藏北高原草地群落的数量分类与排序
2016-12-28王景升姚帅臣王志凯冯继广
王景升, 姚帅臣, 普 穷, 王志凯, 冯继广
1 中国科学院地理科学与资源研究所生态系统网络观测与模拟重点实验室,北京 100101 2 中国人民大学环境学院,北京 100872 3 西藏大学农牧学院高原生态研究所,林芝 860000
藏北高原草地群落的数量分类与排序
王景升1, 姚帅臣2,*, 普 穷3, 王志凯2, 冯继广2
1 中国科学院地理科学与资源研究所生态系统网络观测与模拟重点实验室,北京 100101 2 中国人民大学环境学院,北京 100872 3 西藏大学农牧学院高原生态研究所,林芝 860000
采用TWINSPAN数量分类和DCA、CCA排序的方法,对藏北高原草地29个样点进行统计分析。结果显示:(1)TWINSPAN数量分类将藏北高寒草地群落划分成10种类型。(2)样点DCA排序第一轴基本反映了水分环境梯度,第二轴基本反映了热量梯度。(3)TWINSPAN分类所划分的各群落在DCA排序图上都有各自的分布范围和界限,说明DCA排序能较好的反应各优势群落与其环境资源之间的关系。(4)样点CCA排序表明,影响群落分布的首要环境因子是水分因子(年均降水量)和空间因子(经度),其次是热量因子(年均温度),CCA排序进一步阐明了群落分布决定于水分和温度等环境因子,并间接验证了TWINSPAN的分类结果。(5)物种CCA排序和TWINSPAN分类结果表明:植物群落中物种的分布格局与植物群落类型的分布格局存在一定的相似性。
藏北高原;草地;数量分类;排序;群落
青藏高原是我国乃至亚洲的江河源,向来有世界屋脊和第三极之称,是我国及东亚的重要生态安全屏障区[1]。作为青藏高原的主体部分,藏北高原是青藏高原腹地分布最为广泛、生物区系最为独特、生态功能最为重要的草地生态系统,不仅是亚洲中部高寒环境中最为典型的自然生态系统之一,在世界高寒地区亦具有代表性[2]。然而,随全球气候变化和人类活动加剧,近年来藏北高原高寒草地面临局部地区严重退化问题[3]。目前,对高寒草地的研究多集中于草地退化和沙化的治理、土壤养分流失、生物量动态以及对气候变化的响应等方面[4- 9]。而对大尺度范围内高寒草地群落的内部分类、空间分布和生态因子变化特征以及它们之间的相关性的系统研究较少,群落自身的生态特点、内在规律等尚不十分明确。数量分类和排序分析方法能够系统研究高寒草地群落的内部分类、群丛特征,并能科学地揭示植被—环境间的生态关系[10- 14]。因此,本文应用双向指示种分类法(Two Way Indicator Species Analysis,TWINSPAN)、典范对应分析(Canonical Correspondence Analysis,CCA)和去趋势对应分析(Detrended Canonical Correspondence,DCA)等方法,定量分析了环境因子对高寒草地植物群落分布和物种组成的影响。以期阐明高寒草地的地理分布格局并为合理保护利用当地草地资源提供参考。
1 研究区概况
藏北高原又称“羌塘高原”,位于西藏自治区北部,南自冈底斯—念青唐古拉山脉,北至昆仑山脉,东迄东经91°左右的内外流水系之分水岭,西止于国界,南北宽达760 km,东西长约1200 km(北纬29°53′—36°32′,东经78°41′—92°16′),面积59.5万km2,约占西藏自治区土地面积的一半。藏北高原拥有辽阔的天然草地资源,各类天然草地面积共达4800万hm2,占藏北高原土地面积的81%和西藏自治区天然草地面积的59%,为西藏主要牧区[15]。藏北高原是青藏高原主体,海拔高度在4600—5100 m之间;藏北高原寒冷干燥,大部分地区年平均气温低于0 ℃,最暖月均温不及14 ℃。年均降水量50—400 mm,且80%以上集中在6—8月,年蒸发量大于1500 mm,年平均干燥度指数1.6—20.0,年均风速高于3.0 m/s[16- 19]。天然草地类型主要包括高寒草甸、低草型高寒草原、高寒荒漠草原三大类。其中,高寒草甸的优势种为高山嵩草(Kobresiapygmaea)、紫花针茅(Stipapurpurea)等,总盖度40%—80%;高寒草原的优势种为紫花针茅、青藏苔草(Carexmoorcroftii)、矮火绒草(Leontopodiumnanum)、伊凡苔草(Carexivanovae)等,总盖度15%—50%;高寒荒漠草原的优势种为沙生针茅(Stipaglareosa)、铺散亚菊(Ajaniakhartensis)、紫花针茅等,总盖度10%—20%[20]。
2 研究方法
2.1 群落调查
采用实验生态学中典型样方调查法,在藏北羌塘高原29个样点上各设置1条100 m样线,沿样线设置5个1 m×1 m样方,样方间隔20 m,详细记录样方中各物种的高度、盖度和频度、群落平均高、总盖度等参数。样方合计145个。
2.2 气象数据
以藏北羌塘高原及其周边270个气象站长期监测数据为本底,使用Anualspline插值软件进行插值,并经过中科院地理资源所那曲生态站2009年在那曲-阿里的样带上布设的10套HOBO气象监测系统实测数据验证后,通过GIS平台系统提取获得。主要包括多年平均气温、降雨量、湿润度等。
2.3 数据处理
计算样方内各草本植物的重要值,其计算公式如下[21]:草本植物重要值=(相对盖度 + 相对高度)×100/2野外调查于2014年8月生物量高峰期完成,根据调查数据建立两个矩阵:由29个样点145个样方中74种草本的重要值组成的植被矩阵;由29个样点和5个环境参数组成的环境属性矩阵。应用Wintwins 2.3中双向指示种分析(Two Way Indicator Species Ananlysis,TWINSPAN)进行植被数量分类,应用Canoco for Windows 4.5中的典范对应分析(Canonical Correspondence Analysis,CCA)和去趋势对应分析(Detrended Canonical Correspondence,DCA)进行样方和环境参数的排序。
3 结果与分析
3.1 样方的TWINSPAN数量分类
将29个样点进行TWINSPAN分类并依据植物群落分类和命名原则,结合调查结果的生态分析和群落生境特征的指示物种或优势物种命名群落类型,本区植物群落可分为10类(如图1)。
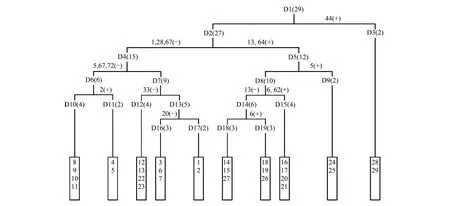
图1 样方TWINSPAN分类树状图Fig.1 Dendrogram of the TWINSPAN classification of 29 plots指示物种 1: 矮火绒草;2:矮金莲花;5:半球齿缘草;6:半卧狗娃花;13:臭棘豆;20:短穗兔耳草Lagotis brachystachya;28:高山嵩草;33:华扁穗草;44:铺散亚菊;62:西藏燥原荠;64:小垫黄芪;67:伊凡苔草;72:紫花地丁
Ⅰ紫花针茅(Stipapurpurea) + 微药羊茅(Festucanitidula) + 伊凡苔草(Carexivanovae)群落
包括样点8—11,共4个样点,分布在班戈县。该群落分布于海拔4620—4645 m,年均温-0.2—0 ℃,年降水量432—466 mm。群落平均盖度20%,优势种为紫花针茅、微药羊茅和伊凡苔草,主要伴生种有矮火绒草、高寒早熟禾(Poakoelzii)、紫花地丁(Stipapurpurea)、圆齿褶龙胆(Gentianacrenulatotruncata)等。
Ⅱ紫花针茅(Stipapurpurea) + 伊凡苔草(Carexivanovae)群落
包括样点4、5,共2个样点,分布在安多县。该群落分布于海拔4590—4630 m,年均温- 1.5℃,年降水量480—520 mm。群落平均盖度32%,优势种为紫花针茅和伊凡苔草,主要伴生种有矮金莲花(Trolliusfarreri)、矮火绒草、短叶羊茅(Festucabrachyphylla)、粗壮嵩草(Kobresiarobusta)、多枝黄芪(Astragaluspolycladus)、高山大戟(Euphorbiastracheyi)、穗三毛(Trisetumspicatum)等。
Ⅲ紫花针茅(Stipapurpurea) + 矮火绒草(Leontopodiumnanum)群落
该群落包括样点12、13、22、23,共4个样点,分布在双湖县和尼玛县。海拔4750—5000 m,年均温-1.8℃,年降水量273—384 mm。群落平均盖度13%,优势种为紫花针茅和矮火绒草,主要伴生种有二裂委陵菜(Potentillabifurca),华扁穗草(Blysmussinocompressus)和青藏苔草等。
Ⅳ高山嵩草(Kobresiapygmaea)群落,包括样点3、6、7,共3个样点,主要分布在安多县。该群落分布于海拔4685—4727 m,年均温-1.8—-1.7℃,气候较为湿润,年降水量517—528 mm。群落平均盖度达到73%,优势种为高山嵩草,主要伴生种有矮火绒草、短穗兔耳草(Lagotisbrachystachya)、高寒早熟禾、高山唐松草(Thalictrumalpinum)、西藏风毛菊(Saussureatibetica)、楔叶委陵菜(Potentillacuneata)、紫花针茅等。
Ⅴ高山嵩草(Kobresiapygmaea) + 紫花针茅(Stipapurpurea)群落
包括样点1、2,共2个样点,分布于那曲县。分布海拔4450—4460 m,年均温0.8℃,年降水量高达570 mm。该群落平均盖度51%,优势种为高山嵩草和紫花针茅,主要伴生种有二裂委陵菜、木根香青(Anaphalisxylorhiza)、西藏风毛菊、无心菜(Anaphalisxylorhiza)和楔叶委陵菜等。
Ⅵ紫花针茅(Stipapurpurea)群落
包括样点14、15、27,该群落分布于双湖县和改则县。分布海拔4600—4860 m,年均温度较低,气候寒冷,降水较少。群落平均盖度17%,优势种为紫花针茅,主要伴生种有臭棘豆(Oxytropischiliophylla)、二裂委陵菜、华扁穗草、小垫黄芪(Astragaluspuivinalis)等。
Ⅶ紫花针茅(Stipapurpurea) + 华扁穗草(Blysmussinocompressus) + 臭棘豆(Oxytropischiliophylla)群落
该群落包括样点18、19、26,分布于尼玛县和改则县。分布海拔4550—4600 m,年均温较高,降水较少。群落总盖度19%—33%,优势种为紫花针茅、华扁穗草和臭棘豆,主要伴生种有半卧狗娃花(Heteropappussemiprostratus)、多枝黄芪、二裂委陵菜和小垫黄芪等。
Ⅷ紫花针茅(Stipapurpurea) + 半卧狗娃花(Heteropappussemiprostratus) + 小垫黄芪(Astragaluspuivinalis)群落
包括样点16、17、20、21,共4个样点,分布于尼玛县。该群落分布于海拔4520—4620 m,年均温较高,年降水量310—330 mm。群落平均盖度17%,优势种为紫花针茅、半卧狗娃花和小垫黄芪,主要伴生种有西藏燥原芥(Ptilotricumwageri)、楔叶委陵菜等。
Ⅸ青藏苔草(Carexmoorcroftii) + 紫花针茅(Stipapurpurea) + 藏布红景天(Rhodiolasmithii)群落
包括样点24、25,分布于措勤县。该群落分布海拔4755—4815 m,年均温-0.8 ℃,年均降水量230—270 mm。群落平均盖度22%,优势种为青藏苔草、紫花针茅和藏布红景天,主要伴生种有半球齿缘草(Eritrichiumhemisphaericum)、垂穗披碱草(Elymusnuta)、短轴嵩草(Kobresiavidua)、裂叶独活(Heracleummillefolium)、沙蒿(Artemisiadesertorum)、无心菜、西藏风毛菊、小垫黄芪等。
Ⅹ沙生针茅(Stipaglareosa) + 紫花针茅(Stipapurpurea) + 铺散亚菊(Ajaniakhartensis)群落
该群落包括样点28、29,分布于革吉县。群落分布海拔4430—4490 m,年均温较高,降水较少,气候非常干旱。群落平均盖度较低,只有11%,物种多为耐干旱植物,优势种为沙生针茅、紫花针茅和铺散亚菊,主要伴生种有西藏燥原芥、固沙草(Orinusthoroldii)等。
3.2 样点DCA排序分析
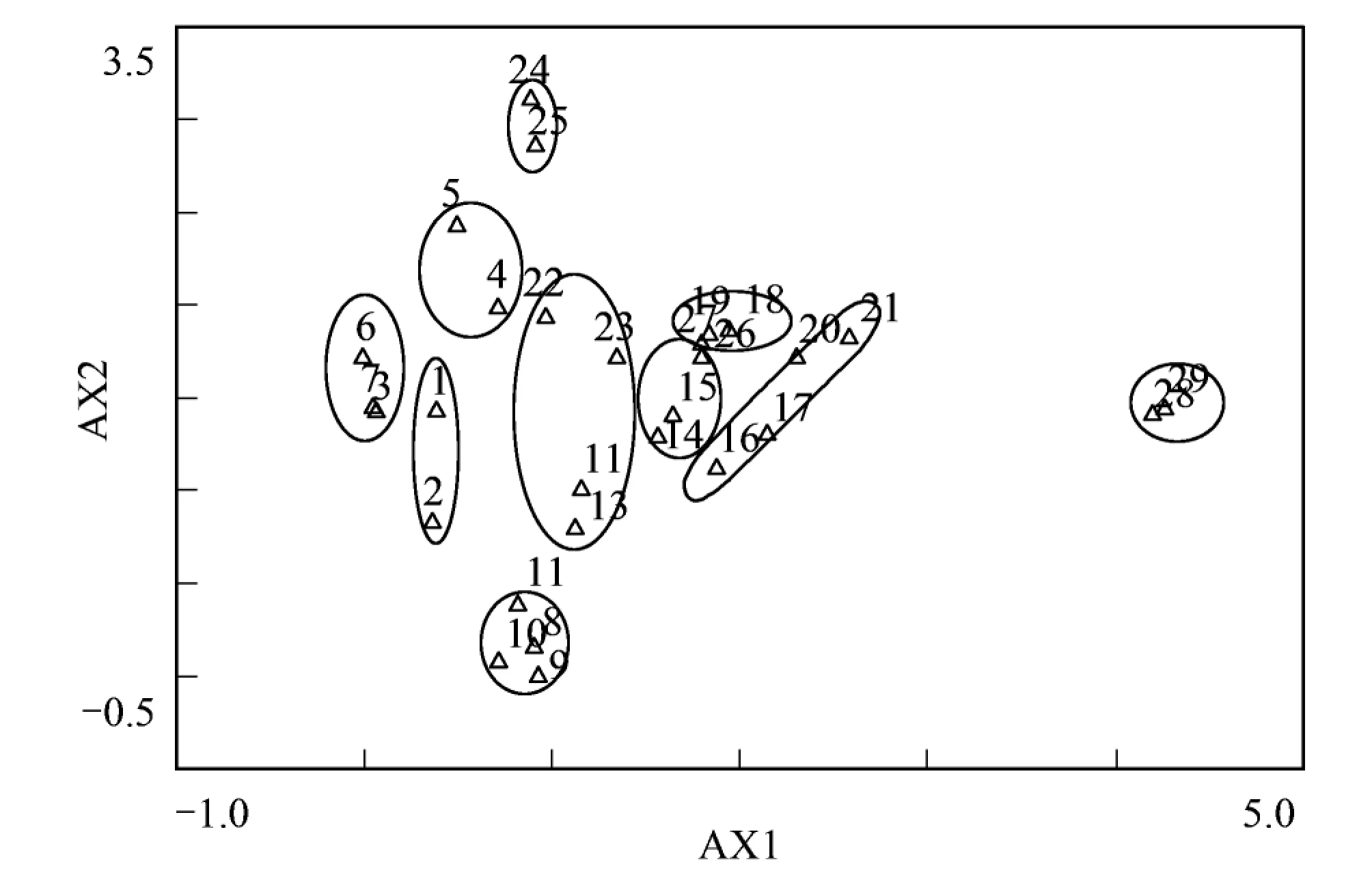
图2 样点DCA二维排序图Fig.2 Two-dimensional DCA ordination diagram of 29 plots
采用DCA对研究区29个样点进行排序分析,结果如图2,得到前4个排序轴的特征值分别为0.681、0.459、0.196、0.119。第一二轴特征值较大,说明这两个排序轴包含的生态信息量多,具有更重要的生态意义。从二维排序图可以看出,TWINSPAN分类结果所得的群落类型在DCA二维排序图上均有自己的分布范围和界限,说明DCA排序较好的反应了各样方之间和样方与环境之间的关系。从排序轴看,第一轴基本反映了水分环境梯度,从左到右,环境条件由较为湿润过渡到干旱。第二轴反映了温度梯度的变化,即第二轴从下往上,年均温逐渐降低。样点的DCA排序图综合反映了植物群落和生境的空间梯度,与TWINSPAN的分类结果完全匹配。
3.3 样点CCA排序
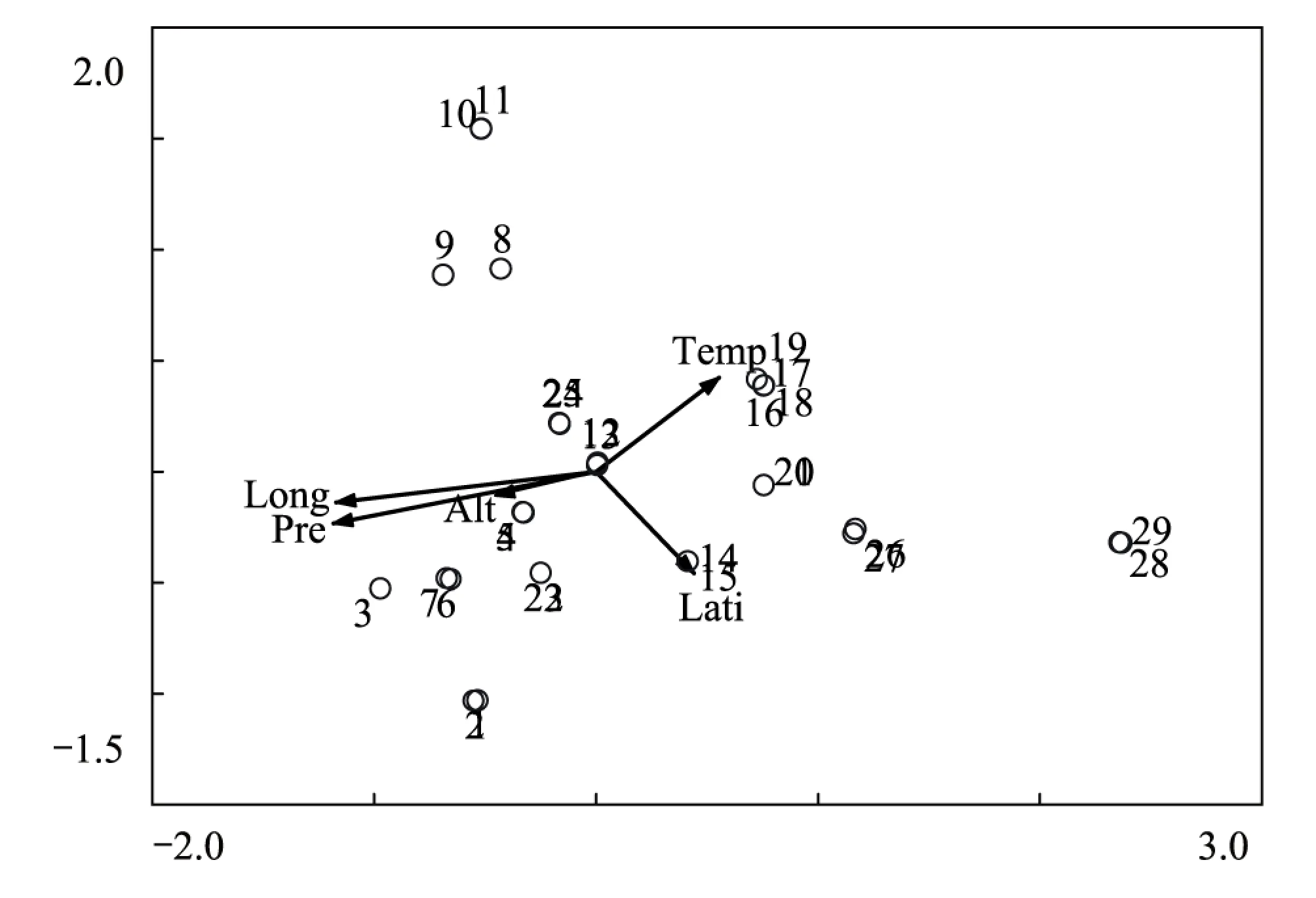
图3 样点CCA排序图Fig.3 CCA ordination diagram of 29 plots
典范对应分析(Canonical Correspondence Analysis,CCA),是一种基于单峰模型的排序方法,样方排序与对象排序对应分析,而且在排序过程中结合多个环境因子,因此可以把样方、对象与环境因子的排序结果表示在同一排序图上[10]。为进一步探讨群落分布与环境因子之间的关系,对29个样点和5个环境因子进行CCA排序,结果如图3。排序图中箭头表示环境因子,箭头连线的长度代表着相应环境因子与研究对象分析相关程度的大小,越长代表其对所研究对象的分布影响越大;箭头连线与排序轴夹角余弦值代表其与排序轴的相关性大小。沿排序图第一轴,随着年降水量的逐渐减少,群落从排序图最左侧生境相对湿润的高山嵩草群落和高山嵩草 + 紫花针茅群落过渡到中间较为干旱的紫花针茅群落、紫花针茅 + 半卧狗娃花 + 小垫黄芪群落、青藏苔草 + 紫花针茅 + 藏布红景天群落,再到最右侧干旱的沙生针茅 + 紫花针茅 + 铺散亚菊群落。沿排序轴第二轴从上向下,随着纬度增加,温度降低,群落由紫花针茅 + 微药羊茅 + 伊凡苔草群落过渡到高山嵩草群落。从排序结果可以看出,影响群落分布的首要环境因子是水分因子(年均降水量)和空间因子(经度),其次是热量因子(年均温度)。即是说,首先,水分条件的差异决定着研究区的植被被分为三种类型:相对湿润的高寒草甸、较为干旱的高寒草原和干旱高寒荒漠草原,其次,在水分条件相差不大的情况下,由于温度的不同,3种植被类型又被分为不同的群落。此外,排序图中可以看出TWINSPAN划分的十类群落在CCA排序中基本都有各自的分布范围和界限,这说明TWINSPAN的分类结果和样方CCA排序是一致的。
3.4 物种的CCA排序
采用CCA对74个物种进行数量分析,结果如图4所示。由图可以看出,物种的分布格局与图3样点DCA中样点的分布格局相似。物种CCA第一轴反映了各物种分布生境的水分条件,沙生针茅、固沙草、西藏燥原芥等耐旱植物分布在排序图最右侧较为干旱的区域,高山嵩草、无茎黄鹌菜、短穗兔耳草、西藏风毛菊等分布生境较为湿润的植物则分布在排序图左侧。半卧狗娃花、斑唇马先蒿、冰川棘豆、梭罗草等与温度相关性比较大,均分布在相对温暖的区域,而紫花针茅、伊凡苔草、二裂委陵菜、华扁穗草、矮火绒草等分布在中央,说明这几种植物生态位适应性较强,分布较为广泛。结合分类结果和样点DCA排序图,可以看到植物群落中物种的分布格局与植物群落类型的分布格局有很大的相关性。
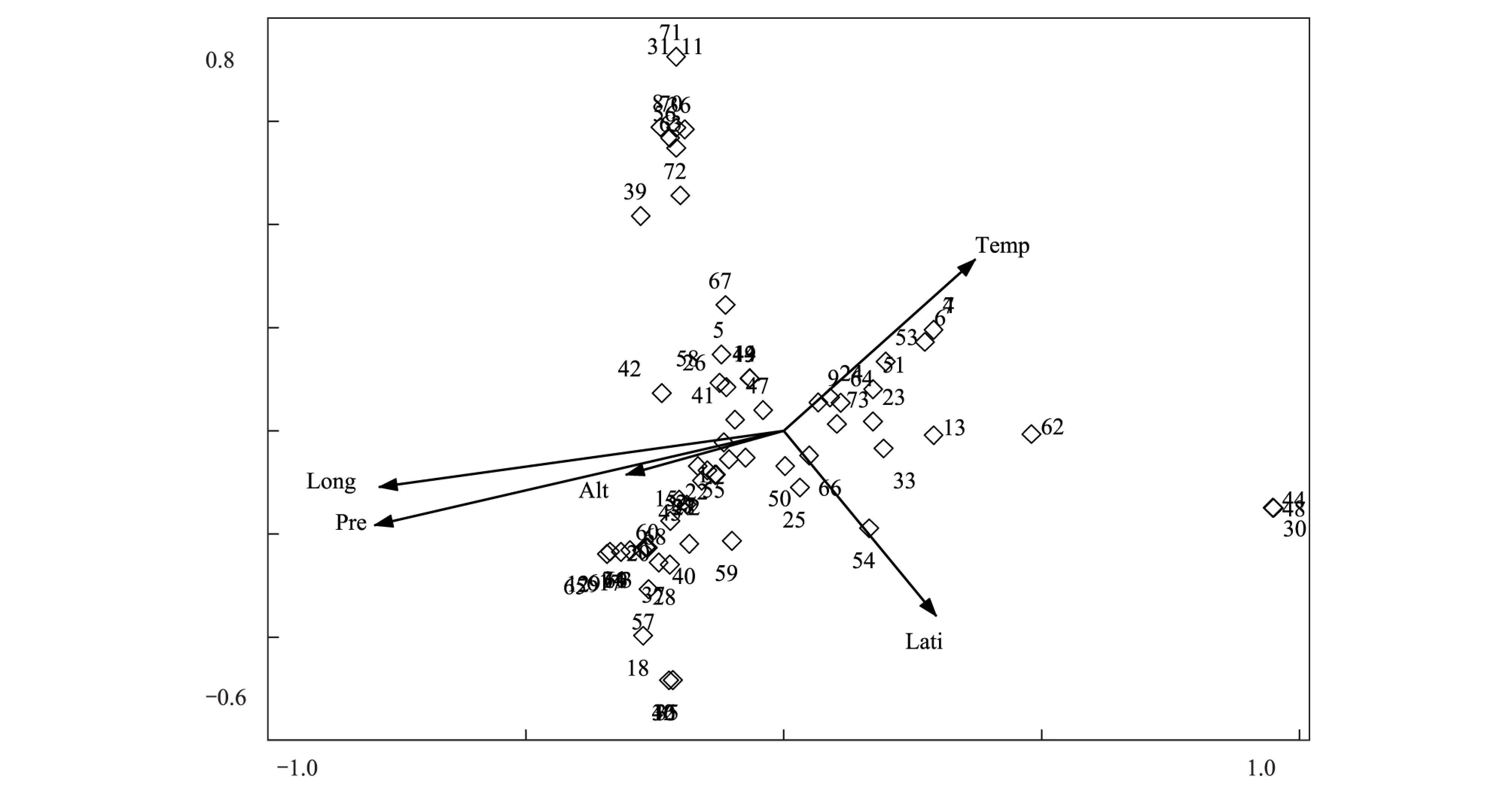
图4 物种CCA排序图Fig.4 CCA ordination diagram of 74 species3: 白蒿 Leontopodium dedekensii; 4: 斑唇马先蒿 Pedicularis longiflora; 7: 冰川棘豆 Oxytropis glacialis; 8: 藏菠萝花 Incarvillea compacta; 9: 藏布红景天; 10: 藏豆 Stracheya tibetica; 11: 藏沙蒿 Artemisia wellbyi; 12: 策勒蒲公英 Taraxacum qirae; 14: 垂穗披碱草; 15: 粗壮嵩草; 16: 单子麻黄 Ephedra monosperma; 17: 垫状点地梅 Androsace tapete; 18: 钉柱委陵菜 Potentilla saundersiana; 19: 独一味 Lamiophlomis rotate; 21: 短叶羊茅; 22: 短轴嵩草;23: 多枝黄芪; 24: 二花棘豆 Oxytropis biflora; 25: 二裂委陵菜;26: 高寒早熟禾; 27: 高山大戟; 29: 高山唐松草;30: 固沙草; 31: 禾叶点地梅 Androsace graminifolia; 32: 黑包风毛菊 Saussurea melanotrica; 34: 金露梅 Potentilla fruticosa; 35: 蕨麻委陵菜 Potentilla anserine; 36: 赖草 Leymus secalinus; 37: 兰石草 Lancea tibetica; 38: 蓝翠雀花 Delphinium caeruleum; 39: 狼毒 Euphorbia fischeriana; 40: 藜 Chenopodium album; 41: 裂叶独活; 42: 木根香青; 43: 螃蟹甲 Phlomis younghusbandii; 45: 青藏苔草; 46: 弱小火绒草; 47: 沙蒿; 48: 沙生针茅; 49: 少花棘豆 Oxytropis pauciflora; 50: 四数獐牙菜 Swertia tetraptera; 51: 碎米荠 Cardamine hirsute; 52: 穗三毛 Trisetum spicatum; 53: 梭罗草 Roegneria thoroldiana; 54: 团垫黄芪 Astragalus arnoldii; 55: 微孔草 Microula sikkimensis; 56: 微药羊茅; 57: 无茎黄鹌菜 Youngia simulatrix;58: 无心菜; 59: 西伯利亚蓼 Polygonum sibiricum; 60: 西藏风毛菊; 61: 西藏棱子芹 Pleurospermum hookeri; 63: 纤杆蒿 Artemisia demissa; 65: 小叶棘豆 Oxytropis microphylla; 66: 楔叶委陵菜; 68 羽叶钉柱委陵菜 Potentilla saundersiana; 69: 羽毛委陵菜 Potentilla plumose; 70: 圆齿褶龙胆; 71: 胀果棘豆 Oxytropis stracheyana; 73: 紫花针茅; 74: 紫菀 Aster tataricus
4 结论与讨论
TWINSPAN数量分类将藏北草原29个样点划分为10种群落类型,即紫花针茅 + 微药羊茅 + 伊凡苔草群落;紫花针茅 + 伊凡苔草群落;紫花针茅 + 矮火绒草群落;高山嵩草群落;高山嵩草 + 紫花针茅群落;紫花针茅群落;紫花针茅 + 华扁穗草 + 臭棘豆群落;紫花针茅 + 半卧狗娃花 + 小垫黄芪群落;青藏苔草 + 紫花针茅 + 藏布红景天群落;沙生针茅 + 紫花针茅 + 铺散亚菊群落。采用DCA对29个样点进行排序分析,各群落在排序图上都有各自的分布范围和界限,可见DCA排序能较好的反应各样方之间和样方与环境之间的关系,同时TWINSPAN的分类结果也在排序图上得到较好的印证。样方DCA排序第一轴基本反映了水分环境梯度,第二轴基本反映了温度梯度。
样点CCA排序结果表明,影响群落分布的首要环境因子是水分因子(年均降水量)和空间因子(经度),其次是热量因子(年均温度),CCA排序进一步阐明了群落分布于环境之间的关系,并再次验证了TWINSPAN的分类结果。物种CCA的排序结果显示,植物群落中物种的分布格局与植物群落类型的分布格局存在一定的相似性。
对于DCA排序轴的生态解释,不同学者的研究结果有所不同,有些学者认为,DCA第一轴反映水分环境梯度,第二轴反映热量环境梯度[21- 24]。部分学者则认为DCA第一轴反映(海拔)热量环境梯度,第二轴反映水分环境梯度[25- 28]。还有学者认为,DCA排序第一轴反映了群落海拔高度的梯度变化,第二轴反映了群落坡度的梯度变化或者坡向的梯度变化[29-30]。Burke等人的研究则表明,在一定程度上,DCA第一轴反映土壤性质和养分梯度,而在小环境中,水分条件和干扰程度决定着植被的空间分布[31]。由此,我们可以看出,DCA排序轴的环境解释在不同的研究地点和尺度,有可能得出不同的研究结论。在降雨相对丰富地区,可能热量因素更大程度上影响了植被的组成和空间分布,而在西部黄土高原等干旱地区,水分条件可能成为影响植被空间分布的主导因素,而在干旱荒漠地区,水分条件相差不大的地区,土壤养分含量也有可能成为影响植被分布的关键因子[24]。藏北高原地处大陆腹地,平均海拔4600m以上,年均温相差不大,年均降水量较少,且由于自身高原地貌的影响,降水随着经度的减小而逐渐减少。在较低的降水水平下,降水的减少会降低土壤中的有效水分、改变土壤的理化性质、影响植物对养分的吸收、运输和利用,进而影响群落中各类型植物的比例和重要性。物种的CCA排序结果显示:在环境较为湿润的高寒草甸中,群落中湿生植物占绝对优势,而在干旱的荒漠草原,旱生植物则占有绝对优势。这充分说明水分条件在很大程度上影响着物种的分布以及群落的结构组成,因此可以认为水分条件是影响藏北草原植被空间分布的主导因素。
综上,本文采用TWINSPAN分类结合DCA和CCA排序的研究方法,对藏北草地的数量分析取得了良好的分类结果,并初步揭示了高寒草地植物群落及环境因子的分布格局。可为藏北草原的保护和利用,以及相关的植被群落研究提供理论依据。
致谢:本文实验样地以中国科学院地理科学与资源研究所藏北高寒草甸生态系统研究站(那曲站)为依托,样地调查及制图过程中得到了站内工作人员和武建双博士、田源和曲芸莹硕士的帮助,石培礼研究员和付刚老师对文章写作给予帮助,特此致谢。
[1] 孙鸿烈, 郑度, 姚檀栋, 张镱锂. 青藏高原国家生态安全屏障保护与建设. 地理学报, 2012, 67(1): 3- 12.
[2] 张荣祖, 郑度, 杨勤业. 西藏自然地理. 北京: 科学出版社, 1982.
[3] 王景升, 张宪洲, 赵玉萍, 秦泗国, 武建双. 羌塘高原高寒草地生态系统生产力动态. 应用生态学报, 2010, 21(6): 1400- 1404.
[4] 武建双, 李晓佳, 沈振西, 张宪洲, 石培礼, 余成群, 王景升, 周宇庭. 藏北高寒草地样带物种多样性沿降水梯度的分布格局. 草业学报, 2012, 21(3): 17- 25.
[5] 张生楹, 陈谷, 陈建纲, 张德罡. 藏北高原高寒草地土壤有效养分含量研究. 草原与草坪, 2012, 32(3): 6- 10.
[6] 蔡晓布, 张永青, 邵伟. 藏北高寒草原草地退化及其驱动力分析. 土壤, 2007, 39(6): 855- 858.
[7] 赵景学, 陈晓鹏, 曲广鹏, 多吉顿珠, 尚占环. 藏北高寒植被地上生物量与土壤环境因子的关系. 中国草地学报, 2011, 33(1): 59- 64.
[8] 杨秀海, 卓嘎, 罗布. 藏北高原气候变化与植被生长状况. 草业科学, 2011, 28(4): 626- 630.
[9] 付刚, 周宇庭, 沈振西, 张宪洲, 武建双. 藏北高原高寒草甸地上生物量与气候因子的关系. 中国草地学报, 2011, 33(4): 31- 36.
[10] 张金屯. 数量生态学. 北京: 科学出版社, 2004.
[11] Oldeland J, Dorigo W, Lieckfeld L, Lucieer A, Jürgens N. Combining vegetation indices, constrained ordination and fuzzy classification for mapping semi-natural vegetation units from hyperspectral imagery. Remote Sensing of Environment, 2010, 114(6): 1155- 1166.
[12] Ermakov N, Makhatkov I. Classification and ordination of north boreal light-coniferous forests of the West Siberian Plain. Plant Biosystems, 2011, 145(S1): 199- 207.
[13] Dale E E Jr, Ware S, Waitman B. Ordination and classification of bottomland forests in the lower Mississippi alluvial plain. Castanea, 2007, 72(2): 105- 115.
[14] Ahmad S S, Quratulann. Vegetation classification in ayubia national park, pakistan using ordination methods. Pakistan Journal of Botany, 2011, 43(5): 2315- 2321.
[15] 胡自治. 青藏高原的草业发展与生态环境. 北京: 中国藏学出版社, 2000: 1- 5, 19- 22, 100- 124.
[16] 毛飞, 卢志光, 郑凌云, 张佳华. 近40年那曲地区日照时数和风速变化特征. 气象, 2006, 32(9): 77- 83.
[17] 毛飞, 卢志光, 张佳华, 郑凌云. 近40年那曲地区气候特征分析. 高原气象, 2007, 26(4): 708- 715.
[18] 王景升, 张宪洲, 赵玉萍, 沈振西, 石培礼, 余成群. 藏北羌塘高原气候变化的时空格局. 资源科学, 2008, 30(12): 1852- 1859.
[19] 毛飞, 卢志光, 张佳华, 侯英雨. 近20年藏北地区AVHRR NDVI与气候因子的关系. 生态学报, 2007, 27(8): 3198- 3205.
[20] 李明森. 藏北高原草地资源合理利用. 自然资源学报, 2000, 15(4): 335- 339.
[21] 马克平, 刘玉明. 生物群落多样性的测度方法: I α多样性的测度方法(下). 生物多样性, 1994, 2(4): 231- 239.
[22] 张峰, 张金屯, 张峰. 历山自然保护区猪尾沟森林群落植被格局及环境解释. 生态学报, 2003, 23(3): 421- 427.
[23] 张桂莲, 张金屯, 程林美. 山西南部山地白羊草群落的数量分类和排序. 草业学报, 2003, 12(3): 63- 69.
[24] 董林水, 张旭东, 周金星, 宋爱云. 晋西黄土丘陵与土石山区交错地带灌木种的数量分类与排序. 生态学报, 2007, 27(7): 3072- 3080.
[25] 岳鹏鹏, 卢学峰, 叶润蓉, 周玉碧, 赵建忠, 彭敏. 长江源区紫花针茅草原群落数量分类与排序. 广西植物, 2014, 34(5): 635- 641.
[26] 史作民, 刘世荣, 程瑞梅, 蒋有绪. 河南宝天曼植物群落数量分类与排序. 林业科学, 2000, 36(6): 20- 27.
[27] 刘秋锋, 康慕谊, 刘全儒. 中条山混沟地区森林乔木种的数量分类与环境解释. 植物生态学报, 2006, 30(3): 383- 391.
[28] 岳鹏鹏, 孙菁, 卢学峰, 叶润蓉, 周玉碧, 杨仕兵, 彭敏. 青海高原紫花针茅草原群落数量分类与排序. 西北植物学报, 2014, 34(5): 1047- 1054.
[29] 何惠琴, 李绍才, 孙海龙, 刘世昌, 熊文兰. 锦屏水电站植被数量分类与排序. 生态学报, 2008, 28(8): 3706- 3712.
[30] 陈云, 王海亮, 韩军旺, 韦博良, 贾宏汝, 叶永忠, 袁志良. 小秦岭森林群落数量分类、排序及多样性垂直格局. 生态学报, 2014, 34(8): 2068- 2075.
[31] Burke A. Classification and ordination of plant communities of the Naukluft Mountains, Namibia. Journal of Vegetation Science, 2001, 12(1): 53- 60.
Quantitative classification and ordination of grassland communities on the Northern Tibetan Plateau
WANG Jingsheng1, YAO Shuaichen2,*, PU Qiong3, WANG Zhikai2, FENG Jiguang2
1KeyLaboratoryofEcosystemNetworkObservationandModeling,InstituteofGeographicSciencesandNaturalResourcesResearch,ChineseAcademyofSciences,Beijing100101,china2SchoolofEnvironmentandNaturalResources,RenminUniversityofChina,Beijing100872,China3TibetInstituteofPlateauEcology,TibetAgriculturalandAnimalHusbandryCollege,Linzhi860000,China
The relationship between vegetation and the environment is one of the fundamental questions in understanding plant species composition and community distribution along an environmental gradient. Understanding vegetation-environment correlations may help predict possible shifts in plant communities in response to climate and land use changes. There are 48 million hm2of natural grasslands on the Northern Tibetan Plateau, accounting for 59% of the total grassland area in the Tibet Autonomous Region. However, these alpine grasslands are threatened by global climate change and intense human activities in recent decades, and are now widely degraded. Quantitative classification and ordination are important methods in examining the internal classification of plant communities, characteristics of their associations, and in revealing the ecological relationships between vegetation and the environment. TWINSPAN classification and DCA,CCA ordination were used to conduct a classification and ordination of the plant communities of 29 plots in the northern Tibetan grassland. Our results indicate that: (1) Northern Tibetan alpine meadows are classified into 10 associations:Stipapurpurea+Festucanitidula+Carexivanovae;Stipapurpurea+Carexivanovae;Stipapurpurea+Leontopodiumnanum;Kobresiapygmaea;Kobresiapygmaea+Stipapurpurea;Stipapurpurea;Stipapurpurea+Blysmussinocompressus+Oxytropis;Stipapurpurea+Heteropappussemiprostratus+Astragaluspuivinalis;Carexmoorcroftii+Stipapurpurea+Rhodiolasmithii; andStipaglareosa+Stipapurpurea+Ajaniakhartensis; (2) The first axis of DCA basically reflected a moisture gradient, and the second axis indicated a thermal gradient; (3) Each community divided by TWINSPAN classification had its specific distribution and boundaries on the DCA ordination diagram, which indicated that DCA ordination can explain the relationships between the plots and environment; (4) CCA plots indicated that the primary environmental factors controlling the distribution of communities were rainfall (average annual rainfall) along longitude, followed by temperature (average annual temperature); CCA sorting further elaborated the relationship between plant community distribution and the environment, and indirectly supported the results of the TWINSPAN classification; (5) Quantitative analysis of 74 species by CCA indicated that the distribution pattern of species and plots in DCA was similar. The first axis of species CCA reflected water conditions of species distribution. Taken together, we conclude that the distribution patterns of plant species communities closely mirror each other. In general, the present study provides an ecological interpretation of the distribution of plant species communities along an environmental gradient on the Northern Tibetan Plateau.
Northern Tibetan Plateau; grassland; quantitative classification; ordination; community
国家科技支撑课题资助项目(2011BAC09B03);科技部973课题资助项目(2013CB956300);国家自然科学基金资助项目(41271067)
2015- 04- 24;
日期:2016- 03- 03
10.5846/stxb201504240848
*通讯作者Corresponding author.E-mail: yaoshuaichen@yeah.net
王景升, 姚帅臣, 普穷, 王志凯, 冯继广.藏北高原草地群落的数量分类与排序.生态学报,2016,36(21):6889- 6896.
Wang J S, Yao S C, Pu Q, Wang Z K, Feng J G.Quantitative classification and ordination of grassland communities on the Northern Tibetan Plateau.Acta Ecologica Sinica,2016,36(21):6889- 6896.
