花针茅(Stipa breviflora)是禾本科针茅属多年生旱生密丛型草本植物,是亚洲中部暖温型荒漠草原的主要建群种(内蒙古植物志编辑委员会,1994),具有生态幅广、适应性强,因其返青早、生产力高、适口性好、耐旱、耐牧性强等特点,是荒漠草原重要的牧草资源(杜利霞等,2008)。因此,加强对短花针茅的研究,从叶片生理生态特性入手,对比不同载畜率下叶片的稳定性碳同位素特征差异及其影响因素,针对水分利用效率策略掌握其响应不同载畜率的生态应对方针,对于分析不同干扰下荒漠植物的生长机制有重要的意义。
试验以荒漠草原短花针茅为研究对象,通过分析植物叶片的稳定性碳同位素组成与叶片水分含量和养分含量的关系,揭示叶片稳定性碳同位素的变化,分析短花针茅的长期水分利用效率,以期为干扰条件下植物群落的组成和演替提供理论分析基础,也为干旱条件下植物的生存策略选择提供参考。
1 材料与方法
1.1 研究区概况
试验基地位于内蒙古四子王旗短花针茅草原(41°47′7″N,111°53′46″E),海拔高度为1450 m。四子王旗位于内蒙古中部、阴山北麓和乌兰察布草原北部,属于中温带大陆性季风气候。试验基地年均气温3.4 ℃,≥10 ℃的年积温为2200~2500 ℃,无霜期为90~120 d,平均日照时数为3117.7 h。年均降水量为280 mm,年均蒸发量约为2300 mm。植被类型为短花针茅(Stipa breviflora)+无芒隐子草(Cleistogenes songorica)+冷蒿(Artemisia frigida),伴生有阿氏旋花(Convolvulus ammanni)、狭叶锦鸡儿(Caragana stenophylla)、阿尔泰狗娃花(Heteropappus altaicus)、栉叶蒿(Neopallasia pectinata)、木地肤(Kochia prostrata)、羊草(Leymus chinesis)等。土壤类型为淡栗钙土。
1.2 试验设计
1.2.1 样地分布
四子王旗短花针茅放牧试验始于2003年6月,采用完全随机区组设计,将试验小区分为3个区组,每个区组设置4种不同处理,分别为对照区(CK)、轻度放牧区(LG)、中度放牧区(MG)、重度放牧区(HG),每个处理各设置3次重复,分别编号为1、2、3(见图1)。依据王明玖等(1994)、卫智军等(2000)对短花针茅草原载畜率的研究,再结合草地实际调查的结果,试验区载畜率值依次为0、0.91、1.82和2.71羊单位/(公顷·半年),放牧绵羊只数分别为0、4、8和12只。各小区的管理措施一致(排除人工误差),试验饲喂羊为当地成年蒙古羯绵羊,每3年更换一批试验饲喂羊,以保证其正常的采食和消化能力,减少因客观误差对试验造成的影响。2003—2016年每年放牧期为半年,从6月1日开始,到11月30日结束。每日早晨6点将绵羊赶入各小区让其自由采食,下午6点赶回圈内饮水、休息。野外取样时间一般选择在该地区生物量的高峰期。

图1 试验样地各放牧小区和区组位置示意图Fig. 1 The experimental paddocks and blocks location
1.2.2 取样方法
2016年8月5—6日在小区内随机采集植物样品。在每个样地沿着南北方向间隔50 m选取短花针茅群落5处,每个群落中分单株随机采集5株中等大小、株丛径基本一致的健康无病虫害的植株,在每个植株上选择5个完全伸展、发育成熟的健康叶片(尽量在每株植物的同一部位采集),用剪刀剪下,混合;采样时,叶片表面的尘土等杂质用毛刷刷净,装入塑料袋后立即带回实验室。
将所取的短花针茅植物样品分为3份,一份用于测定植物的稳定性碳同位素,一份用于测定植物的叶片含水量,剩余的一份用于测定植物叶片的营养元素。
1.2.3 实验测定
(1)植物叶片稳定性碳同位素的测定
将用于测定植物稳定性同位素的短花针茅样品用去离子水洗净并晾干,置于65 ℃恒温条件下烘干48 h,然后用球磨仪粉碎,密封保存。将所有处理好的样品用十万分之一的电子天平分别称取0.7~0.8 mg于小的锡纸帽中包好,利用碳同位素分析仪测定其碳同位素值,每种植物样品重复测定3次。样品的氧化条件为1020 ℃,δ13C值采用PDB标准进行校准,测量误差小于0.05‰。植物叶片δ13C值利用以下公式计算:

式中,Rp是样品中碳元素的重、轻同位素丰度之比(13Cp/12Cp);Rs是国际通用标准物的重、轻同位素丰度之比(13Cs/12Cs),13C标准物为PDB(一种出自美国南卡罗来那州的碳酸盐陨石)。
(2)植物叶片含水量及营养元素的测定
植物叶片含水量测定采用烘干称重法:取事先准备用于测定植物叶片含水量的样品,称取鲜重,然后于65 ℃下烘干后称取干重。
LWC=[(鲜重-干重)/鲜重]×100%
植物全氮、全碳的测定利用元素分析仪。
1.3 数据处理
所测定的数据在Excel中处理后,采用R.3.3.2统计软件进行方差分析、相关分析,利用SigmaPlot 12.5绘制图表。
2 结果与分析
2.1 不同载畜率稳定性碳同位素分布
由图2可知,随着放牧强度的变化,短花针茅叶片δ13C值在-26.5‰~-27.5‰之间变化,平均值为-27.2‰。一般认为全球范围内C3植物的δ13C值为-23‰~-32‰,而任书杰等(2011)研究表明中国478种C3物种叶片δ13C的变化范围为-22‰~-33.5‰,平均值为-27.1‰。本研究中短花针茅叶片δ13C值均落在两者的范围内,平均值与之相比差别较小,表明短花针茅属于C3植物。

图2 不同放牧强度下短花针茅叶片稳定性碳同位素值Fig. 2 The value of foliar δ13C under different grazing rates
随着放牧强度的增加,短花针茅叶片δ13C值偏向更负的值,其中,对照处理为-26.8‰,轻度放牧处理为-27.2‰,中度和重度放牧处理分别为-27.4‰、-27.5‰,对照显著高于中度和重度(P=0.041),放牧显著降低了叶片δ13C值。
2.2 不同载畜率下短花针茅叶片生理指标的变化
对不同载畜率下短花针茅叶片生理指标进行分析发现(图3),随着放牧强度增加,植物叶片各生理指标都发生了不同程度的变化。其中,叶片N质量分数在2.0%~3.0%之间变化,轻度放牧处理为2.3%,中度为2.5%,重度为2.7%,对照为2.1%,且随着放牧强度的增大,叶片N质量分数显著升高(P=0.003);叶片C质量分数在45%~46.5%之间变化,其中,对照处理为46.3%,轻度放牧处理为46.1%,中度为46.1%,重度为45.3%,呈逐渐降低的趋势,但并不显著(P=0.076);叶片C/N比值的变化范围是18%~22%,对照处理为21.6%,轻度放牧处理为20.7%,中度为19.6%,重度为18.5%,C/N比值随着放牧强度的增大而显著降低(P=0.004);叶片LWC的含量为轻度放牧处理53%,中度62%,重度61%,对照48%,总体随放牧强度的增大而显著升高(P=0.094)。
2.3 短花针茅叶片稳定性碳同位素、载畜率及生理指标的关系
对4种不同载畜率下所测得的短花针茅叶片δ13C、载畜率及生理指标进行相关分析和回归分析,结果表明(表1,图4),短花针茅叶片δ13C值与其叶片养分间均存在显著线性相关关系,其中,叶片δ13C值与N质量分数呈显著负相关(r=0.690,P<0.05),植物可通过降低N素的积累来提高自身的δ13C值,即提高水分利用效率;叶片δ13C值与C质量分数呈极显著正相关(r=0.565,P<0.01),与C/N呈极显著正相关(r=0.668,P<0.01),植物通过有机物质的积累提高自身的资源利用能力和水分利用效率;叶片δ13C值与LWC和载畜率间没有显著相关性性(P=-0.241,-0.281),植植物的水分利利用效率与叶片片水分的有效效性及放牧强强度间不存在在直接线性关系系。

表1 短花针茅植物碳同位素、载畜率及生理指标间的相关关系Table 1 Correlation between carbon isotope、stocking rates and physiological index of plant foliar
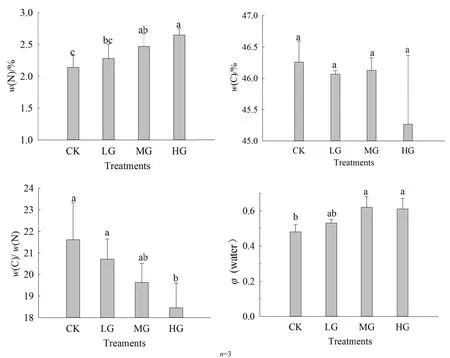
图3 不同放放牧强度下短花针针茅生理指标值Fig. 3 Thee value of foliar phhysiological indicees under different ggrazing rates
叶片的的营养元素除除与叶片的δ13C呈显著相相关性外,与载载畜率也呈显著相关。其中,N质量分分数与载畜率呈呈极显著正相相关(r=0.756,P<0.01)),C质量分数与与载畜率呈极极显著负相关(r=-0.7999,P<0.01),CC/N与载畜率率呈显著负相相关(r=-0.6448,P<0.05),叶叶片含水量与与载畜率间相相关性不显著著。
3 讨论
3.1 不同载载畜率下短花花针茅叶片δδ13C值的变化化
δ13C是是植物干物质质中稳定性性碳同位素比比率(13C/12C)相对于大气气中用于植物光合作用用的13C/12C的度度量,它反映映出植物在光光合作用期间间对13C的主动分分辨能力(王王庆伟等,20010)。不同载载畜率下叶片稳稳定性碳同位位素变化表现为为:随着放牧牧强度的增大,叶片δ13C偏偏向更负的值,其水分利用用效率降低;这这与李昕宇(2013)的研究究结果相似。。植物叶片δ13CC值受植物自身遗传因素素和环境因素素的共同影响,由于植物-环环境连续体的相互作用,使使得这两两方面又形成成不可忽略的的联系,特别别是植物在不不断适适应环境变化化的同时,环环境也在不断断影响着植物物的生生长(马晔等等,2013),所以植物在在面对不同的的干扰扰时,对COO2的分辨能力力不同,导致致叶片δ13CC值发发生变化;放放牧家畜通过过对植被的践践踏、采食,破坏坏了植物叶片片的外部形态态及生长性能能,从而使其其光合合能力降低,进一步引起起胞间CO2浓浓度改变,植植物对对CO2的识别别能力提高,所以叶片δδ13C值降低。
3.22 不同载畜率率下短花针茅茅叶片生理指指标的变化
植物体内的的物质由结构构性物质、功功能性物质和和贮藏藏性物质3类类组成,一般般情况下,C是结构性物物质,N是功能性性物质。对于于同一物种来来说,往往结结构性性物质受环境境的影响较小小,含量相对对较稳定,而而功能能性和贮藏性性物质的含量量受环境影响变化较大大(Roobert et al.,22002)。本研研究中,C作作为结构性物物质在在短花针茅中中对载畜率的的响应不敏感感,虽有降低低的趋趋势但并没有有显著差异,这与上述规规律较一致。这是是因为C是组组成生物量的的基本元素,与植物体的的其他他养分含量相相比,其所占占比例很大(平均为46%%左右右),而放牧牧对植物叶片片C的影响较较小,在统计计上并没有表表现出显著的差异(丁小慧慧等,2012)。

图4 植物叶片片δ13C值与生理指指标的相关关系Fig. 4 The coorrelation betweenδ13C value and phhysiological indices of plant leaves
本研究究表明放牧会会显著改变短短花针茅叶片片N质量分数,随着放牧强度度的增大,叶叶片N质量分分数提高。这与与牛得草等(2011)对黄土土高原地区关关于长芒草(Stiippa bungeanaa)的研究结果果趋同。放牧牧过程中家畜通通过采食、践踏作用消除植植物地上部分分衰老的组织,刺激了植株的再生生长,植物新生的的幼嫩组织通常常具有较高的养分吸收效率率,以迅速提提高叶片中的叶叶绿素含量,促进光合作用用,补偿放牧牧减少的生物量量。N是叶绿素的主要组成成成分,放牧牧干扰下植物叶叶片N质量分分数高,这也是是草原植物补补偿生长的一种种表现(贾婷婷婷等,2013)。
本研究究中,C/N比值值随着载畜率率的增加而降降低,主要是因为为放牧使得叶片N质量分分数升高,C质质量分数降低(但但并不显著),所以叶片片C/N比值降降低。C/N比值大大小表征植物吸收单位养分分元素含量所所同化C的能力力,对调节植物生长有着极极其重要的作作用(薛利红等,,2006),在在一定程度上上可以反映植植物体养分元素素的利用率。本研究结果表表明,随着载载畜率的的增加,植物物的资源利用用能力整体下下降,且载畜畜率对对叶片生理指指标的影响并并不一致,植植物通过不同同的方方式对各项生生理指标进行行调节,以适适应不同的放放牧压压力。
3.33 短花针茅叶片δ13C与与氮素利用效效率
一些学者认认为,植物的的δ13C与氮氮素利用效率率(NUUE)之间存存在权衡,在在自然条件下下,拥有较高高δ13CC的植物,是是以低NUE为代价的(李机密等,20008)。植物叶片片水平的NUUE可以用单单位质量叶NN的倒倒数来表示((Aerts et all.,2000),长长期的NUEE可由由叶片的C/NN比值反映(Livingston eet al.,1999)。本研研究中,短花花针茅叶片的的δ13C与叶片片N质量分分数呈呈显著负相关关,与叶片的的C/N呈正相相关,此结果果与李李善家等(22011)、王英英姿(2013)研究结果一一致,其原因可能能是植物用于于碳同化过程的N占叶片片总N含量的比例较小,而大大部分N存存在于其他蛋蛋白质质和结构组分分之中,即这这部分植物的的光合氮利用用效率率较低,导致致植物的δ13CC值与叶片N质量分数数之间间存在负相关关(史作民等等,2011),出于对干旱旱和干扰环境的一种适应策略,短花针茅可能同时具有较低的水分利用效率和资源利用效率,不存在相互制约的补偿机制。从另外的角度来讲,可以推断本研究所选短花针茅叶片δ13C值变化的主要影响因素可能是气孔限制,并不是光合效率,故叶片营养元素N含量变化并不直接对叶δ13C值产生积极的影响(邱权等,2013)。可见,必须同时考虑叶片氮及其光合作用效率才能准确地反映植物叶片氮含量与光合能力、δ13C值之间的关系,从而更好地理解植物的资源利用策略。
4 结论
(1)随着载畜率的增加,短花针茅叶片的δ13C值显著降低(P=0.041)。
(2)短花针茅叶片N质量分数随着载畜率的增加而显著增加(P=0.003),而C质量分数逐渐降低但不显著(P=0.076),C/N和LWC显著减小(P=0.004,0.094)。
(3)短花针茅叶片δ13C值与N质量分数呈显著负相关(r=0.69,P<0.05),与C质量分数、C/N呈显著正相关(r=0.565,r=0.668;P<0.01),与LWC没有显著相关性(r=0.647,P=0.241)。
AERTS R, CHAPIN F S. 2000. The mineral nutrition of wild plants revisited: A revaluation of processes and patterns [J]. Advances in Eco1ogical Research, 30: 61-67.
CORDELL S, GOLDSTEIN G, MEINZER F C, et al. 2001. Regulation of leaf life-span and nutrient use efficiency of Metrosidero polymorpha trees at two extremes of a long chrono sequence in Hawaii [J]. Oecologia, 127(2): 198-206.
FARQUHAR G, RICHARDS R. 1984. Isotopic composition of plant carbon correlates with water-use efficiency of wheat genotypes [J]. Functional Plant Biology, 11(6): 539-552.
GEBREKIRSTOS A, NOORDWIJK M V, NEUFELDT H, et al. 2011. Relationships of stable carbon isotopes, plant water potential and growth: an approach to asses water use efficiency and growth strategies of dry land agro forestry species [J]. Trees, 25(1): 95-102.
ISABEL P A, QUEREJETA J I, MARIA NAZARET G A, et al. 2014. Elemental and stable isotope composition of Pinus halepensis foliage along a metal (loid) polluted gradient: implications for phytomanagement of mine tailings in semiarid areas [J]. Plant and Soil, 379(1-2): 93-107.
LIVINGSTON N J, GUY R D, SUN Z J, et al. 1999. The effects of nitrogen stress on the stable carbon isotope composition, productivity and water use efficiency of white spruce (Picea glauca ( Moench) Voss) seedlings [J]. Plant, Cell & Environment, 22(3): 281-289.
LUO J X, ZANG R G, LI C Y. 2006. Physiological and morphological variations of Picea asperata populations originating from different altitudes in the mountains of southwestern China [J]. Forest Ecology and Management, 221(1-3): 285-290.
PONTON S, DUPOUEY J L, BREDA N, DREYER E. 2002. Comparison of water-use efficiency of seedlings from two sympatric oak species: genotype environment interactions [J]. Tree Physiology, 22(6): 413-422.
ROBERT W S, JAMES J E. 2002. Ecological Stoichiometry: The Biology of Elements from Molecules to the Biosphere [M]. Princeton: Princeton University Press.
ZHOU Y C, CHENG X L, FAN J W, et al. 2016. Relationships between foliar carbon isotope composition and elements of C3species in grasslands of Inner Mongolia, China [J]. Plant Ecology, 217(7): 883-897.
陈拓, 秦大河, 任贾文, 等. 2000. 甘肃马衔山平车前叶片δ13C的海拔和时间差异[J]. 西北植物学报, 20(4): 672-675.
丁小慧, 宫立, 王东波, 等. 2012. 放牧对呼伦贝尔草地植物和土壤生态化学计量学特征的影响[J]. 生态学报, 32(15): 4722-4730.
杜利霞, 李青丰. 2008. 放牧对荒漠草原短花针茅繁殖特性的影响[J].山西农业大学学报: 自然科学版, 28(1): 4-6.
冯虎元, 安黎哲, 王勋陵. 2000. 环境条件对植物稳定碳同位素组成的影响[J]. 植物学通报, 17(4): 312-318.
古琛, 杜宇凡, 乌力吉, 等. 2015. 载畜率对荒漠草原群落及植物功能群生物量的影响[J]. 生态环境学报, 24(12): 1962-1968.
贾婷婷, 袁晓霞, 赵洪, 等. 2013. 放牧对高寒草甸优势植物和土壤氮磷含量的影响[J]. 中国草地学报, 35(6): 80-85.
李机密, 黄儒珠, 郑怀舟, 等. 2008. 长汀水土流失区重建植被WUE及相关叶性状研究[J]. 亚热带资源与环境学报, 3(4): 23-30.
李善家, 张有福, 陈拓. 2011. 西北油松叶片δ13C特征与环境因子和叶片矿质元素的关系[J]. 植物生态学报, 35(6): 596-604.
李昕宇. 2013. 放牧、增雨对典型草原植物稳定性同位素效应的影响[D].呼和浩特: 内蒙古大学.
李永宏. 1988. 内蒙古锡林河流域羊草草原和大针茅草原在放牧影响下的分异和趋同[J]. 植物生态学与地植物学学报, 12(3): 27-34.
林光辉. 2010. 稳定同位素生态学: 先进技术推动的生态学新分支[J].植物生态学报, 34(2): 119-122.
林植芳, 林桂珠, 孔国辉, 等. 1995. 生长光强对亚热带自然林两种木本植物稳定碳同位素比、细胞间CO2浓度和水分利用效率的影响[J].热带亚热带植物学报, 3(2): 77-82.
刘贤赵, 王国安, 李嘉竹, 等. 2011. 中国北方农牧交错带C3草本植物δ13C与温度的关系及其对水分利用效率的指示[J]. 生态学报, 31(1): 123-136.
刘艳杰, 许宁, 牛海山. 2016. 内蒙古草原常见植物叶片δ13C和δ15N对环境因子的响应[J]. 生态学报, 36(1): 235-243.
刘钟龄, 雍世鹏, 王义风. 1985. 内蒙古植被[M]. 北京: 科学出版社.
马剑英, 陈发虎, 夏敦胜, 等. 2008. 荒漠植物红砂叶片δ13C值与生理指标的关系[J]. 应用生态学报, 19(5): 1166-1171.
马晔, 刘锦春. 2013. δ13C在植物生态学研究中的应用[J]. 西北植物学报, 33(7): 1492-1500
内蒙古植物志编辑委员会. 1994. 内蒙古植物志[M]. 第二版. 第五卷.呼和浩特: 内蒙古人民出版社: 200-202.
牛得草, 董晓玉, 傅华. 2011. 长芒草不同季节碳氮磷生态化学计量特征[J]. 草业科学, 28(6): 915-920.
邱权, 李吉跃, 王军辉, 等. 2013. 柴达木盆地白刺叶片δ13C与叶片和土壤养分指标的关系[J]. 西北植物学报, 33(11): 2301-2308.
任书杰, 于贵瑞. 2011. 中国区域478种C3植物叶片碳稳定性同位素组成与水分利用效率[J]. 植物生态学报, 35(2): 119-124.
史作民, 程瑞梅, 刘世荣. 2004. 高山植物叶片δ13C的海拔响应及其机理[J]. 生态学报, 24(12): 2901-2906.
王明玖, 马长升. 1994. 两种方法估算草地载畜量的研究[J]. 中国草地, (5): 19-22.
王庆伟, 于大炮, 代力民, 等. 2010. 全球气候变化下植物水分利用效率研究进展[J]. 应用生态学报, 21(12): 3255-3265.
王英姿. 2013. 灵石山不同海拔米槠林优势种叶片δ13C值与叶属性因子的相关性[J]. 生态学报, 33(10): 3129-3137.
王玉涛, 李吉跃, 程炜, 等. 2008. 北京城市绿化树种叶片碳同位素组成的季节变化及与土壤温湿度和气象因子的关系[J]. 生态学报, 28(7): 3143-3151.
卫智军, 韩国栋, 杨静, 等. 2000. 短花针茅荒漠草原植物群落特征对不同载畜率水平的响应[J]. 中国草地, (6): 1-5.
熊鑫, 张慧玲, 吴建平, 等. 2016. 鼎湖山森林演替序列植物-土壤碳氮同位素特征[J]. 植物生态学报, 40(6): 533-542.
薛利红, 杨林章, 范小晖. 2006. 基于碳氮代谢的水稻氮含量及碳氮比光谱估测[J]. 作物学报, 32(3): 430-435.
赵艳艳, 徐隆华, 姚步青, 等. 2016. 模拟增温对高寒草甸植物叶片碳氮及其同位素δ13C和δ15N含量的影响[J]. 西北植物学报, 36(4): 777-783.
郑璐嘉, 黄志群, 何宗明, 等. 2015. 林龄、叶龄对亚热带杉木人工林碳氮稳定同位素组成的影响[J]. 林业科学, 51(1): 22-28.
2017. Respons of Stipa breviflora leaf δ13C to stocking rates [J]. Ecology and Environmental Sciences, 26(4): 613-619.
WANG Yating, ZHAO Tianqi, WANG Xi, GU Chen, CHEN Wanjie, DU Yufan, ZHAO Mengli.
Respons of Stipa breviflora Leaf δ13C to Stocking Rates
WANG Yating, ZHAO Tianqi, WANG Xi, GU Chen, CHEN Wanjie, DU Yufan, ZHAO Mengli
College of Grassland, Resources and Environment, Inner Mongolia Agricultural University, Hohhot 010019, China
The carbon stable isotope (δ13C) value and nutrient content in plant leaves reflect the survival and adaption strategy of plants to absorb carbon maximum optimally. The long-term water use efficiency (WUE) of plant was indicated by the δ13C value as well. The objective of this study was to assess the effect of different stocking rates on resource utilization of Stipa breviflora. The δ13C, carbon (C), nitrogen (N), C/N ratio, water content (LWC) in leaves were determined to analyze the adaption mechanism. Results showed that: the δ13C value in S. breviflora leaves reduced (P=0.041) as stocking rates increased, and they were -26.8‰, -27.2‰, -27.4‰, and -27.5‰ in control check (CK), light grazed (LG), moderate grazed (MG) and heavy grazed (HG), respectively. That also meant the WUE decreased with increasing stocking rates. The N percent in S. breviflora leaves elevated as stocking rates increased, and they were 2.1%, 2.3%, 2.5% and 2.7%, respectively. While the C percent in S. breviflora leaves were not observed differences (P=0.076) and they were 46.3%, 46.1%, 46.1% and 45.3% with increasing stocking rates. The C/N ratio in S. breviflora leaves were lowered (P=0.004) as stocking rates increased, and they were 26.1%, 20.7%, 19.6% and 18.5%, respectively. That meant organic matter accumulation and resource absorption of S. breviflora declined with increasing stocking rates. The δ13C were negatively associated with N percent (r=-0.690, P<0.05) while positively related to C percent and C/N ratio (r=0.565, r=0.668; P<0.01). Leaf δ13C values could indicate the accumulation of plant organic matter and resource utilization.
stable carbon isotope; desert steppe; stocking rates; physiological indices; water use efficiency
10.16258/j.cnki.1674-5906.2017.04.010
Q945.79; X173; S812.29
A
1674-5906(2017)04-0613-07
王亚婷, 赵天启, 王玺, 古琛, 陈万杰, 杜宇凡, 赵萌莉. 2017. 短花针茅(Stipa breviflora)叶片δ13C对载畜率的响应[J]. 生态环境学报, 26(4): 613-619.
国家自然科学基金项目(31460110);内蒙古农业大学草业与草地资源重点实验室
王亚婷(1990年生),女,硕士研究生,主要从事草地生态学方向研究。E-mail: 791831520@qq.com
*通信作者。赵萌莉,E-mail: nmgmlzh@126.com
2017-03-06
