小麦蛋白胨促进乳酸菌增殖的研究
2017-04-26孙文敬辛晓亚郭文杰崔凤杰方颂平朱本国冯岳琴夏统前
孙文敬,辛晓亚,郭文杰,崔凤杰*,方颂平,朱本国,冯岳琴,夏统前
1(江苏大学 食品与生物工程学院,江苏 镇江,212013)2(安徽瑞福祥食品有限公司,安徽 亳州,236800)
小麦蛋白胨促进乳酸菌增殖的研究
孙文敬1,辛晓亚1,郭文杰2,崔凤杰1*,方颂平2,朱本国2,冯岳琴1,夏统前1
1(江苏大学 食品与生物工程学院,江苏 镇江,212013)2(安徽瑞福祥食品有限公司,安徽 亳州,236800)
对比分析了制备的小麦蛋白胨(WP)与市售蛋白胨的理化性质及其部分(全部)替代发酵氮源对乳酸菌增殖的影响。结果表明,该小麦蛋白胨理化性质已基本达到市售蛋白胨的质量标准要求。小麦蛋白胨所含的非游离氨基酸大分子量(Mw)肽段(>500 u)的比例高于其他市售植物蛋白胨,但其有利于乳酸乳球菌、干酪乳杆菌、罗伊氏乳杆菌和植物乳杆菌等增殖,活菌数分别达到9.64×108CFU/mL,2.30×109CFU/mL,1.60×109CFU/mL和1.14×109CFU/mL。对比发酵前后肽段分子质量分布发现,干酪乳杆菌可能主要吸收和/或降解小麦蛋白胨中分子量大于180 u的肽段为氮源供其生长。
小麦蛋白胨;氮源;乳酸菌;培养基;增殖
蛋白胨是一种以蛋白质为底物、经蛋白酶水解得到的水溶性混合物,主要是由胨、肽和氨基酸组成[1]。作为微生物培养基的基础成分,蛋白胨可以提供大部分微生物生长和发育所需要的氮源[2],已广泛用于发酵工业和医药工业,也作为功能性食品和化妆品的配料应用于食品或化妆品等生产[3]。蛋白胨从来源上可分为动物源、植物源和微生物蛋白胨。传统发酵工业主要采用酪蛋白为来源的蛋白胨进行日常生产。但动物源蛋白胨缺点明显,如存在潜在的病毒污染、成分不明确和不利于产物纯化等。因此,疫苗、抗体和干扰素等生物制药领域对动物源蛋白胨的使用安全顾虑逐渐增加[4]。而植物和微生物源蛋白胨具有无污染和高安全性等优点,现已成为生物制药领域的优先选择。
目前,国内外诸多研究集中在利用酶法水解玉米蛋白、大豆蛋白、菜籽饼粕等植物蛋白制备抗氧化肽、降压肽和抗菌肽等功能肽。而开发和利用植物蛋白生产发酵氮源及其性能评价的系统研究仍相对较少。如李润娇等将花生粕酶解制备花生蛋白胨,并用于乳酸菌的培养[5];莫芬等酶解小麦面筋蛋白得到了不同水解度产物,其具有显著影响酿酒酵母的增殖及风味物质组成的作用[6];而白凤翎等发现,分子质量小于5 ku的大豆蛋白水解物具有明显的促进酸奶乳酸菌增殖效果[7]。国外已有较多将植物蛋白水解液应用于蛋白药物生产和疫苗制备等方面的研究。如KIM等采用大豆蛋白水解物、小麦蛋白水解物及酵母提取物三者的复合物培养悬浮型重组CHO 细胞,以促进抗体合成[8]。FALLAH研究发现酶解鲢鱼加工副产物制备的鱼蛋白胨可作为有效氮源用于培养金黄色葡萄球菌[9]。BENEDINI等证实,将大豆蛋白胨替代兽疫链球菌的传统脑心浸液(Brain-Heart Infusion)和羊血培养基,菌体生长良好,未造成菌体生长损伤和代谢偏转[10]。
我国拥有丰富的小麦蛋白资源。作为小麦淀粉加工副产品,小麦蛋白具有较高的营养价值,但由于其溶解性较差而限制了其在食品发酵工业中的应用。近年来,开发小麦蛋白新用途,实现其高值化应用是研究的热点之一。目前国内外有关小麦蛋白的研究多集中在其活性多肽的酶法制备和功能评价等方面。本课题组前期已较为系统地开展了酶法水解小麦蛋白生产抗氧化肽的研究,包括筛选蛋白酶、优化其水解条件以及解析小麦蛋白肽结构等[11-13]。然而,有关小麦蛋白水解物作为氮源应用于微生物菌体生长和产物合成的研究鲜有报道[6]。本项目在前期研究的基础上,拟对比分析制备的小麦蛋白胨与市售蛋白胨的理化性质,考察研究小麦蛋白胨(部分)替代市售蛋白胨等氮源培养乳酸菌的性能,进而通过分析其肽段分布和氨基酸组成,探讨小麦蛋白胨促进乳酸菌增殖的可能机制,为扩大小麦蛋白胨应用领域提供了一定的理论指导。
1 材料与方法
1.1 材料
1.1.1 小麦蛋白胨
小麦蛋白胨WP:小麦蛋白(蛋白含量73.37%,安徽瑞福祥食品有限公司由经中性蛋白酶(1.0×106U/g,南宁东恒华道生物科技有限公司)和胰蛋白酶(2.5×106U/g,上海生工科技有限公司)酶解后[12],经板框过滤、喷雾干燥制成的淡黄色粉末状样品。
小麦蛋白胨E1(ORG E1)购自法国Organotechnie公司;Oxoid NO.1植物蛋白胨购自英国Oxoid公司;Fluka 93452小麦蛋白胨购自Sigma公司;麦麸蛋白胨Hypep 4601N购自美国Kerry 公司。
牛肉膏、酵母膏、胰蛋白胨、葡萄糖、乙酸钠、柠檬酸氢二铵、吐温-80、K2HPO4·3H2O、MgSO4·7H2O 和MnSO4·H2O等购自国药集团化学试剂有限公司。
1.1.2 菌株来源
乳酸乳球菌乳酸亚种(Lactococcuslactissubsp.lactis)CICC 6242,购自中国工业微生物菌种保藏管理中心;罗伊氏乳杆菌(Lactobacillusreuteri)和植物乳杆菌(Lactobacillusplantarum),由加拿大西安大略大学和加拿大益生菌研发中心GREGOR Reid教授提供。干酪乳杆菌(Lactobacilluscasei),由本实验室采用常规乳酸菌分离方法获得并保存。
1.2 培养基
改良MRS液体培养基 (g/L):胰蛋白胨 10,牛肉膏 10,酵母膏5,K2HPO4·3H2O2,乙酸钠5,葡萄糖 20,吐温80 1 mL,柠檬酸氢二铵2,MgSO4·7H2O 0.58,MnSO4·H2O 0.25,pH6.5,121 ℃灭菌20 min。
MRS固体培养基:在液体培养基的基础上加入15%的琼脂。
部分替代发酵培养基 (g/L):将改良MRS培养基中胰蛋白胨替换成制备的小麦蛋白胨或市售蛋白胨,其他组成及含量均不变。
完全替代发酵培养基 (g/L):将改良MRS培养基中胰蛋白胨、牛肉膏和酵母膏全部替换成制备的小麦蛋白胨或市售蛋白胨,添加量为20 g/L,其他组成及含量均不变。
1.3 乳酸菌培养
将各乳酸菌菌株接入MRS培养基于37℃下活化12 h后,以3% (v/v)接种量接入装有 60 mL种子培养基的100 mL三角瓶中,37℃静置培养12 h后制成发酵种子液。
分装部分替代或完全替代发酵培养基60 mL/100 mL三角瓶,121 ℃灭菌20 min后,接入3% (v/v) 种子液,37 ℃静置培养12 h后,分别测度乳酸菌增殖性能(OD600nm值,活菌数和菌体干重)。
1.4 乳酸菌增殖性能测定
OD600nm测定:以空白培养基调零,分光光度计于600 nm下测定发酵液OD值。
活菌计数:采用平板菌落计数法测定发酵液中乳酸菌的活菌数,发酵终止后,取1mL发酵液,以无菌水稀释成10-1~10-7等7个梯度,选取适宜梯度0.1 mL涂布于平板上,每个样品重复3次,37 ℃培养48 h,选取菌落数在 30~300 之间的平板进行菌落计数,活菌数用CFU/mL 表示。
菌体干重:取25 mL发酵液,转速8 000 r/min离心10 min,取沉淀烘干至恒重后测菌体干重。
1.5 蛋白胨样品理化性质测定
准确称取各蛋白胨样品,配制成质量浓度为20 g/L的溶液,观察其溶解性、溶液外观色泽等,并测定其pH值;采用GB 5009.5—2010凯氏定氮法测定其总氮含量,并计算其蛋白含量;采用DNS法测定其总糖含量。
1.6 蛋白胨样品中氨基酸种类和含量的测定
取一定量蛋白胨样品于试管中,分别加入10 mL 6 moL/L的HCl溶液,置110 ℃水解24 h后过滤定容至50 mL;取滤液0.5 mL蒸干,加入1 mL样品稀释液充分溶解,用氨基酸自动分析仪测定。
1.7 蛋白胨样品中肽分子量分布的测定[6]
采用高效液相色谱法测定不同来源蛋白胨中肽段分子量分布。色谱条件为:(a)色谱柱:TSKgel 2000 SWXL 300 mm×7.8 mm;柱温:30 ℃;(b)流动相:乙腈/水/三氟乙酸,45/55/0.1(v/v);流速:0.5 mL/min;进样量:20 μL;洗脱时间:20 min;(c)检测波长:UV 220nm。
1.8 数据分析
实验设计为每样3个重复。结果采用SPSS10.0软件进行处理,为3个重复的平均值±标准偏差。采用one way ANOVA进行试验数据的方差分析,P<0.05 表明结果具有显著性差异。
2 结果与分析
2.1 不同来源蛋白胨理化性质分析
基于前期研究结果,以小麦蛋白为原料,采用中性蛋白酶和胰蛋白酶水解,酶解液经板框过滤取清夜,喷雾干燥制成淡黄色粉末状的小麦蛋白胨样品。表1汇总了制备的小麦蛋白胨WP与其他不同来源蛋白胨的理化性质。
由表1可知,小麦蛋白胨WP中蛋白含量为74.56%,高于Oxoid(73.76%)和Fluka 93452(67.02%)。小麦蛋白胨WP还含有少量糖(10.60%),其溶解性能良好,溶液(2%,w/v)外观为淡黄色,澄清透明。由此可知,该小麦蛋白胨WP的理化性质已基本达到市售蛋白胨的质量标准要求。
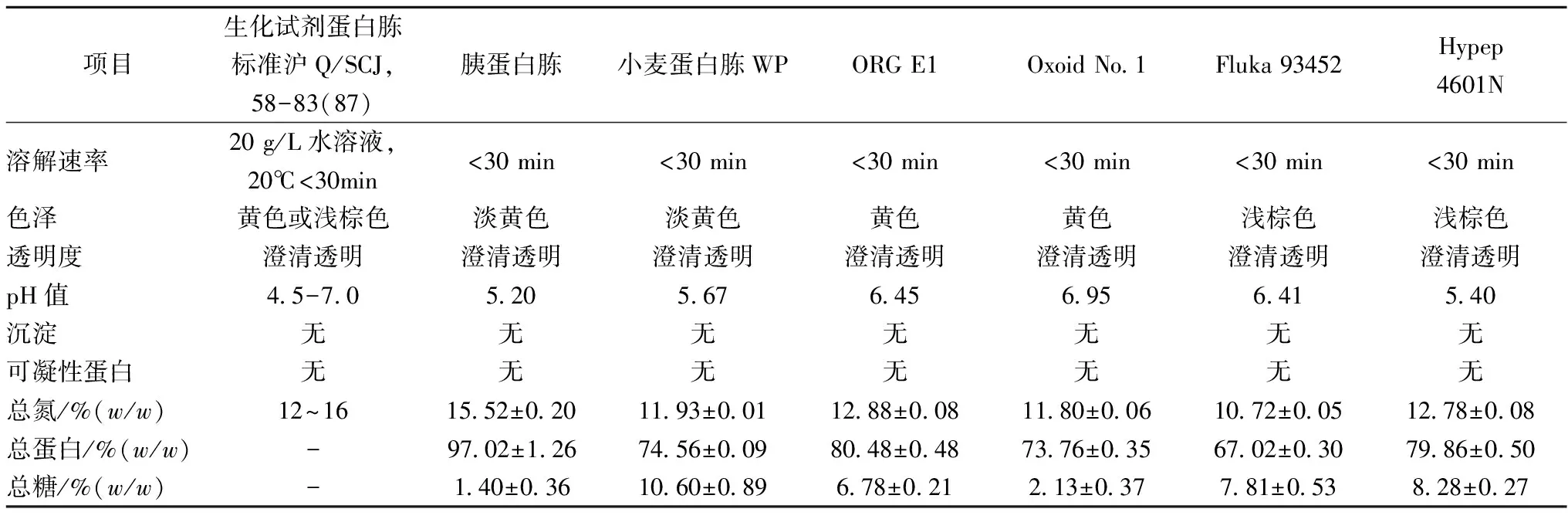
表1 不同来源蛋白胨常规理化性质
2.2 不同来源蛋白胨中氨基酸组成分析
不同来源蛋白胨氨基酸组成及含量分析见表2,结果表明,小麦蛋白胨WP的氨基酸种类丰富,与其他几种市售试剂级蛋白胨一样,含有17种氨基酸,总量达 71.7%,仅低于胰蛋白胨(85.9%)和Hypep 4601N(79.4%)。
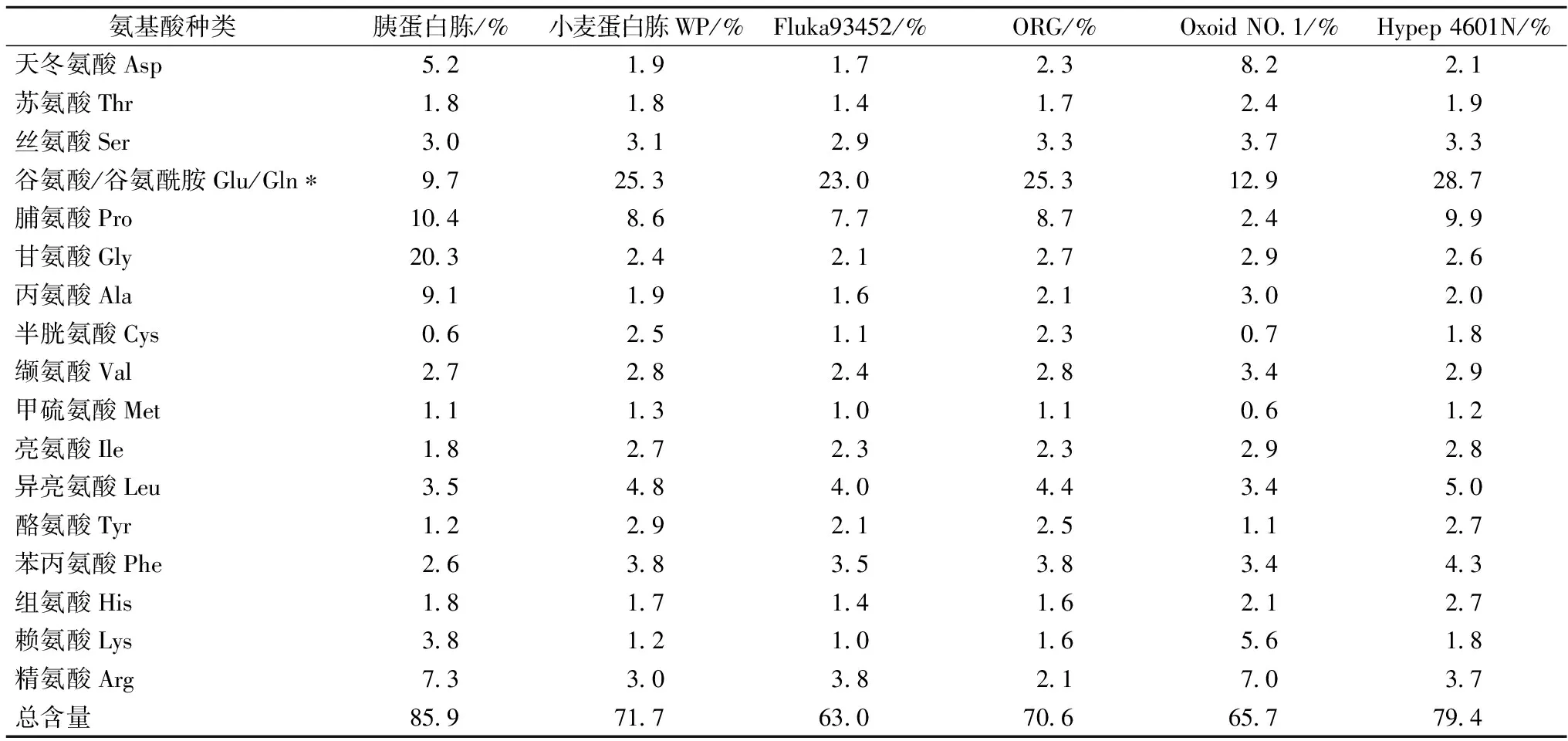
表2 不同来源蛋白胨中氨基酸组成分析
注:*酸水解过程中谷氨酰胺被变成谷氨酸。
表3归纳了6种蛋白胨样品中含量较高的6种氨基酸种类和比例。各蛋白胨均含有较高量的谷氨酸/谷氨酰胺、脯氨酸和精氨酸;苯丙氨酸、异亮氨酸、丝氨酸的含量也相对较高。在植物源蛋白胨中,谷氨酸的含量均最高,如小麦蛋白胨WP、Fluka 93452、ORG E1和Hypep 4601N中谷氨酸含量分别达到为25.3%、23.0%、25.3%和28.7%,明显高于胰蛋白胨。一般认为,氨基酸形式的氮源有利于提高微生物的生长和发酵产率[14]。然而,白凤翎等研究发现,保加利亚乳杆菌和嗜热链球菌等乳酸菌以大豆蛋白水解物为氮源的生长效果明显优于仅添加天冬酰胺、谷氨酰胺、谷氨酸等20种氨基酸培养基,可能主要是寡肽而非游离氨基酸显著影响乳酸菌的生长[7]。本研究制备的小麦蛋白胨富含谷氨酸/谷氨酰胺、脯氨酸和异亮氨酸等,有望为乳酸菌、酵母菌和丝状真菌等菌体生长和产物合成提供有效氮源。
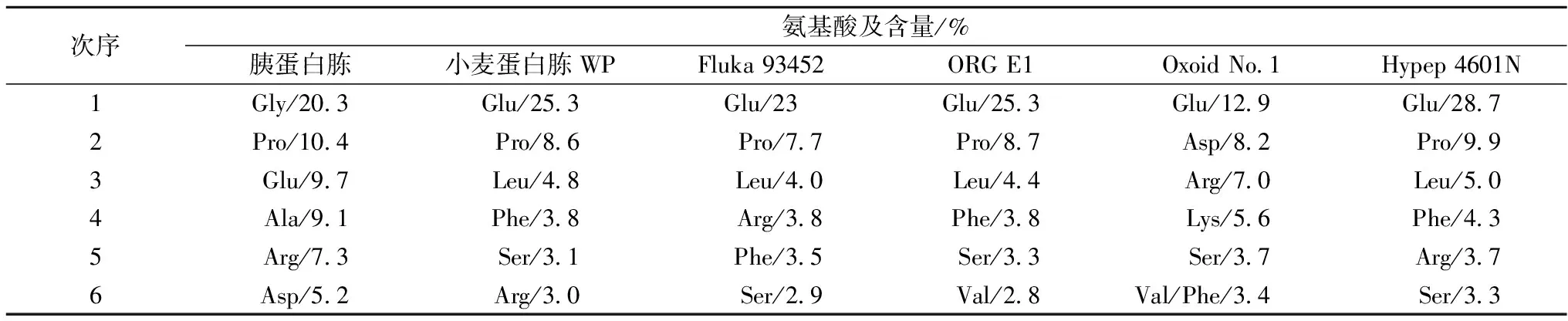
表3 不同蛋白胨中含量较高的氨基酸
2.3 小麦蛋白胨作为替代氮源对乳酸菌生长性能的影响
为评价制备的小麦蛋白胨WP作为发酵氮源的可行性,将其替代乳酸菌生长培养基(MRS)的蛋白胨,对比考察其对罗伊氏乳杆菌、保加利亚乳杆菌、植物乳杆菌及干酪乳杆菌等乳酸菌生长性能的影响。结果如图1所示。
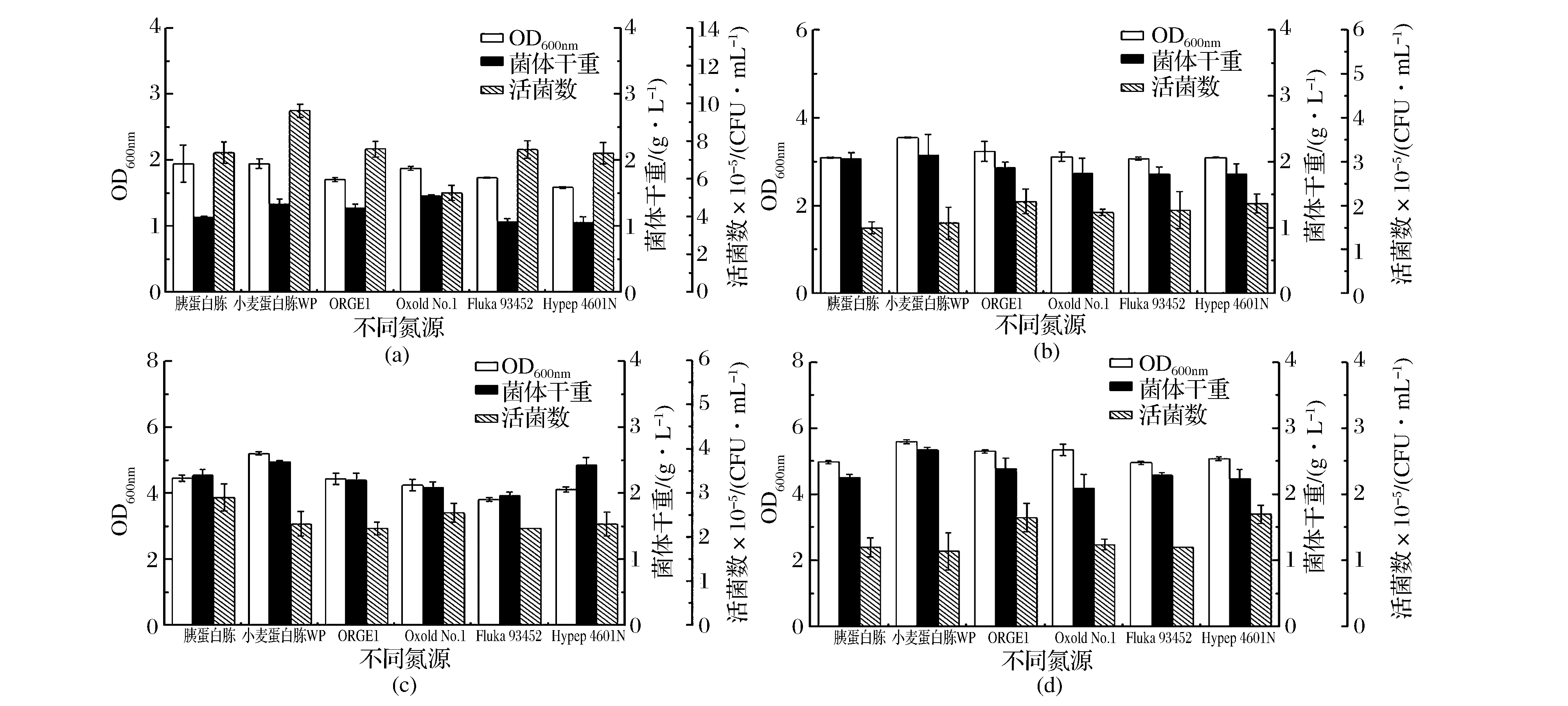
(a)乳酸乳球菌; (b)干酪乳杆菌;(c)罗伊氏乳杆菌;(d)植物乳杆菌(a)Lactococcus lactis subsp lactis(b)Lactobacillus casei(c)Lactobacillus reuteri(d)Lactobacillus plantarum图1 不同蛋白胨对乳酸菌生长量的影响Fig.1 Effect ofpeptones sourced from various companies on the growth of lactic acid bacteria
由图1可知,乳酸乳球菌可利用6种蛋白胨进行生长(图1(a)),但不同来源的蛋白胨对其增殖影响显著。其中,小麦蛋白胨WP替代MRS中蛋白胨发酵12 h后,乳酸乳球菌生长性能达到最大,其OD600nm值、菌体干重和活菌数分别为1.950、1.340 g/L和9.64×108CFU/mL。其次为ORG E1和Fluka 93452。当以小麦蛋白胨WP为替代氮源用于培养干酪乳杆菌、罗伊氏乳杆菌和植物乳杆菌,经12 h发酵后,各菌株生长性能均优于原MRS培养基。与其他市售植物蛋白胨相比,小麦蛋白胨WP实验组的OD600nm值和菌体干重均为最高;但其活菌数略低于其他市售植物蛋白胨,约为1.14×109~2.30×109CFU/mL(图1(b-d))。
2.4 乳酸菌对肽段吸收情况的研究
为研究乳酸菌吸收和利用测试蛋白胨中肽段或氨基酸的模式和可能的机理,将小麦蛋白胨WP和其他市售蛋白胨全部替代MRS培养基中氮源(蛋白胨、酵母膏和牛肉膏)进行培养干酪乳杆菌,对比分析发酵12 h前后各发酵醪中肽段分子量分布及其含量。结果如图2和表4所示。
由图2可知,当各蛋白胨样品完全替代MRS培养基中氮源时,干酪乳杆菌的菌体OD600nm值均在1.0左右,显著低于MRS培养基或部分氮源替代培养基,表明原MRS培养基中酵母膏和牛肉膏等氮源对其生长也起到促进作用。当小麦蛋白胨WP完全替代MRS培养基中的氮源时,干酪乳酸菌表现出良好的生长性能,其OD600nm值、菌体干重和活菌数分别为0.915,0.948 g/L,和2.31×109CFU/mL,高于Oxoid NO.1和Hypep 4601N组。

图2 不同蛋白胨对干酪乳杆菌生长量的影响Fig.2 Effect ofpeptones sourced from various companies on the growth of Lactobacillus casei
蛋白质经过水解会得到大小不同的多肽片段。蛋白质原料、蛋白酶的种类以及酶解条件均会显著影响原料的水解效率和产物品质。表4为不同来源蛋白胨经干酪乳杆菌发酵12 h前后肽段的分子量分布。由表4可知,当发酵0 h时,5种植物源蛋白胨中,Fluka 93452、ORG 、Oxoid No.1和Hypep 4601N的肽段分子量均较低,分子量>5 000 u的肽段含量很低,几乎为0,小于1 000 u的肽段比例为95%以上,而小于500 u的肽段比例为均高于85%,特别是分子量小于180 u的游离氨基酸含量均高于51%,为小麦蛋白胨中游离氨基酸含量的2倍,这表明市售的国外品牌的植物蛋白胨产品的分子量均较低。小麦蛋白胨WP中大分子量肽段含量相对其他蛋白胨较高,其中,分子量>5 000 u的肽含量为0.96%,分子量在3 000~5 000 u的肽含量也略高,是其他植物源蛋白胨的3~10倍,小于3 000 u的小分子肽段,即分子量为500~1 000 u、1 000~2 000 u及2 000~3 000 u的肽段含量均明显高于其他几种植物源蛋白胨。
微生物会选择性吸收蛋白胨中不同分子量(Mw)肽段促进其生长和发酵。由表4可以看出,经干酪乳杆菌发酵12 h后,小麦蛋白胨WP中分子量大于180 u以上的肽段相对含量均有不同程度的降低,而分子量低于180 u的游离氨基酸相对含量则有较大幅度的增加,提示干酪乳杆菌可能主要吸收和/或降解小麦蛋白胨WP中分子量大于180 u的肽段为氮源供其生长。
干酪乳杆菌利用4种市售植物蛋白胨中肽段的模式与小麦蛋白胨基本一致。一般认为,微生物会选择性吸收蛋白胨中不同分子量的肽段促进其生长和发酵。如MO等发现,酿酒酵母在发酵过程中,几乎不吸收利用分子量大于10 ku 和 5~10 ku的肽段(利用率小于7%),而主要利用50%以上的分子量小于1 ku的肽段[15]。在本研究中,虽然制备的小麦蛋白胨WP中分子量小于500 u的肽段含量较低,但并未影响乳酸菌的增殖,这可能与微生物的种类和发酵产物有关。如刘定杭等采用不同酶解条件制备出含有不同分子量肽段的牛骨蛋白胨,其对副干酪乳杆菌生长和发酵产物抑菌效果影响不大,而含低分子量肽段的蛋白胨样品可提高低温蛋白酶的酶活[16]。ZHANG等也发现分子量小于3 ku的酪蛋白活性肽对乳酸菌有明显促生长活性,并能显著减少酸奶发酵时间,提高成品酸奶中乳酸菌的数量[17]。而乳酸菌主要是依赖于降解蛋白质和多肽来满足细胞合成代谢对氨基酸的需求。如MCKAY等证实,乳酸乳球菌必须依靠自身蛋白水解系统降解乳中酪蛋白来维持其生长需要[18]。该蛋白水解系统主要包括将大分子酪蛋白水解成多肽的胞外蛋白酶、将多肽转运进入细胞的转运系统和将多肽水解形成游离氨基酸的肽酶[19]。如,乳酸乳球菌的寡肽转运系统Opp主要转运含有4~11个氨基酸残基的肽段(分子量约为500~1 000 u)[20],而在小麦蛋白胨WP培养基中,分子量为500~1 000 u的肽段含量为19.04%,因此,乳酸菌完全可以将其转运至胞内进一步水解和利用,实现其增殖。
3 结论
本项目以小麦蛋白为原料制备得到的淡黄色粉末状小麦蛋白胨WP,其蛋白含量为74.56%,溶解性能良好,溶液外观为淡黄色,澄清透亮。完全(部分)替代氮源发酵实验结果表明,小麦蛋白胨WP可作为氮源提供干酪乳杆菌等乳酸菌生长必需的寡肽和氨基酸,且菌体主要利用WP所含的分子量大于180 u的肽段实现其快速增殖。综上,制备的小麦蛋白胨具有良好的理化品质,并可为乳酸菌生长提供充分的氮源。后续将通过改进生产工艺进一步降低大分子肽段的比例,深入探讨其促进乳酸菌等微生物增殖和产物合成的相关机理,为开发高品质小麦蛋白胨提供一定的研究基础。

表4 不同培养基中的肽的分子量分布
[1] CHAPMAN M, MARIANO K, MACREADIE I. Lupin peptone as a replacement for animal-derived peptone in rich culture media for yeast [J].Journal of Microbiological Methods, 2015, 109:39-40. DOI: 10.1016/j.mimet.2014.12.005.
[2] YAO WY, WU X, ZHU J, et al.Utilization of protein extract from dairy manure as a nitrogen source byRhizopusoryzaeNRRL-395 for L-lactic acid production[J]. Bioresource Technology, 2010, 101(1):4132-4138. DOI:10.1016/j.biortech.2010.01.004.
[3] 张萍. 酶解罗非鱼碎肉制备生化试剂蛋白陈的工艺研究[D].福州:福建农林大学,2009.
[4] DAVAMI F, VALDI L, RAJENDRA Y, M WURM F. Peptone supplementation of culture medium has variable effects on the productivity of CHO cells[J].International Journal of Molecular Medicine,2014,3(3):146-156.
[5] 李润娇.花生粕制备生化试剂蛋白胨的工艺研究[D].济南:山东师范大学,2014.
[6] 莫芬,赵谋明,赵海锋.小麦面筋蛋白水解物对酿酒酵母增殖和发酵性能的影响[J].食品工业科技,2012,33(22):222-225.
[7] 白凤翎.蛋白水解物促乳酸菌增殖及高密度培养体系研究[D].北京:北京林业大学,2010.
[8] KIM S H, LEE G M.Development of serum-free medium supplemented with hydrolysates for the production of therapeutic antibodies in CHO cell cultures using design of experiments[J].Applied Microbiology and Biotechnology,2009,83(4):639-648.DOI: 10.1007/s00253-009-1903-1.
[9] FALLAH M, BAHRAM S, JAVADIAN S R. Fish peptone development using enzymatic hydrolysis of silver carp by-products as a nitrogen source inStaphylococcusaureusmedia[J].Food Science & Nutrition,2015,3(2):153-157.DOI:10.1002/fsn3.198.
[10] BENEDINI L J, SANTANA M H. Effects of soy peptone on the inoculum preparation ofStreptococcuszooepidemicusfor production of hyaluronic acid[J].Bioresource Technology,2013,130:798-800.doi:10.1016/j.biortech.2012.12.161.
[11] 崔凤杰,闫桂强,黄达明,等.酶法降解小麦面筋蛋白制备抗氧化产物的研究[J].食品研究与开发,2007,28(11):81-84.
[12] 崔凤杰,孙文敬,张玲,等.小麦面筋多肽D-WG-0-Ⅰ的分离纯化与结构鉴定[J].食品科技,2013,38(4):164-168.
[13] 林琳,张玲,崔凤杰,等.双酶法水解小麦面筋蛋白提高产物抗氧化活性[J].食品科技,2009,34(12):186-190.
[14] 刘政,杨绍斌,宋小美,等.氨基酸类营养对微生物生长及活性的影响[J].黑龙江农业科学,2010(5):13-15.
[15] MO F, ZHAO H, LEI, H et al.Effects of nitrogen composition on fermentation performance of brewer's yeast and the absorption of peptides with different molecular weights[J].Applied Biochemistry and Biotechnology,2013,171(6):1339-1350.DOI: 10.1007/s12010-013-0434-5.
[16] 刘定杭.牛骨蛋白胨中多肽分布及对工业菌株发酵的影响[D].哈尔滨:黑龙江大学,2011.
[17] ZHANG Q,REN J,ZHAO M,et al. Isolation and characterization of three novel peptides from casein hydrolysates that stimulate the growth of mixed cultures ofStreptococcusthermophilusandLactobacillusdelbrueckiisubspbulgaricus[J]. Journal of Agricultural and Food Chemistry,2011,59(13):7 045-7 053.DOI:10.1021/jf200903u.
[18] MCKAY L L, BALDWIN K A. Simultaneous loss of proteinase and lactose-utilizing enzyme activities inStreptococcuslactisand reversal by transduction[J].Applied Microbiology,1974,28(3):342-346.
[19] 白凤翎,张柏林,赵宏飞.乳酸菌蛋白代谢研究进展[J].食品科学,2010,31(19):381-384.
[20] KUNJI E R S, FANG G, JERONIMUS-STRATINGH C M, et al. Reconstruction of the proteolytic pathway for use β-casein byLactococcuslactis[J].Molecular Microbiology,1998,27(6):1 107-1 118.
Promoting growth of four lactic acid bacteria by using prepared wheat peptones
SUN Wen-jing1, XIN Xiao-ya1, GUO Wen-jie2, CUI Feng-jie1*, FANG Song-ping2, ZHU Ben-guo2, FENG Yue-qin1, XIA Tong-qian1
1(School of Food and Biological Engineering, Jiangsu University, Zhenjiang 212013, China) 2(Anhui Ruifuxiang Food Co. Ltd.,Bozhou 234800,China)
The physical and chemical features of prepared wheat peptone (WP) and its effect on lactic acid bacteria (LABs) growth were investigated and compared with other commercial peptones. Results showed that the prepared WP met the requirements of peptone quality standards and contained higher percentages of high-molecular-weight peptides (>500 u) than those in other commercial peptones. WP promoted the growth of four LABs includingLactococcuslactissubsp lactis,Lactobacillusreuteri,Lactobacilluscasei, andLactobacillusplantarumwith the highest OD600nmvalues and the microbial viable counts reached to 9.64×108CFU/mL, 2.30×109CFU/mL, 1.60×109CFU/mL, 1.14×109CFU/mL, respectively. Further studies indicated thatL.caseiseemed to preferably utilize high-molecular-weight peptides (>180 u) in WP as the nitrogen source for cell growth.
wheat peptone; nitrogen source; lactic acid bacteria; media; cell growth
10.13995/j.cnki.11-1802/ts.201703012
研究员(崔凤杰为通讯作者,E-mail:fengjiecui@163.com)。
安徽瑞福祥食品有限公司科技项目; 江苏省高等学校大学生实践创新训练计划项目(201610299126H) 、 江苏大学"青年骨干教师培养工程"青年学术带头人培育计划资助.
2016-09-22,改回日期:2016-10-20
