非线性回归方法建立亚热带常绿阔叶树种地上生物量相对生长方程
2016-12-21侯燕南吴惠俐
侯燕南 ,吴惠俐
(1. 中南林业科技大学,湖南 长沙 410004;2. 湖南省林业厅 造林处,湖南 长沙 410004)
非线性回归方法建立亚热带常绿阔叶树种地上生物量相对生长方程
侯燕南1,2,吴惠俐1
(1. 中南林业科技大学,湖南 长沙 410004;2. 湖南省林业厅 造林处,湖南 长沙 410004)
我国亚热带地区森林类型多样、组成结构复杂、碳吸存能力高,是我国亚热带地区重要的碳库。然而,关于亚热带常绿阔叶林生物量和碳储量估算方法的研究报道较少。本研究选择亚热带常绿阔叶林中3个优势树种青冈Cyclobalanopsis glauca、豹皮樟Litsea rotundifolia和木荷Schima superba为研究对象,构建以胸径(DBH)为自变量的树种各器官生物量相对生长方程。结果表明:3个常绿阔叶树种各器官生物量估算相对生长方程具有显著相关性(P<0.000 1),拟合效果好,决定系数均大于0.914 7;青冈树种生物量相对生长方程拟合效果优于木荷和豹皮樟;树干和地上部分生物量相对生长方程拟合效果优于树枝和树叶。本研究结果为亚热带常绿阔叶林生物量和碳储量的估算提供准确有效的估算方法。
亚热带常绿阔叶林;青冈;豹皮樟;木荷;相对生长方程;胸径
我国亚热带地区气候为温暖湿润季风气候[1-2]。此气候区天然林中树种丰富,在其演替后期常绿阔叶树种占主导地位。由于长期的人类活动干扰和土地利用,大部分的常绿阔叶林已经转变为人工林或次生林[3]。如需通过可持续经营管理措施将人工林或次生林演替为常绿阔叶林,必须明确常绿阔叶林林分结构是如何影响森林生长动态和生物地球化学循环的,而树种和林分生物量数据是重要的基础数据[4],有效的、可接受的精确程度上的生物量估算方法是必不可少的[5]。
相对生长方程是常用的生物量估算方法之一。相对生长方程常以简单的测树变量进行构建,如胸径(DBH)、树高(H)[6-7]等。目前,量化某个区域、某个生物地理分布区、某个国家甚至全球尺度的森林生物量和碳储量日渐需要,这促进了热带[8]、温带[9]和寒带[10]树种或森林的相对生长方程的构建与检验,也促进了全球相对生长方程数据库的发展与完善[11]。在中国,关于温带[9]和亚热带[12]森林生物量相对生长方程构建已有报道,但多针对于具有重要经济价值的针叶树种,如杉木Cunninghamia lanceolata和马尾松Pinus massoniana等。对于常绿阔叶树种很少有合适可用的相对生长方程,构建常绿阔叶树种生物量相对生长方程为亚热带森林生物量和碳储量估算提供重要的估算方程。
本研究以亚热带常绿阔叶林中3个优势树种(青冈Cyclobalanopsis glauca、木荷Schima superba和豹皮樟Litsea rotundifolia)为研究对象,主要研究目的为:(1)构建树干、树枝、树叶和地上生物量相对生长方程;(2)检验树种相对生长方程的精确度。
1 材料与方法
1.1 研究地概况
本研究地在湖南省靖州县排牙山国有林场进行。排牙山国有林场纬度为 26°24′~ 26°35′N,经度为 109°27′~ 109°38′E,此处为雪峰山系低山丘陵地貌,海拔为230~107 5 m。气候属于中亚热带季风湿润气候,年均温为16.7 ℃,其最冷月(1月)的月均温为5.7 ℃,最热月(7月)的月均温为26.8 ℃,无霜期,290 d左右。年均降水量为125 0 mm,降雨量集中在4月到8月,年均蒸发量为884 mm。母岩为紫色砂页岩,海拔低于600 m区域的土壤为红壤,海拔高于600 m区域的土壤为山地黄壤。植被类型主要为杉木人工林和由本土树种组成的次生林。
1.2 研究方法
1.2.1 生物量测定
生物量的测定于2014年10月之前完成。样地调查后,根据树叶形态及其物候特征,选择常绿阔叶林中3个优势树种(青冈Cyclobalanopsis glauca、木荷Schima superba和豹皮樟Litsea rotundifolia)进行研究。测定这3个树种的胸径(DBH)、树高(H)、地上生物量(kg)以及树所在坐标位置,并进行记录。根据每个树种DBH的最大值和最小值,设定5个等级,于每个等级随机选取两棵树,共10棵样本树。各树种尺寸见表1。

表1 生物量相对生长方程构建的样本量和样本树尺寸Table 1 Sample size and dimensions of trees harvested to determine allometric biomass equations
样本树地上部分生物量测定方法采用收获法。用电锯将样本树伐倒,进行树种鉴定并记录各测树指标变量值(如1.3 m处胸径,树高,树基部直径,1/2处直径,低于最低活枝处直径,冠幅)。将树干从1.3 m处、2 m间隔处截断,然后直至树冠。将树枝从树干的各个部分分离,并记录各树枝所在的位置(地面高)。树干部分和树枝部分(带叶)原地称量。所有给定树种树干部分重量之和为树干的总鲜重。基于树枝的直径,于茎每个部位树枝(带叶)随机取3到5个随机样本,用于计算树枝和叶片生物量比。将每个树枝样本的叶片分离,并分开称量树叶和树枝的鲜质量。树枝和树叶的比值可用来估测树叶和树枝的总鲜质量。叶片和树枝的次级样本以及树干各部分所截圆盘均被装入布袋,然后带回实验室进行水分含量的测定。所有样品均在80 ℃烘箱中烘干至恒质量。树地上部分鲜重通过各自水分含量转换为干质量。树地上部分生物量就是树干、树枝和树叶生物量的总和。
1.2.2 相对生长方程构建
通过上述生物量测定方法获得了树干、树枝、树叶和地上部分生物量,利用散点图来检验各器官生物量与DBH之间的关系,结果如图1所示。通过散点图分析,结果表明常绿阔叶树树干、树枝、树叶和地上部分生物量与DBH之间呈明显的非线性关系。
考虑到上述分析,决定利用非线性回归模型构建各器官生物量估算的相对生长方程[13],方程如下:

式(1)中,Wi为树器官i的生物量(kg/tree),包括树干、树枝、树叶和地上生物量;DBH为自变量;a和b为回归系数;ε为正态分布N(0,σ2)假定的随机误差。
本研究所有数据分析均在R3.2.0软件[14]进行。
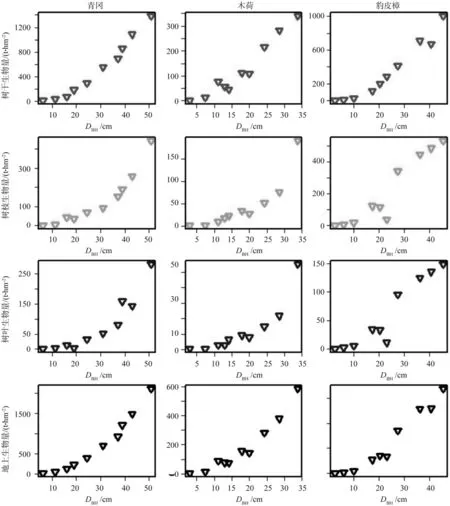
图1 常绿阔叶树(青冈,木荷,豹皮樟)树干、树枝、树叶和地上部分生物量(kg/tree)与DBH(cm)散点(n=120)Fig. 1 Observed stem, branch, leaf and aboveground biomass (kg/tree) plotted against DBH (cm) for evergreen broadleaved species (Cyclobalanopsis glauca, Schima superba and Litsea rotundifolia) (n=120)
2 结果与分析
2.1 青冈地上生物量相对生长方程
青冈树干、树枝、树叶和地上部分生物量方程均显著相关(p<0.001)。各器官生物量相对生长方程RMSE分别为:树干46.396 1,树枝15.983 8,树叶18.446 2,地上部分46.497 4。各器官生物量相对生长方程R2值分别为:树干0.991 7,树枝0.988 2,树叶0.973 3,地上部分0.999 5。通过分析,青冈树干和地上生物量方程拟合效果比树枝和树叶拟合效果好。结果如表2所示。
2.2 木荷地上生物量相对生长方程
木荷树干、树枝、树叶和地上部分生物量方程均具有显著相关性(p<0.001)(见表2)。树干、树枝、树叶和地上生物量相对生长方程RMSE分别为:21.440 9,12.648 5,3.420 6和21.514 3。树干、树枝、树叶和地上生物量相对生长方程R2值分别为:0.983 2,0.929 1,0.961 8和0.987 9。通过分析,木荷各器官生物量相对生长方程拟合效果整体比青冈稍差,而与青冈相似的是树干和地上部分生物量相对生长方程拟合效果优于树枝和树叶。结果如表2所示。

表2 3个常绿树种地上部分各器官i生物量(Wi,kg)与胸径(DBH,cm)的相对生长方程 [ln(Wi)=a+b×ln(DBH)]†Table 2 Allometric equations [ln(Wi)=a+b×ln(DBH)]relating aboveground biomass component i (Wi, kg) to diameter at breast height (DBH, cm) for the three tree species
2.3 豹皮樟地上生物量相对生长方程
豹皮樟各器官生物量,包括树干、树枝、树叶和地上部分,生物量相对生长方程均著相关(p<0.001)。树干生物量相对生长方程RMSE为56.034 7,树枝为66.815 1,树叶为18.709 5,地上部分为109.170 6。树干生物量相对生长方程R2值为0.977 0,树枝和树叶为0.914 7,地上部分为0.972 1。通过分析,豹皮樟各器官生物量相对生长方程拟合效果整体差于青冈和木荷,而与青冈和木荷相似的是树干和地上部分生物量相对生长方程拟合效果优于树枝和树叶。结果如表2所示。
3 结论与讨论
本研究构建了亚热带常绿阔叶林中3个优势树种(青冈、木荷和豹皮樟)各器官生物量估算相对生长方程。3个常绿阔叶树种拟合的所有相对生长方程,拟合效果非常好,其拟合决定系数R2值均大于0.914 7,其中青冈树干和地上生物量相对生长方程拟合效果最好,决定系数R2值分别为0.991 7和0.999 5。尽管3个常绿树种各器官生物量相对生长方程拟合效果很好,但相对来说,以DBH为估算方程自变量对树干和地上生物量估算精度更高。本研究获得的树种各器官生物量估算的相对生长方程能在高精确度水平上估算亚热带常绿阔叶林的森林生物量。
对以DBH为自变量的所有相对生长方程进行分析,发现DBH是地上各器官生物量估算的良好预测变量(P<0.000 5)(表2),此研究结果与前人研究报道的结果一致[8-15]。当增加其它测树因子(如树高、冠副等)作为共同变量时,并未明显增加生物量估算精度[16-17],又由于在林分调查过程中,野外树高测量工作量大,且存在一定的误差。所以,在实际工作中,建议采用DBH作为生物量估算的预测变量。
我国亚热带森林物种丰富,结构复杂,生物多样性高,导致森林生物量和碳储量的估算难度大且精度低。因此,构建亚热带森林中更多优势树种的高精度生物量估算的相对生长方程,并对其进行精度验证对我国亚热带森林生物量和碳储量估算是非常重要的,这需要更进一步的研究。
[1]Zhang J, Ge Y, Chang J,et al.Carbon storage by ecological service forests in Zhejiang Province, subtropical China[J]. Forest Ecology and Management, 2007, 245:64-75.
[2]Yu GR, Chen Z, Piao SL,et al. High carbon dioxide uptake by subtropical forest ecosystems in the East Asian monsoon region[J]. proceedings of the national academy of sciences of the united states of america, 2014, 111(13):4910-4915.
[3]Zhang H, Guan D, Song M. Biomass and carbon storage of Eucalyptus and Acacia plantation in the Pearl River Delta, South China[J]. Forest Ecology and Management, 2012, 277:90-97.
[4]Ares A, Fownes JH. Comparisons between generalized and specific tree biomass functions as applied to tropical ash(Fraxinus uhdei)[J]. New forests, 2000, 20(3):277-286.
[5]Bi H, Murphy S, Volkova L,et al.Additive biomass equations based on complete weighing of sample trees for open eucalypt forest in south-eastern Australia[J]. Forest Ecology and Management, 2015, 349:106-121.
[6]Williams RJ, Zerihun A, Montagu KD,et al.Allometry for estimating aboveground tree biomass in tropical and subtropical eucalypt woodlands: towards general predictive equations[J].Agriculture ecosystems & environment, 2005, 53(7):607-619.
[7]Pilli R, Anfodillo T, Carrer M. Towards a functional and simplified allometry for estimating forest biomass[J]. Forest Ecology and Management, 2006, 237(1):583-593.
[8]Basuki TM, Laake PEv, Skidmore AK,et al. Allometric equations for estimating the above-ground biomass in tropical lowland Dipterocarp forests[J]. Forest Ecology and Management 2009, 257:1684-1694.
[9]Wang CK. Biomass allometric equations for 10 co-occurring tree species in Chinese temperate forests[J]. Forest Ecology and Management, 2006, 222(1):9-16.
[10]Berner LT, Alexander HD, Loranty MM,et al. Biomass allometry for alder, dwarf birch, and willow in boreal forest and tundra ecosystems of far northeastern Siberia and north-central Alaska[J]. Forest Ecology and Management, 2015, 337:110-118.
[11]Henry M, Bombelli A, Trotta C,et al.GlobAllomeTree:international platform for tree allometric equations to support volume, biomass and carbon assessment[J]. Iforestbiogeosciences and forestry, 2013, (6): 326-330.
[12]Xiang WH, Liu SH, Deng XW,et al. General allometric equations and biomass allocation of Pinus massoniana trees on regional scale in southern China[J]. Canada Ecological Research,2011, 26(4):697-711.
[13]Wagner, R.G, Ter-Mikaelian, M.T. Comparison of biomass component equations for four species of northern coniferous tree seedlings. Annals of Forest Science, 1999, 56:193-199.
[14]R Core Team (2015). R: A language and environment for statistical computing. R Foundation for Statistical Computing,Vienna, Austria. URL http://www.R-project.org/.
[15]Zianis D, Mencuccini M. On simplifying allometric analyses of forest biomass[J]. Forest Ecology and Management, 2004,187(2):317-332.
[16]zhang C, Peng DL, Huang GS,et al.Developing Aboveground Biomass Equations Both Compatible with Tree Volume Equations and Additive Systems for Single-Trees in Poplar Plantations in Jiangsu Province, China[J]. forests, 2016, 7(2):32-47.
[17]张绘芳, 高亚琪, 李 霞, 等. 新疆西伯利亚云杉生物量模型研究[J]. 中南林业科技大学学报, 2015, 35(11): 115-120.
Using nonlinear regression method to develop allometric equations for aboveground biomass estimate of three evergreen broadleaved tree species in subtropical China
HOU Yan-nan1,2, WU Hui-li1
(1. Central South University of Forestry and Technology, Changsha 410004, Hunan, China;2. Silviculture Administration Section, Forestry Department of Hunan Province, Changsha 410004, Hunan, China)
Subtropical China has diverse forests which contain a diversity of tree species and exhibit a high carbon (C) sequestration capacity. However, at present, the equations of the forest biomass and C stocks estimation in subtropical China have been rare reported.Hence, we selected evergreen broadleaf forest of subtropical China to develop species-speci fi c allometric equations for three dominant species (Cyclobalanopsis glauca,Schima superba,Litsea rotundifolia) for the function group of evergreen broadleaved species. Speciesspeci fi c equations using DBH alone as the predictor variable fi tted the data well for each tree component. Species-speci fi c equations for three tree species performed well with high correlation coef fi cients (R2>0.9147,P<0.0001), the fi tted effects of species-speci fi c allometric equations forC. glaucawere corresponding lightly higher than that forS. superbaandL. rotundifolia, the fi tted effects of species-speci fi c allometric equations for stem and aboveground were higher than that for branch and leaf. Our study may provide precise and valid equations for forest biomass and C stocks estimation in evergreen broadleaved forest of subtropical China.
subtropical evergreen broadleaved forest;Cyclobalanopsis glauca;Litsea rotundifolia;Schima superba; allometric equations; diameter at breast height
S718.55+4.2
A
1673-923X(2016)12-0098-04
10.14067/j.cnki.1673-923x.2016.12.017
http: //qks.csuft.edu.cn
2016-05-06
国家林业公益性行业专项项目(201304317)
侯燕南,高级工程师,博士研究生;E-mail:yannanhou@hotmail.com
侯燕南,吴惠俐. 非线性回归方法建立亚热带常绿阔叶树种地上生物量相对生长方程[J].中南林业科技大学学报,2016,36(12): 98-101, 107.
[本文编校:文凤鸣]
