稻谷贮藏蛋白与米饭质地研究
2017-06-05李兴军
◎李兴军 韩 旭 王 昕
稻谷贮藏蛋白与米饭质地研究
◎李兴军 韩 旭 王 昕
稻谷主要贮藏蛋白是米谷蛋白,占总蛋白含量的80%,醇溶蛋白占百分之几。本文就稻谷贮藏蛋白的类型、颖果内沉积、提取方法及理化定性进行回顾与综述,同时重点讨论了贮藏蛋白影响大米糊化和米饭质地的机理,提出稻谷贮藏蛋白研究的热点方向。
稻谷 大米 谷蛋白 醇溶蛋白 蛋白质组学
稻米是世界半数人口的大宗食物,主要提供能量和蛋白质。与小麦比较,稻谷贮藏蛋白的功能特性近年研究有进展。稻谷主要贮藏蛋白类似豆类和其它双子叶植物的11S球蛋白 (legumins),醇溶性蛋白只是其微少的成分。稻谷贮藏蛋白基本是谷蛋白 (glutelin),属于球蛋白家族,而小麦、大麦和黑麦是醇溶蛋白,属于麦醇溶蛋白家族[1]。贮藏蛋白质之间共价和非共价相互作用引起结构差异,导致大米水合的蛋白复合物的大小分布不同于小麦,稻谷中聚合蛋白成分与单聚物强烈相互作用,聚合物的链长小于小麦,米粉团粘弹性质量低于小麦,但是小麦质量评价的面团试验方法适合定性米粉团 (Gujral and Rosell 2004)。本文对稻谷贮藏蛋白的提取定性及功能研究进展,以期对我国稻谷储藏及加工有指导意义。
一、稻谷蛋白含量与贮藏蛋白类型
稻谷蛋白含量范围在6%-9%,品种之间有差别。蛋白含量受气候、农艺条件所影响[2]。施用氮肥增加稻谷蛋白含量,在抽穗期喷洒硫酸铵溶液,蛋白含量从6%增加到10%;施用尿素,蛋白含量仅略增加[3]。稻谷蛋白质在籽粒中分布不均匀,在麸皮和胚乳外周浓度较高,接近籽粒中心则含量减小。抛光的大米比糙米蛋白含量低,加工的大米还大约保持了原有蛋白的82%。
按照 Osborne对谷物贮藏蛋白分类方法,Cagampang等 (1966)对脱壳和抛光的3个稻谷品种蛋白 (含量6.8-8.5%)分析表明,水溶性蛋白(清蛋白)占总蛋白的3.8-8.8%,盐溶性蛋白(球蛋白)占 9.6-10.8%,醇溶性蛋白占 2.6-3.3%,碱溶性蛋白 (谷蛋白)占66.1-78.0%。也常用变性剂SDS和还原剂 (DTT、2-ME)提取稻谷谷蛋白。稻谷谷蛋白氨基酸组成较醇溶蛋白平衡,醇溶蛋白缺乏赖氨酸和色氨酸。与其它谷物比较,稻谷谷蛋白含量虽低,却是其品质好于其它谷物。
有生物活性的稻谷清蛋白分子量范围较宽,10-20 kDa(Juliano 1985)。球蛋白组由四类球蛋白α、β、γ、δ组成,表观分子量范围16-200 kDa[4]。醇溶蛋白在稻谷胚乳中的含量是 18-20%[5]。根据电泳迁移率,区分的稻谷醇溶蛋白的3种亚基表观分子量分别是10、13和16 kDa(Ogawa等1987),只有亚基13 kDa在没有还原剂的情况下溶解于乙醇,而含半胱氨酸多的其它两种亚基能够形成长的聚合物,溶解时需要还原(Hibino等1989)。
稻谷谷蛋白亚基能够形成巨型的高分子复合物,被二硫键和疏水相互作用所稳定 (Utsumi 1992)。除了氢键和二硫键外,大量的聚合作用(Sugimoto等1986)和糖基化作用 (Wen和Luthe 1985),导致稻谷聚合的谷蛋白层分不溶于水限[6]。稻谷谷蛋白以前体57 kDa多肽合成后,转录时切成两种亚基α(酸性)和β(碱性)亚基,表观分子量各是30-39和19-25 kDa(Juliano 1985)。这两个亚基通过分子间二硫键共价连接在一起,导致谷蛋白分子量变化范围64~500 kDa。这种高度有序的结构解释了稻谷谷蛋白缺乏功能特性。
至今鉴定了6个稻谷谷蛋白基因,按照核苷酸序列相似性将它们分为A-和B-型谷蛋白亚家族[7]。A-和B-型谷蛋白分别有8个和5个半胱氨酸残基,其中4对半胱氨酸形成两类二硫键,一类是α和β多肽之间形成分子内二硫键,另一类是α多肽内形成二硫键。
二、颖果内贮藏蛋白定位
关于水稻贮藏蛋白在颖果内积累,Kim等(1986)报道,水稻花后3-8天蛋白显著积累,增加32、21、15 kDa条带。Udaka等[8]报道,稻谷胚乳在花后5-7天积累谷蛋白,花后7-9天积累醇溶蛋白。SDS-PAGE分析表明,醇溶蛋白是15 kDa的单肽,谷蛋白两个亚基α和β各是32和21kDa(Zhao等1983)。醇溶蛋白和谷蛋白的物理化学特性在古代水稻与现代水稻品种之间类似。不过,Udaka等 [8]观察到醇溶蛋白类型对古水稻Kuro-mochi品种是15 kDa多肽,对古水稻Tsushimazairai品种包括15和13 kDa。Ogawa等[9]对红粒稻米Nihonsakari品种观察到醇溶蛋白三个亚基各是10、13、16 kDa。还需要分析醇溶蛋白是单聚物还是多聚物?
现有的稻谷醇溶蛋白的报道表明,这类蛋白许多特性如分子量、氨基酸序列等,不同于大多数其它谷物的醇溶蛋白。稻谷醇溶蛋白和谷蛋白在籽粒的不同蛋白体中沉积[10]。按照蛋白体来源,水稻有两类蛋白体 (PB),内质网来源的PB-I含有醇溶蛋白,占大米总蛋白的20%;液胞PB-II含有谷蛋白和球蛋白,占总蛋白的60%。按照形状,Bechtel和 Pomeranz (1978)鉴定了晶体型、小球形和大球形3种蛋白体。稻谷胚乳中心区域仅含有大球形蛋白体,整个胚乳组织无数小球形蛋白体,而大球形蛋白体最少。Tanaka等(1980)观察到大球形蛋白体在透射电子显微镜下显示离心环,它是醇溶蛋白和谷蛋白的沉积位点;小球形蛋白体内基本上是谷蛋白,分布遍及淀粉胚乳。在蒸煮过程中蛋白体能够保持原样,大球形蛋白体对消化酶有抗性,降低了蛋白质的消化率。这没有成为一个营养问题,由于稻谷蛋白的生物价随着蛋白消化率降低而增加,净蛋白利用率与其它谷物蛋白接近,不消化的蛋白是一部分醇溶蛋白 (Tanaka等1978)。
稻谷胚乳中没有发现基质蛋白 (Harris and Juliano 1977),这是水稻区分于其它谷物种类。其它谷物中,大量的蛋白以颗粒之间的基质形式存在。可解释为,稻谷胚乳中蛋白体之间连接的蛋白微纤丝,形成一个网络,在电子显微镜下难以观察到。已经鉴定了一个淀粉粒联合的60 kDa蛋白,也叫蜡质基因产物,包埋在淀粉颗粒中[11-12]。这个蛋白是淀粉合成酶 (GBSS),与直链淀粉含量相关。
三、稻谷贮藏蛋白提取方法、理化特性及氨基酸组成
(一)分离方法
广泛研究了稻谷蛋白质的提取。Shewry和Miflin (1985)认为稻谷醇溶蛋白提取率,50%的丙醇>55%异丙醇>60-70%乙醇。Sugimoto等(1986)表明,5种醇水溶液中,55%丙醇提取水稻醇溶蛋白最有效。15 kDa亚基对应醇水溶液提取的醇溶蛋白。Udaka等[8]采用4种醇的水溶液对5个古水稻品种和5个现代品种提取醇溶蛋白。55%丙醇提取最有效,提取3.3-7.8%的总蛋白。55%的叔丁基-丁醇提取2.6-4.9%的总蛋白。70%乙醇和60%的异丙醇提取稻谷醇溶蛋白最不高效。他们采用不同试剂提取稻谷谷蛋白,发现0.1%NaOH、2%SDS、2%SDS+0.5%2-ME、2% SDS+5M尿素+0.5%2-ME均能够提取87%以上的谷蛋白,其中2%SDS+5 M尿素+0.5%2-ME对提取谷蛋白最有效,加入2-ME稍微提高。谷蛋白层分通常包含淀粉,在尿素存在下淀粉与谷物样品发生凝胶化,导致谷蛋白难以提取(Juliano和Boulter 1976)。
采用碱液溶解稻谷谷蛋白并加入巯基乙醇打破二硫键,或碱性提取缓冲液中加入6-8M尿素或0.5%SDS (Snow and Brooks 1989)。0.05%乙酸和5M尿素不能够提取谷蛋白。8M尿素解离谷蛋白多肽不是特别有效。2%SDS加到10 mM Tris-HCl溶液 (pH 8.5)可以提取谷蛋白。
Van Der Borght等[14]对一个粳稻品种提取胚乳蛋白,2%SDS提取64%的蛋白,加入6 M尿素提取率达到79%。采用20-100 mM NaOH溶液,提取70-81%的蛋白。最佳的蛋白提取率2%SDS +6.M尿素+0.5-1.5%DTT。0.1%NaOH比2%SDS提取谷蛋白高效,但是引起蛋白降解。碱液提取期间谷蛋白构象发生改变。在高浓度碱液,由于电荷排斥谷蛋白解离为亚基;蛋白质被修饰导致解聚集、分子量减少而变得可溶 (Shewry and Miflin 1985,Hamada 1997)。可见,分离和溶解大米谷蛋白的高效方法还没有建立。定性大米蛋白需要了解其溶解的分子基础。
对米糠蛋白,强烈的解离溶液是0.1M乙酸+ 3.0 M 尿素+0.01 M 溴化十六烷基三甲基铵(CTAB),可提取蛋白67-94%[13]。
(二)理化特性
Li和 Okita (1988)从水稻品系M201分离的基因编码15 kDa的醇溶蛋白。古水稻和现代水稻均具有 15 kDa的醇溶蛋白。 Horikoshi等(1991)电泳时表明,粳稻日本晴 15 kDa的醇溶蛋白 5个条带的p I各是5.6、7.1、7.3、7.6和7.8。Ukada等 [8]对Akita-kura稻谷品种醇溶蛋白SDS-PAGE条带纯化,随后二维电泳发现几个条带p I在5.5-6.5。古水稻和现代水稻的谷蛋白由两个亚基α和β组成。谷蛋白的二维电泳表明,两个21和32 kDa条带分别是酸性和碱性多肽。古水稻黑粒和红粒大米的总蛋白与两种贮藏蛋白含量之间有相关性,醇溶蛋白回收率3.0%-7.8%,总蛋白则6.9%-11.4%;12个古水稻品种的醇/谷蛋白比率是4.9:78.1。现代水稻与古水稻蛋白含量几乎一样。
采用凝胶过滤层析分析稻谷谷蛋白分子量分布,Tecson等 (1971)采用Sephadex G-200层析,还原和烷基化作用导致谷蛋白部分解离为亚基,获得了完整谷蛋白1/10分子量的层分 (MW 65 kDa)。Villareal和Juliano (1978)在Ultrogel 44柱分离稻谷谷蛋白,效果不佳,在Ultrogel 34再次层析获得均一的亚基。Snow和 Brooks (1989)采用Sepharose CL-6B凝胶过滤层析分离稻谷粗蛋白提取物,洗脱的4个层分没有完全分辨,MW范围较宽。将阴离子交换和凝胶过滤结合,Zarins和Chrastil(1992)分离了谷蛋白33、22、14 kDa三种亚基。Hamada等[13]在ShodexProtein WS-2003层析分离米糠蛋白,重复性好,测定了表观分子量分布,深入层析和定性了单一多肽,但是没有获得分辨很好的峰。Katsube-Tanaka等[7]采用Sephacryl S-300HR分离低分子量聚合的谷蛋白。Van Der Borght等[14]采用Shodex Protein KW-80凝胶柱层析获得6个峰。峰I、II是谷蛋白α和β亚基形成的二聚物、三聚物或高级聚合物;峰III是谷蛋白α和β亚基通过分子间二硫键形成的二聚物;峰IV和V分布是谷蛋白α和β亚基;峰VI含有低分子量的清蛋白、球蛋白及醇溶蛋白。高分子量谷蛋白聚合物在还原条件时发生部分解离。总之通过凝胶过滤分离稻谷谷蛋白多肽,分离α和β亚基不高效。
综上所述,采用不同的方法分析稻谷蛋白,分离的层分含有重叠的数量不同成分,而不是分离成为单一成分。采用还原剂如DTT、2-ME或化学试剂如SDS、尿素处理,改变谷蛋白的聚合行为,它们被分解成亚基,这些亚基通过SDSPAGE或者反向高效液相色谱进行分离。完整的和还原稻谷蛋白分子量的分布采用粒度排阻色谱(SE-HPLC)定性。
(三)氨基酸组成
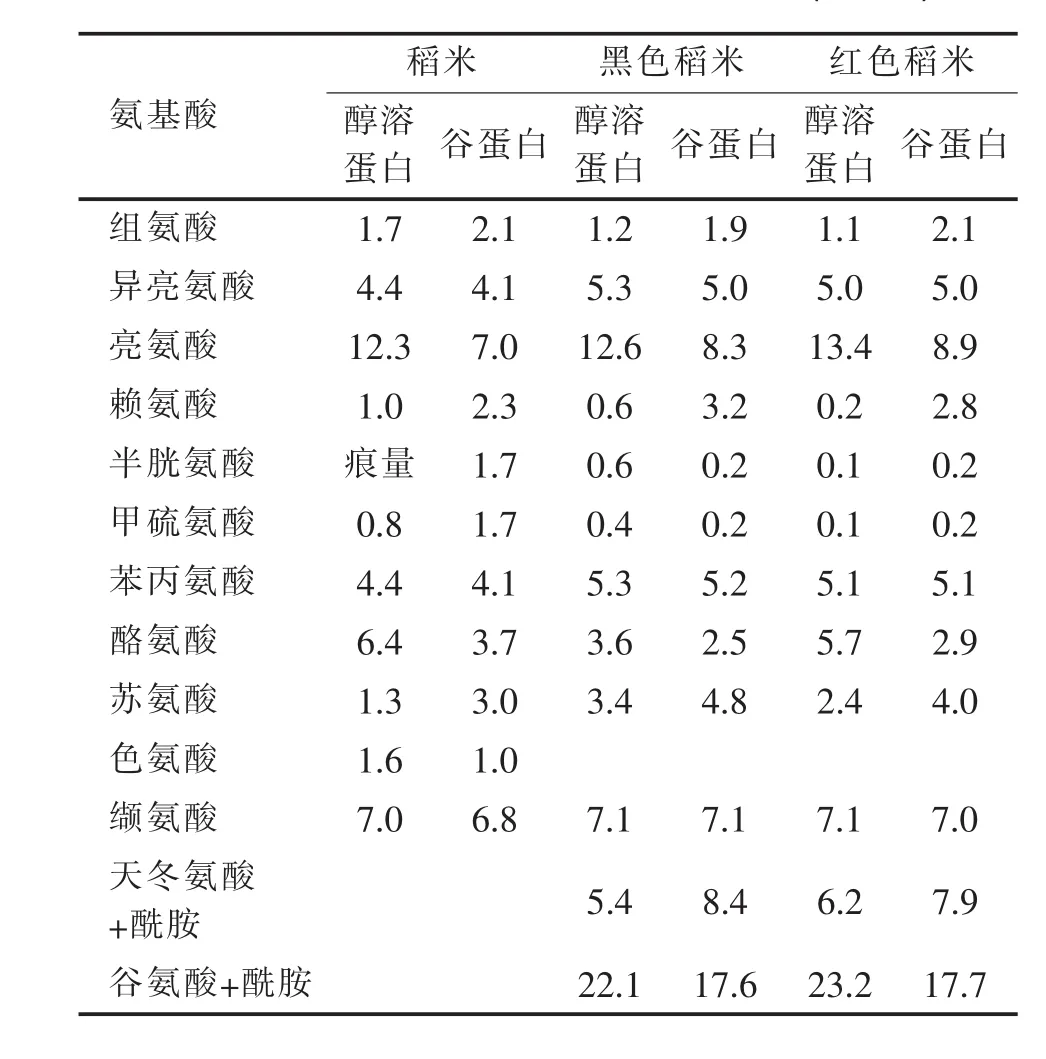
表1 稻谷贮藏蛋白中氨基酸组分含量 (mol%)
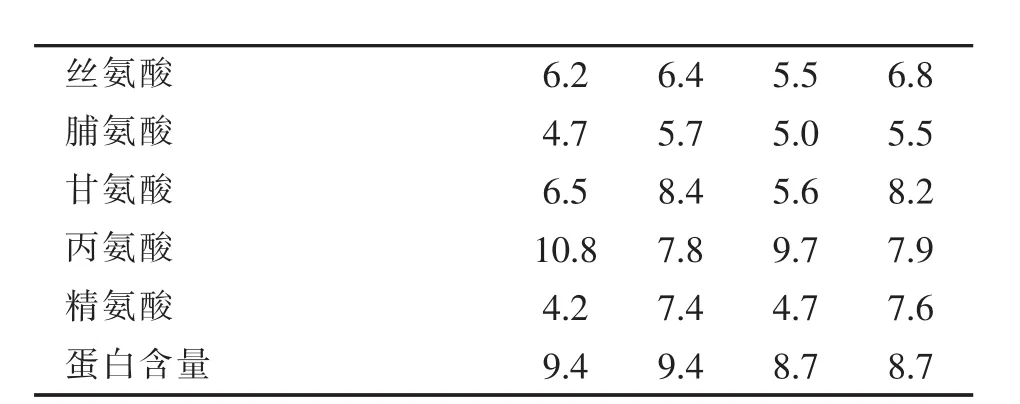
注:稻米数据引自Shew ry(2007);黑粒和红粒古水稻引自Udaka等 (2000)。
Horikoshi等 (1991)表明,日本晴粳稻品种醇溶蛋白含有 131个氨基酸,分子量14930。Ukada等[8]分析表明,古水稻醇溶蛋白中高含量氨基酸有谷氨酸+酰胺、丙氨酸及亮氨酸,低含量氨基酸是赖氨酸、含硫氨基酸。黑粒和红粒古水稻贮藏蛋白之间氨基酸组成没有差异 (表1)。古水稻品种之间给定氨基酸含量最高值与最低值比较,醇溶蛋白比谷蛋白的差异大。在醇溶蛋白中,最高值与最低值差异30%的10种氨基酸是甲硫氨酸、半胱氨酸、赖氨酸、酪氨酸、苏氨酸、脯氨酸、甘氨酸、天冬氨酸+酰胺、甘氨酸、亮氨酸。在谷蛋白中,最高值与最低值差异30%的4种氨基酸有脯氨酸、苏氨酸、酪氨酸及赖氨酸。除过甲硫氨酸和半胱氨酸,其它11个氨基酸平均差异14%。甲硫氨酸和半胱氨酸均占谷蛋白的0.1-0.3 mol%。稻谷醇溶蛋白中的赖氨酸含量,高于其他谷物醇溶蛋白中的赖氨酸含量(Shewry 2007)。
四、贮藏蛋白与稻谷陈化
(一)储藏中稻谷品质劣变
储藏过程中稻谷品质劣变 (陈化)的分子生理机制尚不清楚。稻谷质地特性在收获后数月中发生显著地变化。新收获的大米蒸煮时是粘性、糊状的,储藏几个月后则变得不粘、片状化。这些变化归属稻谷籽粒内发生了多种物理化学变化,大多数这些变化是经验结论。常见大米品种蛋白质含量约7%,且浮动不大,蛋白质含量不是品质的指示指标。韩国主产的六种稻谷品种(Chucheongbyeo,Nampyeongyeo, Ilpumbyeo, Ilmibyeo, OdaebyeoHwayeoungbyeo)在5℃、15℃和25℃储藏12个月后基于储藏温度,在储藏期间稻谷品种间蛋白质含量的变化相似。这个结果与Tran等 (2005)发表的相一致,即五个大米品种在5℃和25℃储藏超过10个月,蛋白质含量没有变化。
大米品质评价中忽略蛋白质结构与类型的影响,由于稻谷蛋白质以蛋白体形式存在,介质蛋白质很少 (Harris and Juliano 1977)。成熟米粒中保持完整的蛋白体影响米饭品质不符合逻辑,由于它们从胚乳成分中分离;可以设想少量的介质蛋白或淀粉粒相关的蛋白影响米饭黏弹性。分析蛋白质在陈化中的作用,许多研究检测巯基(SH) 含量和二硫键的变化[15-16]。Moritaka和Yasumatsu (1972) 采用粉质仪 (Brabender farinograph)测定米粉的稠度,加入氧化剂时米粉稠度增加,在氧化环境中形成二硫键,从游离巯基释放H2S。与40℃储藏比较,5℃储藏稻谷蒸煮米饭香味成分中H2S含量大,即低温延缓稻谷陈化。40℃储藏期间游离巯基减少,二硫键增加,抑制淀粉粒的膨胀,导致米饭不粘、变硬。新收获的稻谷约40%的总半胱氨酸是氧化态 (二硫键),在40℃储藏12个月则60%的总半胱氨酸是氧化态[17]。Hamaker and Griffin (1990)在米饭蒸煮水中加入5 mM DTT显著增加米饭粘度,对中短米粒米饭粘度的影响大于长粒米,在米粉中加入还原剂切开二硫键或蛋白酶,Brabender粘度峰值降低,而分离的大米淀粉不受影响。于是认为大米蛋白的结构影响米饭质地。稻谷收获后前几个月期间,Brabender粘度增加,部分原因是形成二硫键 (Indudhara Swamy等1978)。可见,稻谷陈化期间通过两个半胱氨酸基团的氧化,或通过巯基和二硫键的交换反应,形成二硫键蛋白复合物,限制糊化期间淀粉粒的膨大,这样影响质地。
Chrastil和Zarins (1992)报道,中粒和长粒稻谷储藏期间二硫键数目增加。中粒稻谷的谷蛋白 (Oryzenin)储藏前含有0.2%巯基 (SH),储藏后仅含0.14%SH。长粒稻谷Oryzenin也呈现相似的趋势。储藏过程中两种稻谷低分子量多肽减少,高分子量多肽增加。由储藏引起的Oryzenin多肽亚基分布的这些相对变化,较整个Oryzenin分子平均分子量的变化小 (储藏期间几乎加倍)。多肽亚基成分的这些变化,对储藏期间整个Oryzenin表观分子量的增加起重要作用。大米储藏期间游离氨基酸含量增加,蛋白质的溶解性降低,清蛋白溶解性降低最多 (Zhou等2001)。储藏7年期间,大米的乙酸可溶性蛋白减少(Bolling 1978)。
(二)蛋白含量与米饭质地
早期的研究集中在籽粒主要成分淀粉对大米物理化学特性的影响,尤其是直链淀粉和支链淀粉的长B-主链决定了蒸煮大米的糊化和质地特性。第二大成分蛋白质对米饭食用品质报道较少。在二十世纪60年代,认为大米籽粒成分,而不是淀粉,影响米饭蒸煮的粘性。淀粉体积能膨大64倍,蒸煮米饭不能膨大到这个程度,由于包围淀粉粒的蛋白和细胞壁抑制胚乳的膨胀。Primo等(1962)认为籽粒外围蛋白含量影响米饭质地。与蛋白含量低的大米比较,蛋白含量高的大米米饭硬而难嚼。Oňate等 (1964)分析了3个稻谷品种,每个品种取自蛋白质含量差异的两个试验小区,认为与蛋白含量显著差异相关的感官指标有风味、嫩度及粘性,低蛋白含量的稻米更加有风味、嫩度及粘性。蛋白质含量不影响色泽、香味、直链淀粉含量。Juliano等1965年研究,在同样蒸煮水中,同样品种低蛋白含量的样品较高蛋白含量的样品更嫩、更粘、更有风味。同一品种内样品的直链淀粉含量是基本一样的。品种之间米饭质地的差异不能够以蛋白质含量来解释。Taichung 65品种具有高蛋白含量和中等直链淀粉含量,粘性感官得分低在于其蛋白含量。美国大米品种粗蛋白含量相似,它们之间的质地差异不能完全被蛋白质含量所解释。美国南部中心大米品种蛋白质含量高于加州地区同样品种蛋白质含量,而且质地有差异。蛋白质含量仅部分解释这个现象。直链淀粉含量相似的大米品种糊化和质地特性差别很大 (Champagne等1999)。米粉与大米淀粉amylograph粘度之间的差异说明除过直链淀粉外的其他成分影响米粉蒸煮特性 (Singh等2000)。
米饭蒸煮时间与蛋白含量呈现正相关关系(Chakrabarthy and Dwarakanath 1972)。高蛋白含量大米样品较低蛋白含量大米水分吸收少,较多的蛋白在淀粉粒周围形成一层厚障碍,延缓了淀粉粒吸收水分。
抽穗期间施用氮肥影响蛋白含量和蒸煮大米质地。Yanase等 (1984)对不同程度施用氮肥的16个稻谷品种质地测定表明,高蛋白含量的稻米蒸煮时通常不粘,糙米蛋白含量与大米蒸煮粘性之间的相关系数是-0.561(在5%水平达到显著水平)。Tamaki等 (1989)发现,抽穗期间叶喷硫酸铵 (2 g氮/盆),水不溶蛋白或贮藏蛋白增加了3-4% (蛋白从6%增加到 10%)。利用Foods-Zenken质地测定仪分析,当蛋白含量从6%增加到10%,蒸煮稻米咀嚼度是直线增加的。当蛋白含量从6%增加到8.5%,粘度和质地风味指数(粘度×咀嚼度)增加到最大值,接着随蛋白含量从8.5%增加到10%它们由最大值减少。高蛋白含量的大米蒸煮较难咀嚼、粘性低,而低蛋白含量的大米比中等蛋白含量的大米较软、不粘。
稻谷颖果发育期间米饭质地特性的变化与蛋白质含量相关。Tamaki等 (1989)发现,抽穗后15到50天稻谷蛋白百分率减少约0.75%,蒸煮期间淀粉膨胀受水不溶蛋白、直链淀粉及淀粉脂肪抑制,造成质地差异。当稻谷达到成熟时,蒸煮大米变得更粘、易咀嚼。蛋白质含量通常与蒸煮米粉的粘度负相关,但是它对硬度有正效果[18-19]。认为蛋白质可能通过限制糊化淀粉粒的膨大调控淀粉的膨胀。
(三)米粉糊化和质地
Hamaker和Griffin[20]加入还原剂2-ME到煮饭水切开二硫键,打破蛋白质的结构。对直链淀粉低含量、粘性稻谷,粘度极大增加。当还原剂加入到米粉-水浆中,布氏连续黏度图(Brabender amylograph)曲线较低,当用蛋白酶处理时也类似地较低。Martin和 Fitzgerald[21]报道,采用蛋白酶去除米粉蛋白,RVA曲线轮廓全部降低,沿着曲线所有点粘度降低。米粉中加入DTT切开了蛋白质中二硫键连接降低米粉悬浮液粘度曲线,作用类似蛋白酶处理。Derycke等[19]在米饭蒸煮水中加入DTT,几乎对所有米饭增加粘度、减少硬度。这说明存在蛋白质障碍物和蛋白质的水合作用一起影响米饭粘度曲线,确定了蒸煮米饭的质地。Xie等[22]发现蒸煮水中加入DTT,对蜡质大米米饭粘度显著减少;对非蜡质大米米饭则保持不变;两种米饭的质地变软。这说明二硫键连接的网络蛋白在蒸煮米饭的质地中起重要作用;直链淀粉障碍物的加强是负责米饭粘度的一个重要因素。在高剪切力存在下蛋白结构打破之后观察到较低的粘度,认为是由于膨胀淀粉粒的脆性增加。淀粉粒相关的蛋白转递强度到糊化淀粉颗粒,通过减少直链淀粉分子漏出或者通过物理作用紧密结合淀粉粒。
将还原和非还原条件下测定的米粉糊粘度联系到膨大淀粉粒的溃败,说明蛋白质赋予糊化淀粉粒一定程度的强度或刚性 (Hamaker and Griffin 1993)。在蛋白质结构被打破后,Brabender粘度减少。然而,当以前糊化的米粉糊采用低剪切率的粘度计 (Brookfield Viscometer) 或稠度计(Bostwick Consistometer)测定时,在还原试剂存在下米粉糊粘度增加。这表明,在与淀粉粒结合的正常二硫键蛋白网络缺乏情况下,淀粉粒膨大到较大的程度,产生粘性更大的米粉糊。但是Brabender amylography中存在大量的剪切,膨大的糊化淀粉粒更易溃败,导致米粉糊粘度较低。利 用 流 变 仪 (Carri-Med Controlled Stress Rheometer)试验表明,当还原剂存在时,在较低程度胁迫下糊化米粉糊的结构发生溃败。这些研究表明,特定的蛋白质影响大米粉淀粉粒的糊化特性,这直接相关于全谷物蒸煮米饭的质地特性。需要定性这种特定蛋白质-淀粉的关系。这种特定蛋白负责质地变化,可以通过育种或加工技术调节米饭质地。
(四)贮藏蛋白影响质地特性的机理
了解大米蛋白质如何影响米饭或米粉糊的质地特性,早期的理论解释是模型系统中谷蛋白(glutelin或oryzenin)与淀粉的分子联合,以及淀粉粒联合的蛋白与淀粉凝胶化的关系。
1.淀粉-谷蛋白模式系统:Chrasti (1990)发现,蒸煮米饭的粘性与oryzenin对淀粉、直链淀粉及支链淀粉的结合比率成正相关。利用分离的淀粉、直链淀粉与无淀粉的oryzenin在模式系统中测定 Oryzenin/淀粉摩尔结合比率,与oryzenin吸收光谱比较,在淀粉存在下oryzenin吸收光谱存在差异。中粒和长粒稻谷及其分离的组分,在4℃和40℃储藏 12个月测定蒸煮物的粘度,粘度排序是新收获的中粒大米中最高,其次是储藏在4℃、40℃的大米。这说明谷蛋白对米饭粘性有显著影响,试图了解米饭质地的生化基础及改善方法。
水稻谷蛋白缺乏突变体LGC1显著积累醇溶蛋白,胚乳总蛋白水平与正常稻谷没有差异 (Iida等1993,1997)。Baxter等[23]报道醇溶蛋白显著地影响稻谷米粉的糊化和质地特性。高醇溶蛋白含量相关于显著高度溃败粘度,但是减低米粉形成凝胶的硬度、粘性和胶粘度 (gumminess)。他们还报道清蛋白显著影响稻米粉的糊化和质地特征[24]。稻谷淀粉中存在的清蛋白在蒸煮最初阶段延缓淀粉吸收水分,在后期则加快淀粉水分吸附,含有清蛋白的淀粉样品较纯淀粉样品最终水分吸附多。清蛋白溶于水的本质表明,蛋白质-水-淀粉相互作用影响大米的物理特性。
2.胚乳蛋白对糊化淀粉粒的膨胀起障碍作用。一度认为如其它谷物籽粒,胚乳中连续的蛋白质介质围绕大米淀粉粒形成淀粉粒糊化的障碍(Little and Dawson 1960)。随后稻谷胚乳微结构观察表明,稻谷中的蛋白几乎完全存在于蛋白体,不存在连续的介质蛋白 (Bechtel and Pomeranz 1978)。蒸煮后含有谷蛋白的蛋白体保持原样 (Tanaka等1978),不可能引起质地变化。对大米蛋白-质地关系研究不深入。在煮饭过程中对淀粉糊化抑制的另一因子是,与淀粉粒密切联合的蛋白,它影响颗粒中淀粉糊化的方式,膨胀时淀粉粒如何保持它的形状的方式。Zhou等[17]对37℃储藏的Doongara大米品种采用蛋白酶或纤维素酶处理,显著改变了热特性 (淀粉糊化曲线),在扫描电子显微镜下观察到去除蛋白或打破了细胞壁残留增强了水分穿透,允许淀粉更容易发生糊化作用。
Sano(1984)和Villareal and Juliano (1986)鉴定的60 kDa这种蛋白包埋在淀粉颗粒内,即淀粉粒结合淀粉合成酶 (GBSS)蛋白,在高直链淀粉含量稻谷以较高水平存在。Hamaker和 Griffin[15]表明,用还原试剂2-ME打破该蛋白结构时,米粉糊化程度增加,长粒、非粘性稻米粉较粘性稻米粉糊化程度低,当二硫键被切开时糊化程度增加较大。Hamaker等 (1991)发现这个蛋白与米饭粘度的相关系数类似直链淀粉含量与粘度相关系数。由于蜡质淀粉几乎100%是支链淀粉,糯米淀粉中没有GBSS,就抵抗蛋白酶消化 (Mu-Forster and Wasserman 1998)。GBSS保持非蜡质米饭糊化淀粉粒的刚性,在蛋白酶解后保持糊化淀粉糊的高粘度[22]。Dobo等[25]分析了美国和欧洲水稻种质库中GBSS基因的多态性。GBSS蛋白被单一基因编码,鉴定了3个GBSS等位基因。粳稻存在Wxb等位基因,籼稻存在Wxa等位基因,糯稻存在wx等位基因[12]。WXa等位基因的水稻品种GBSS蛋白是WXb等位基因的水稻品种的3倍多。
也认为淀粉糊化抑制的蛋白以稀疏的介质形态与淀粉粒密切结合,如Greenwell和 Schofield(1986)发现的与小麦淀粉粒周边结合的15 kDa的蛋白,即Friabilin。Friabilin在软麦淀粉较硬麦淀粉丰富, 它具有 3个成分 Puroindoline a (PINA)、Puroindoline b (PINB)和籽粒软化蛋白GSP-1(Rahman等1994)。PINA和PINB的编码基因分别是PinA和PinB(Gautier等1994)。 稻谷不含有PinA和PinB同源框,在转基因水稻M202中表达小麦Pins,增加了稻谷籽粒软度[26]。Wada等[27]采用生物活性珠法将含有硬度基因的节节麦 (Aegilops tauschii,普通小麦D基因组供体)基因组区域导入粳稻品种日本晴,获得了稳定表达Pinb基因的同质转基因水稻。他们近来报道了T4代表达Pinb基因的同质转基因水稻的胚乳细胞超微结构及米粉物理化学特性[28]。非转基因稻谷的复合淀粉粒深埋在介质材料中,而转基因稻谷复合淀粉粒被空隙包围。转基因与不转基因稻谷淀粉粒的形态和大小没有差异。但是转基因大米粉颗粒小、淀粉破坏率高、糊化时粘度低,即Puroindoline b减少稻谷的籽粒硬度。同时发现Puroindoline b在稻谷胚乳复合淀粉粒表面起作用,而非在多边形淀粉粒起作用。
(五)稻谷醇溶蛋白提高人体抗白血病免疫反应
稻谷蛋白是谷物蛋白中营养价值较高者之一,无色、富有必需氨基酸,具有柔和风味、低过敏原及低胆固醇[29]。Komatsu and Hirano (1992)指出大米球蛋白与小麦麦谷蛋白之间氨基酸序列有相似性,但是大米储藏蛋白不含有负责脂泻病的毒性抗原决定基,在营养学上很重要。Liao等[30]报道,从粳稻台湾9号品种胚乳的水提取物,能够刺激从单核细胞释放细胞浆,抑制人白血病细胞的生长,诱导该细胞分化。进一步分析发现,温度稳定的胚乳蛋白质提取物能够增大这种效果,蛋白质组学鉴定的功能性蛋白包括代谢型的、运输、贮藏、抗氧化、发育及抗病蛋白,其中贮藏蛋白最丰富。对4种贮藏蛋白分别分析,醇溶蛋白处理的人外周血液单核细胞培养基中α肿瘤坏死因子的产生增加,在该培养基培养人白血病U937细胞,生长受抑制,并向单核细胞分化。醇溶蛋白的多克隆抗体肯定了这种效果。稻谷醇溶蛋白不能够被小麦醇溶蛋白抗体识别,但是抗白血病活性高于小麦谷蛋白和醇溶蛋白[31]。
五、展望
(一)稻谷贮藏蛋白对米饭质地起重要影响作用
贮藏蛋白如何影响米饭质地的机制还不清楚,可能通过调节蒸煮期间水分扩散到淀粉粒,调控淀粉粒膨胀。贮藏蛋白和淀粉之间的化学相互作用也可能影响米饭质地。已经明确了总蛋白与米饭物理化学特性有关联,探讨了单一蛋白质成分如谷蛋白、GBSS蛋白、醇溶蛋白及清蛋白对米粉糊化和质地特性的影响。这些研究进展揭示了调控米饭质地通过特定蛋白育种,或陈化期间、加工期间修饰它们。利用蛋白质组学和遗传工程方法鉴定影响米饭质地的蛋白。
(二)在发展中国家,稻谷供应20%的膳食蛋白摄取
由于它的氨基酸种类不齐完和微量营养水平有限 (尤其是精米),大米作为大宗食品导致营养不良。具有乳糜泻病的病人已经趋向营养失调,在于面筋不消化导致的免疫反应破坏了小肠的粘膜,减少营养吸收的容量。另外,大多数无面筋产品是低的微量营养素,这增加缺乏的危险性。为了改善大米无面筋产品的营养品质,加入其他蛋白。改变稻米粉的特性,制作类似小麦粉产品的加工产品。以前的研究是通过加入小麦面筋蛋白,或者将高分子量麦谷蛋白亚基 (HMW-GS)结合到稻谷的聚合态蛋白[32],改变大米粉面团的流变学特性。蛋白质加工系统在稻谷和小麦之间是保守的,近来已经成功将小麦HMW-GS编码基因转化水稻[33-34],表明通过遗传工程方法可以改良大米粉面团制作特性。还需要深入了解米粉的蛋白组成与它的面团形成特性之间的关系。
(三)稻谷贮藏蛋白的利用有限,在于它是水不溶的,不能发育粘弹性网络类似结构[35]
通过酸脱酰胺增大米谷蛋白水溶性,加入葡聚糖形成网络类似结构,进而改善大米蛋白的功能特性[36]。由于贮藏蛋白之间共价和非共价相互作用导致的结构特性差异,导致稻谷和小麦面团中水合蛋白复合物的粒度分布是不同的,稻谷中聚合物的长度较小麦中的短,结果是稻谷面团较小麦面团的粘弹性质量小得多。已经成功采用小麦质量评估的面团试验方法定性稻谷粉样品制作面包的流变学特性[37-38]。结合我国国情,定性大米贮藏蛋白与米粉面团、面条加工之间的关系。
(四)稻谷球蛋白与小麦谷蛋白亚基的氨基酸序列之间存在某些相似性
但是稻谷贮藏蛋白不含有导致腹腔疾病的毒性抗原决定基,这从营养角度考虑具有特殊的意义。利用蛋白质组学技术鉴定稻谷贮藏蛋白,提高人体抗疾病免疫反应是令人关注的研究方向。
[1]Shewry PR,Casey R.Seed Proteins[M].Dordrecht, The Netherlands:KluwerAcademic Publishers,1999.
[2]Juliano BO.Ricegrain quality:Problemsand challenges [J].CerealFoodsWorld,1990,35:245.
[3]Perez CM,Juliano BO,De Datta SK,et al.Effect of nitrogen fertilizer treatment and source and season on grain quality of IR64 rice[J].PlantFoods forHuman Nutrition,1990, 40:123.
[4]Komatsu S,Hirano H.Rice seed globulins:a protein similar to wheat seed glutelin [J].Phytochemistry,1992,31: 3455-3459.
[5]Li X,Okita TW.Accumulation of prolamins and glutelins during rice seed development:a quantitative evaluation [J].Plantand CellPhysiology,1993,34:358-390.
[6]Hamada JS.Separation andmolecularmassdistribution of rice proteins by size-exclusion high-performance liquid chromatography in a dissociating buffer [J].Journal of Chromatography A,1996,734:195-203.
[7]Katsube-Tanaka T,Duldulao JBA,Kimura Y,etal.The two subfamilies of rice glutelin differ in both primary and highorderstructure[J].BiochimicaetBiophysica Acta,2004,1699: 95-102.
[8]Udaka J,Koga T,TsujiH,etal.Efficientextraction and some properties of storage proteins(Prolamin and glutelin)in ancient rice cultivars[J].Journal of Nutritional Science and Vitaminology,2000,46:84-90.
[9]Ogawa N.Fundamental studies on characterization of red rice[J].Shokuseikatsu-kenkyu,1993,14(4)4-10.
[10]CaoXH,WenHB,LiCJ,GuZX.Differencesin functional properties and biochemical characteristics of congenetic rice proteins[J].JournalofCerealScience,2009,50:184-189.
[11]Villareal CP,Juliano BO.Comparative levels ofwaxy gene product of endosperm starch granules of different rice ecotypes[J].Starch,1989,41:369.
[12]Hirano HY,Sano Y.Molecular characterization of the waxy locus of rice (Oryza sativa L.) [J].Plant and Cell Physiology,1991,32:989-997.
[13]Hamada JS,Spanier AM,Bland JM,etal.Preparative separation of value-added peptides from rice bran proteins by high-performance liquid chromatography [J].Journal of Chromatography A,1998,827:319-327
[14]Van Der BorghtAVD,Vandeputte GE,Derycke V,et al.Extractability and chromatographic separation of rice endosperm proteins[J].Journal of Cereal Science,2006,44: 68-74.
[15]HamakerBR.GriffinVK.Effectofdisulfidebondcontaining protein on rice starch gelatinization and pasting [J].Cereal Chemistry,1993,70:377-380.
[16]Zhou ZK,RobardsK,HelliwellS,Blanchard C.Effect of storage temperature on rice properties.Food Research International,2010,43:709-715.
[17]Chrastil J.Protein-starch interactions in rice grains. Influence of storage on oryzenin and starch [J].Journal of Agriculturaland Food Chemistry,1990,38:1804.
[18]Lyon BG,Champagne ET,Vinyard BT,etal.Sensory and instrumental relationships of texture of cooked rice from selected cultivarsand postharvesthandlingpractices[J].Cereal Chemistry,2000,77:64-69.
[19]Derycke V,Veraverbeke WS,Vandeputte GE,et al. Impact of proteins on pasting and cooking properties of nonparboiled and parboiled rice [J].Cereal Chemistry,2005,82:468-474.
[20]Hamaker BR,Griffin VK.Changing the viscoelastic propertiesofcooked rice through protein disruption [J].Cereal Chemistry,1990,67:261.
[21]MartinM,FitzgeraldMA.Proteinsonricegrainsinfluence cooking properties [J].Journal of Cereal Science,2002,36: 285-294.
[22]Xie L,Chen N,Duan B,et al.Impact of proteins on pasting and cooking propertiesofwaxy and non-waxy rice[J]. JournalofCerealScience,2008,47:372-379.
[23]Baxter G,Blanchard C,Zhao J.Effectof prolamin on rice (Oryza sativa L.)textural and pasting propterties [J]. JournalofCerealScience,2004,40:205-211.
[24]Baxter G,Zhao J,Blanchard C.Albumin significantly afftects pasting and textural characteristics of rice flour[J]. CerealChemistry,2010,87(3):250-255.
[25]Dobo M,Ayres N,Walker G,etal.Polymorphism in the GBSSgene affectsamylase content in USand European rice germplasm [J].JournalofCerealScience,2010,52:450-456.
[26]Krishnamurthy K,Giroux MJ.Expression of wheat puroindoline genes in transgenic rice enhances grain softness [J].Nature Biotechnology,2001,19:162-166.
[27]Wada N,Kajiyama S,Akiyama Y,et al.Bioactive beads-mediated tansformation of ricewith large DNA fragments containing Aegilpos tauschii genes [J].Plant Cell Reports, 2009,28:759-768.
[28]Wada N,Kajiyama S,Cartagena JA,etal.The effects of puroindoline b on the ultrastructure of endosperm cells and physicochemical properties of transgenic rice plants [J]. JournalofCerealScience,2010,51:182-188.
[29]Chrastil J.Correlationsbetween and physicochemical and functionalpropertiesof rice[J].JournalofAgriculturaland Food Chemistry,1992,40:1683-1686.
[30]Liao HF,Chen YY,Yang YC,et al.Rice (Oryza sativa L.)inhibits growth and induce differentiation of human leukemic U937 cells through activation of peripheral blood mononuclear cells[J].Food and Chemical Toxicology,2006, 44(10):1724-1729
[31]Chen YJ,Chen YY,Wu CT,et al.Prolamin,a rice protein,augments anti-leukaemia immune response [J]. JournalofCerealScience,2010,51:189-197.
[32]Oszvald M,TomoskoziS,Larroque O,KeresztenyiE, Tamas L,Bekes F.Characterization of rice storage proteins by SE-HPLC and micro Z-arm mixer[J].Journal of Cereal Science,2008,48:68-76.
[33]Oszvald M,Jenes B,TomoskoziS,etal.Expression of the 1Dx5 high molecular weight glutenin subunit protein in transgenic rice[J].CerealResearch Communication,2007,35: 1543-1549.
[34]ArakiE,Ikeda TM,Ohgihara Y,etal.Developmentof transgenic rice (Oryza sativa L.)expressing wheat high-and low-molecular-weightglutenin subunitproteins[J].Breeding Science,2008,58:121-128.
[35]MarcoC,RosellCM.Effectofdifferentprotein isolates and transglutaminase on rice flour properties [J].Journal of Food Engineering,2008,84:132-139.
[36]Li XH,Liu YL,Yi CP,et al.Microstructure and rheological properties of mixtures of acid-deamidated rice protein and dextran [J].Journal of Cereal Science,2010, 51:7-12.
[37]Sivaramakrishman HP,Senge B,Chattopadhyay PK. Rheologicalpropertiesof rice dough formaking rice bread [J]. JournalofFood Engineering,2004,62:37-45
[38]Gujral HS,Rosell CM.Improvementofbreadmaking quality of rice flour by glucose oxidase [J].Food Research International,2004,37:75-81.
(作者分别为:国家粮食局科学研究院,吉林大学食品科学与工程学院)
国家教育部留学归国启动基金 (CZ1008)
李兴军,男,1971年出生,副研究员,博士,粮食生化与多糖工程
