miRNA211在人皮肤黑素瘤的表达及功能研究
2016-11-06徐妍董达科华海康朱小红
徐妍 董达科 华海康 朱小红
214002 江苏,无锡市第二人民医院肿瘤科(徐妍),皮肤科(董达科、华海康、朱小红)
miRNA211在人皮肤黑素瘤的表达及功能研究
徐妍 董达科 华海康 朱小红
214002 江苏,无锡市第二人民医院肿瘤科(徐妍),皮肤科(董达科、华海康、朱小红)
目的探讨miRNA211(miR⁃211)在恶性黑素瘤进展过程中的表达以及靶分子MMP⁃16与miR⁃211表达的相关关系。方法实验分阴性对照组、空载体对照组、miR⁃211过表达组。TaqMan荧光定量PCR检测miR⁃211在黑素细胞、黑素瘤细胞的表达,以及在痣、黑素瘤组织中的表达。SRB试验和流式细胞仪分别检测细胞增殖和细胞周期分布。甲基纤维素克隆形成试验和Transwell迁移试验研究细胞克隆形成和细胞迁移。用集落大小衡量克隆形成力,而同一迁移距离下的相对细胞数用来衡量细胞迁移能力。TaqMan荧光定量PCR检测比较miR⁃211表达增加前后MMP⁃16 mRNA表达的变化。结果miR⁃211在黑素瘤细胞G361、C32和A375表达量分别是0.09± 0.02,0.000 52±0.000 20,0.000 03±0.000 01(F=10410,P<0.01)。miR⁃211在黑素瘤标本的表达量(0.17±0.03)显著低于痣的表达量(0.87±0.08),t=9.118,P<0.01。在体外,miR⁃211表达上调的黑素瘤细胞miR⁃211过表达组与阴性对照组及空载体对照组比较,细胞增殖与细胞周期分布,差异无统计学意义。miR⁃211过表达组、空载体对照组的克隆形成力分别是0.49±0.05、0.85±0.09,两组比较,差异有统计学意义(t=2.19,P<0.05)。miR⁃211过表达组、空载体对照组的克隆形成力分别是0.49±0.06、0.82±0.09,两组比较,差异有统计学意义(t=3.15,P<0.05)。而空载体对照组与阴性对照组比较,差异无统计学意义。转染miR⁃211 mimics后24 h,在miR⁃211过表达组和空载体对照组的MMP⁃16 mRNA水平比较分别是0.33±0.02和0.91±0.03,两组比较,t=11.30,P<0.01;48 h分别是0.52±0.01和0.96±0.02,两组比较,t=5.02,P<0.05;72 h分别是0.71±0.01和0.97±0.03,两组比较,t=3.85,P<0.05。空载体对照组与阴性对照组在3个时间点的比较,差异均无统计学意义。结论miR⁃211在恶性黑素瘤的细胞水平和组织水平低表达。miR⁃211抑制黑素瘤细胞的锚着非依赖性生长及细胞的迁移。miR⁃211表达上调后,MMP⁃16 mRNA表达下调,可能是miR⁃211下游的靶分子从而影响黑素瘤的侵袭转移。
黑色素瘤;微小RNAs;肿瘤转移;基质金属蛋白酶16;miRNA211
恶性黑素瘤是一种皮肤黏膜的恶性肿瘤。在世界范围内,皮肤恶性黑素瘤发病率的年增长率3%~5%[1]。在我国每年新发病例约2万[2]。2007年,国外两个微小RNA(miRNA)芯片研究发现黑素瘤特异表达的 miRNA(miR)[3⁃4]。后续的基因芯片的实验结果发现,miR⁃200家族、miR⁃211、miR⁃141、miR⁃182、miR⁃183等在皮肤黑素瘤组织、细胞水平特异性表达[5⁃6]。王震英等[7⁃8]发现,let⁃7a在黑素瘤中表达下降,且let⁃7a可抑制A375细胞增殖并促进A375细胞凋亡,同时下调caspase3蛋白表达。实验证明,基质金属蛋白酶(MMP)1、2、3、9、14、15、16在体内外均高表达,并表现出侵袭的表型[9]。通过Diana Tool(http://diana.imis.athena ⁃innovation.gr/DianaTools/index.php)筛选 miRNA211(miR⁃211)的靶基因时,我们发现在357个预测靶分子MMP⁃16排在17位,尚无实验相关数据。因此,我们初步探讨了miR⁃211新的靶分子MMP⁃16与miR⁃211表达的相关关系。
资料与方法
一、临床资料
收集2010—2012年经组织病理学确诊18例痣中,男10例,女8例;年龄23~71岁,中位年龄50.5岁。2002—2012年经病理组织学确诊41例黑素瘤石蜡切片。41例黑素瘤患者中男23例,女18例。年龄31~74岁,中位年龄53.3岁。
二、主要材料
原代黑素细胞HER1及恶性皮肤黑素瘤细胞A375、C32、G361、MelM培养基及补充物(美国ATCC公司)。MEM、DMEM培养基、胎牛血清、OPTI⁃MEM无血清培养基和胰蛋白酶细胞培养所需试剂(美国Gibco公司)。细胞总RNA(Trizol,美国Invitrogen公司)和组织总RNA(Recover AllTM,美国Ambion公司)抽提试剂。转染试剂(siPORTNeoFX Transfection Agent)、miRNATaqMANqPCR 试 剂、MMP⁃16 Taq Man Assay、miR⁃211类似物、miR⁃Scramble空载体对照试剂(美国Applied Biosystems公司)。Matrigel™基底膜基质(美国BD Biosciences公司)。甲基纤维素(美国RD System公司)。
三、方法
1.细胞培养、转染及转染效率检测:将人黑素细胞系接种于MelM培养基,人黑素瘤细胞系接种于DMEM培养基,随后置于37℃、5%CO2的恒温箱中培养,隔日换液或传代培养。选择A375细胞进行下游功能实验。按说明书要求以等量OPTI⁃MEM无血清培养基对miR⁃211类似物或空载体对照进行稀释。后将转染试剂与上述稀释的小RNA按要求混匀、静置后加入待用培养板。最后将对数生长期的A375细胞接种于含有上述小RNA和转染试剂混液的培养板中。接种浓度参考12孔板1×105/孔,6孔板2×105/孔浓度。培养基每天更换。转染效率的检测使用FAM⁃labelled Pre⁃miR的阴性对照,通过流式细胞仪检测荧光吸收率来确定转染效率。
2.各细胞及组织的RNA抽提及cDNA合成:按照Trizol和RecoverAllTM说明书步骤抽提各株细胞总RNA和组织中的总RNA,取RNA于紫外分光度计下检测260 nm和280 nm的吸光度(A值),A260/280为1.8~2.1时,符合纯度要求。按照miRNA反转录试剂盒说明书合成cDNA。
3.实时定量PCR检测miR⁃211和MMP⁃16的表达:miR⁃92在黑素瘤及黑素细胞表达稳定性高于其他内参照,因此选择miR⁃92作为miRNAqPCR的内参照,β肌动蛋白作为mRNA表达检测的内参照。将合成的cDNA稀释10倍,再按qPCR说明书配制反应体系,通用引物由试剂盒提供,每个样本设置3个复孔取均值。PCR反应步骤:95℃10 min,95 ℃ 15 s,60 ℃ 1 min,40个循环。miR⁃211序列:UUCCCUUUGUCAUCCUUCGCCU。记录各实验孔的 Ct值,目的基因的相对表达量 F=2⁃ΔΔCt,其中ΔCT=CT目的基因⁃CT内参基因,ΔΔCT= ΔCT实验组-ΔCT对照组,2-ΔΔCt表示目的基因相对于对照组的表达倍数。
4.细胞增殖、细胞周期检测:实验设阴性对照组、空载体对照组、miR⁃211过表达组。每组重复16孔,重复3次实验。以1 000~1 500个细胞/孔浓度接种于96孔板,每组有16个复孔。实验检测24、48、72、96 h。磺酰罗丹明B(SRB)实验步骤如下:每孔加入50 μl 25%(V/V)TCA,4℃孵育不少于1 h。用水清洗96孔板10 min,自然干。加入50 μl的0.057%(W/V)SRB溶液,室温孵育30 min。用1%(V/V)150 μl乙酸洗板4次,自然干。最后每孔加入150 μl 10 mmol/L Tris溶液(pH 10.5)振荡孵育1 h,在A510 nm读板获取数据。
碘化丙锭检测细胞周期:上述3个组每组2管重复3次取均值。用胰酶消化转染后的A375细胞。随后用PBS冲洗、离心细胞各2次。新细胞团重悬于柠檬酸缓冲液并保存在-20℃。在FACS检测前将细胞柠檬酸缓冲液中先后加入一定量的A溶液混合2 min、B溶液10 min及C溶液10 min后,此时的细胞混合液即可行FACS细胞周期检测。该实验分别检测了转染后24、48、72 h的细胞周期分布情况。
5.克隆形成实验:克隆的大小用来衡量克隆形成力。上述3个组每组2孔重复3次取均值。分别检测转染后第1、3、6、9、12天的克隆大小。具体步骤:将1.8%(W/V)琼脂糖胶溶液加热至沸,液态的琼脂糖胶与2×MEM培养基按1∶1比例混合,在6孔板中每孔加入2 ml的上述混合液,室温放置使其凝固。将甲基纤维素溶液与2×MEM按说明书要求的比例混合。随后将转染后16~18 h的细胞与甲基纤维素培养基按比例混合。以每孔2 ml甲基纤维素细胞混合液加入到准备好的琼脂糖胶6孔板,每孔细胞量为5 × 103~ 104个。第1、3、6、9、12天在显微镜下观察摄片。每实验孔计算30个克隆大小的均值。Image J软件计算克隆大小,Gel Doc计算每孔的克隆数量。
6.Transwell细胞迁移实验:上述3个组每组2孔重复3次取均值。先将Matrigel在冰上慢慢溶解,PBS和Matrigel按1∶1比例混匀。在插有Transwell的24孔板中每孔加入100 μl上述混合液。24孔板在37℃孵育箱孵育30 min使得Matrigel在Transwell底部凝固。将转染后16~18 h的细胞按(1~4)×105/ml浓度与培养基混合制成细胞混悬液。当Matrigel凝固后将Transwell倒置。再将每孔100 μl的细胞混悬液加到Transwell的底部过滤膜上,然后将倒置的板放入细胞培养箱2~4 h。此时细胞粘附在细胞板,将细胞板正置。随后用无血清培养基浸洗每个Transwell孔3次。最后在Transwell每孔里加入100 μl含血清的常规培养基。上述含有Transwell的24孔板放入孵育箱培养5 d。每日更换常规培养基。观察摄片前对细胞进行固定和染色。共聚焦显微镜成像系统按每10 μm间隔进行扫描摄片,Image J软件处理后续的图像数据。使用相对细胞数对3组进行细胞迁移力的比较。相对细胞数是指将进行细胞计数的40 μm处细胞数/0 μm处细胞数,这样可以排除因初始加细胞人为造成的细胞数量不同的误差。
7.统计分析:用GraphPad Prism Version 5.0软件进行数据分析。计量资料以±s表示。计量资料的的两两比较采用Studentt检验,多组比较采用单因素方差分析。以P<0.05为差异有统计学意义。
结 果
一、miR⁃211在黑素细胞和黑素瘤细胞的表达
miR⁃211在3株黑素瘤细胞G361、C32、A375的表达量相比黑素瘤细胞HER1分别是0.09±0.02、0.000 52±0.000 2、0.000 03±0.000 01(F=10 410,P<0.01)。miR⁃211在3株黑素瘤细胞的表达量相比黑素细胞显著下降,尤其是在A375细胞中,miR⁃211的表达量仅是黑素细胞的万分之一。
二、miR⁃211在痣与黑素瘤组织标本的表达
miR⁃211在痣的表达量0.87±0.08(n=18),在黑素瘤的表达量0.17±0.03(n=41),两者比较差异有统计学意义(t=9.118,P<0.01)。同样的,miR⁃211在黑素瘤组织中的表达量比痣组织的表达量减少了80%。
三、miR⁃211类似物转染A375细胞的转染效率及转染后miR⁃211的表达
用含有荧光蛋白的空载体小RNA转染人黑素瘤A375细胞24、48、72 h后,用FACS检测转染效率,转染效率超过80%。体外转染miR⁃211类似物至A375细胞后通过荧光qPCR方法测得miR⁃211表达上调百万倍。
四、miR⁃211过表达后细胞增殖、细胞周期分布
miR⁃211过表达组与阴性对照组、空载体对照组在细胞增殖和细胞周期的比较,差异均无统计学意义,见表1,2。在本实验中没有发现miR⁃211的上调能够影响A375细胞的增殖及凋亡。
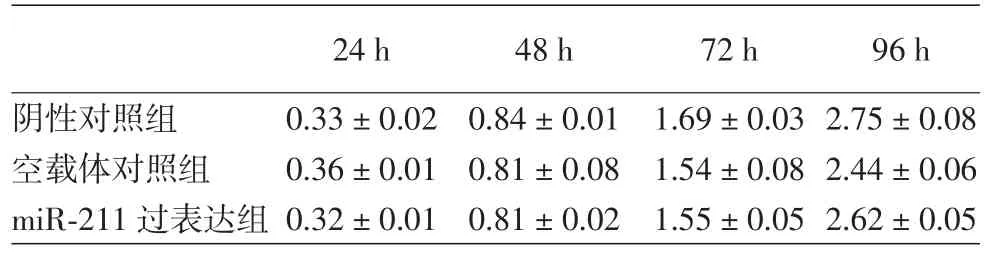
表1 3个组A375瘤细胞细胞增殖的情况
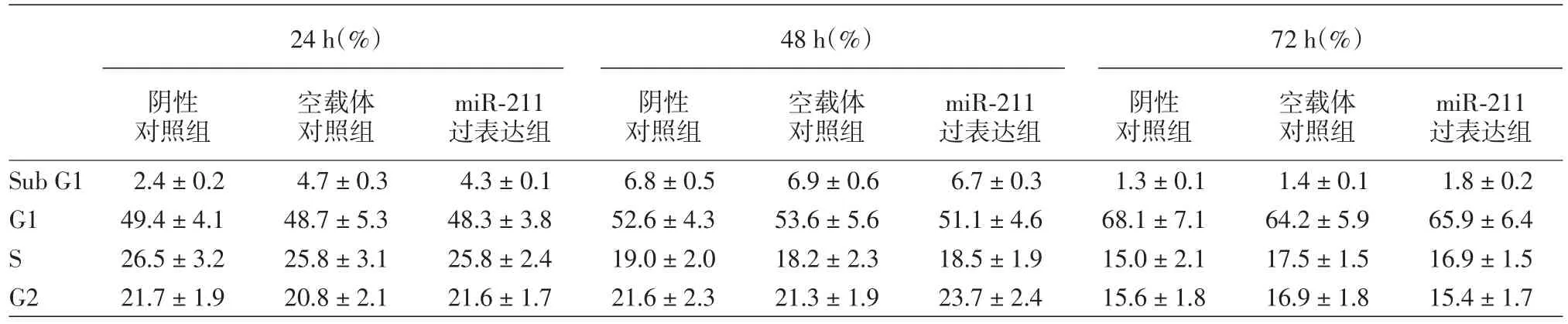
表2 3个组A375瘤细胞细胞周期的情况
五、miR⁃211过表达后细胞克隆形成力及侵袭力
1.克隆形成试验:第6天,miR⁃211过表达组、空载体对照组的克隆形成力分别是0.49±0.05、0.85±0.09,两组比较,差异有统计学意义(t=2.19,P<0.05,图1A~1C);阴性对照组和空载体对照组的差异无统计学意义(t=1.83,P>0.05)。在A375细胞中,miR⁃211的过表达使得肿瘤细胞克隆较miR⁃211未过表达瘤细胞的克隆缩小了43%。第12天,可见miR⁃211过表达组克隆个数(305)明显少于阴性对照组(770)、空载体对照组的克隆数量(547)(图1D~1F)。

图1 甲基纤维素实验比较转染后第6天阴性对照组、空载体对照组和miR⁃211过表达组A375细胞的致瘤能力 1A~1C:每组随机选取30个克隆,并计算出每个克隆大小的±s(×100);1D~1F:GelDoc拍摄了1组实验在观察至转染后第12天,3个组细胞克隆的数量大体观
2.Transwell细胞迁移试验:实验发现,转染后第5天miR⁃211过表达组、空载体对照组的细胞迁移力分别是0.49±0.06、0.82±0.09,两组比较差异有统计学意义(t=3.153,P<0.05)。阴性对照组和空载体对照组的差异无统计学意义(t=1.046,P>0.05,图2)。在A375细胞中,miR⁃211的过表达使得细胞迁移的距离显著小于miR⁃211未过表达的瘤细胞,肿瘤细胞迁移距离缩短了41%。
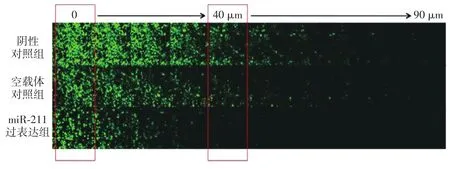
图2 Transwell细胞迁移实验比较阴性对照组、空载体对照组和miR⁃211过表达组3个组A375细胞的侵袭转移能力。转染后第5天,3个组从40 μm的这层开始miR⁃211过表达组的相对细胞数量与阴性对照组和空载体对照组比较,差异有统计学意义(相对细胞数是将进行细胞计数的40 μm处细胞数/0 μm处细胞数)。每组均随机选6个视野中细胞数的±s。共聚焦显微镜拍摄0~90 μm共100 μm长的距离。每层厚度10 μm
六、miR⁃211过表达后MMP⁃16 mRNA的表达
24 h后,空载体对照组和miR⁃211过表达组相对表达量分别是0.91±0.03、0.33±0.02,两组比较,t=11.3,P<0.01。48 h后的相对表达量分别是0.96±0.02、0.52±0.01,两组比较,t=5.02,P<0.05。72 h后的相对表达量分别是0.97±0.03,0.71±0.01,两组比较,t=3.85,P< 0.05。阴性对照组和空载体对照组的MMP⁃16 mRNA表达差异无统计学意义。miR⁃211过表达组的MMP⁃16的表达量在转染后3 d内均显著减少。在A375瘤细胞,MMP⁃16在miR⁃211过表达后24 h表达量减少了近60%。上述结果提示miR⁃211与MMP⁃16表达呈负相关。
讨 论
研究发现,miRNA除了能够精确地确定单一肿瘤的类型[10],还能够发挥重要的抑癌基因和(或)原癌基因的作用。因此,miRNA不仅可以成为新型的生物标志物,进一步研究特异表达的miRNA可以为肿瘤的临床治疗提供新的方向。
通过已发表的miRNA基因芯片的研究结果,我们挑选了一些特异表达的miRNA进行表达的检测及功能的研究。我们发现miR⁃211在恶性黑素瘤细胞和黑素瘤组织中均显著低表达。而有些miRNA,比如miR⁃205、23b它们在黑素瘤组织和黑素瘤细胞中表达情况并不一致[5]。在功能实验中,我们进一步发现增加miR⁃211在黑素瘤中的表达显著降低了黑素瘤细胞的克隆形成力和肿瘤侵袭力,但并未影响肿瘤细胞的增殖及细胞周期分布,这与Boyle实验组的结果一致[11]。而Mazar等[12]实验发现miR⁃211同时影响肿瘤细胞增殖和侵袭转移。我们的实验结果虽然没有发现miR⁃211的上调能够影响A375细胞的增殖及凋亡,但细胞克隆的形成除了表现在细胞增殖外,还表现在细胞锚着非依赖性生长能力方面。我们实验的阳性结果说明miR⁃211在抑制肿瘤发生发展中的作用主要表现在抑制细胞锚着非依赖性生长的特性上。
国外研究发现,miR⁃211的直接靶基因,如IGF2R、TGFBR2、NFAT5和NUAK1与细胞周期、免疫调节及侵袭转移息息相关[13⁃14]。在寻找miR⁃211新的靶基因的过程中我们选择了MMP⁃16。除了在筛选miR⁃211的预测靶基因MMP⁃16排位较高以外还结合MMP⁃16本身在肿瘤的作用。MMP⁃16与在多种侵袭性强的恶性肿瘤高表达的MMP⁃14不同,它仅在部分侵袭性高的结节型恶性肿瘤如胶质瘤、肝癌、胃癌及黑素瘤高表达[15]。研究发现,MMP⁃16在黑素瘤作为一种基质依赖的调节因子影响黑素瘤的侵袭转移。它的表达与快速纤维蛋白侵袭有关,从而限制胶原的侵袭,且必须依赖MMP⁃14来促进结节型细胞的增殖[15]。MMP⁃16还被认为是黑素瘤的不良预后因子。本研究初次探讨了MMP⁃16与miR⁃211在黑素瘤表达的相关关系,发现MMP⁃16在miR⁃211的表达呈负相关。miR⁃211与MMP⁃16的相关关系是如何进一步影响黑素瘤的发生发展尚待进一步研究。
[1]Lens M.Current clinical overview of cutaneous melanoma[J].Br J Nurs,2008,17(5):300 ⁃305.DOI:10.12968/bjon.2008.17.5.28825.
[2]CSCO黑素瘤专家委员会.中国黑素瘤诊治指南(2011版)[J].临床肿瘤学杂志,2012,17(2):159⁃171.DOI:10.3969/j.issn.1009⁃0460.2012.02.015.Melanoma Committee of CSCO.Guidelines for the diagnosis and treatment of melanoma(2011)[J].Chin Clin Oncol,2012,17(2):159⁃171.DOI:10.3969/j.issn.1009⁃0460.2012.02.015.
[3]Gaur A,Jewell DA,Liang Y,et al.Characterization of microRNA expression levels and their biological correlates in human cancer cell lines[J].Cancer Res,2007,67(6):2456 ⁃2468.DOI:10.1158/0008⁃5472.CAN⁃06⁃2698.
[4]Blower PE,Verducci JS,Lin S,et al.MicroRNA expression profiles for the NCI⁃60 cancer cell panel[J].Mol Cancer Ther,2007,6(5):1483⁃1491.DOI:10.1158/1535⁃7163.MCT⁃07⁃0009.
[5]Philippidou D,Schmitt M,Moser D,et al.Signatures of microRNAs and selected microRNA target genes in human melanoma[J].Cancer Res,2010,70(10):4163⁃4173.DOI:10.1158/0008⁃5472.CAN⁃09⁃4512.
[6]Mueller DW,Rehli M,Bosserhoff AK.miRNA expression profiling in melanocytes and melanoma cell lines reveals miRNAs associated with formation and progression of malignant melanoma[J].J Invest Dermatol,2009,129(7):1740⁃1751.DOI:10.1038/jid.2008.452.
[7]王震英,张捷,王焱,等.微小RNA⁃let⁃7a对黑素瘤细胞系A375细胞凋亡的影响[J].中华皮肤科杂志,2010,43(8):575⁃578.DOI:10.3760/cma.j.issn.0412⁃4030.2010.08.020.Wang ZY,Zhang J,Wang Y,et al.Effect of microRNA⁃let⁃7a on apoptosis in melanoma cell line A375[J].Chin J Dermatol,2010,43(8):575⁃578.DOI:10.3760/cma.j.issn.0412⁃4030.2010.08.020.
[8]王震英,王焱,薛燕宁,等.let⁃7a微小RNA在黑素瘤中的表达及其对A375细胞增殖的影响[J].中华皮肤科杂志,2010,43(5):358⁃361.DOI:10.3760/cma.j.issn.0412⁃4030.2010.05.021.Wang ZY,Wang Y,Xue YN,et al.Expression of microRNA let⁃7a in melanoma and its effect on the proliferation of A375 human melanoma cell line[J].Chin J Dermatol,2010,43(5):358⁃361.DOI:10.3760/cma.j.issn.0412⁃4030.2010.05.021.
[9]Ntayi C,Hornebeck W,Bernard P.Involvement of matrix metallo⁃proteinases(MMPs) in cutaneous melanoma progression[J].PatholBiol(Paris),2004,52(3):154⁃159.DOI:10.1016/j.patbio.2004.02.002.
[10]Zhang B,Pan X,Cobb GP,et al.microRNAs as oncogenes and tumor suppressors[J].Dev Biol,2007,302(1):1 ⁃12.DOI:10.1016/j.ydbio.2006.08.028.
[11]Boyle GM,Woods SL,Bonazzi VF,et al.Melanoma cell invasi⁃veness is regulated by miR⁃211 suppression of the BRN2 transcription factor[J].Pigment Cell Melanoma Res,2011,24(3):525⁃537.DOI:10.1111/j.1755⁃148X.2011.00849.x.
[12]Mazar J,DeYoung K,Khaitan D,et al.The regulation of miRNA⁃211 expression and its role in melanoma cell invasiveness[J].PLoS One,2010,5(11):e13779.DOI:10.1371/journal.pone.0013779.
[13]Levy C,Khaled M,Iliopoulos D,et al.Intronic miR⁃211 assumes the tumor suppressive function of its host gene in melanoma[J].Mol Cell,2010,40(5):841 ⁃849.DOI:10.1016/j.molcel.2010.11.020.
[14]BellRE,KhaledM,NetanelyD,etal.Transcription factor/microRNA axis blocks melanoma invasion program by miR⁃211 targeting NUAK1[J].J Invest Dermatol,2014,134(2):441 ⁃451.DOI:10.1038/jid.2013.340.
[15]Tatti O,Arjama M,Ranki A,et al.Membrane⁃type⁃3 matrix metalloproteinase(MT3⁃MMP)functions as a matrix composition⁃dependent effector of melanoma cell invasion[J].PLoS One,2011,6(12):e28325.DOI:10.1371/journal.pone.0028325.
Expression and function of miRNA211 in human cutaneous melanoma
Xu Yan,Dong Dake,Hua Haikang,Zhu Xiaohong
Department of Oncology,Wuxi No.2 People′s Hospital,Wuxi 214002,China(Xu Y);Department of Dermatology,Wuxi No.2 People′s Hospital,Wuxi 214002,China(Dong DK,Hua HK,Zhu XH)
ObjectiveTo determine the expression of miRNA211(miR⁃211)in the development of malignant melanoma,and to investigate the correlation between miR⁃211 and its target molecule,matrix metalloproteinase 16(MMP⁃16).MethodsCultured A375 melanoma cells were divided into 3 groups:miR⁃211 overexpression group and mock⁃vehicle group transfected with miR⁃211 mimics and empty vehicle respectively,and negative control group receiving no treatment.TaqMan fluorescence⁃based quantitative PCR was performed to determine the expression of miR⁃211 in HER1 primary melanocytes,A375,C32 and G361malignant melanoma cell lines,as well as in nevus tissues(n=18)and melanoma tissues(n=41),and to evaluate changes of MMP⁃16 mRNA expression in A375 cells before and after the overexpression of miR⁃211.Sulforhodamine B(SRB)assay and flow cytometry were conducted to evaluate cellular proliferative activity and determine cell cycle distribution respectively,and methylcellulose assay and Transwell assay to evaluate colony formation and cell migration abilities respectively.The size of selected colonies was used to represent colony formation ability,while the ratio of the number of migrating cells to that of non⁃migrating cells to represent cell migration ability.ResultsThere were significant differences in the expression level of miR⁃211 among the G361,C32 and A375 cells(0.09±0.02vs.0.000 52±0.000 20vs.0.000 03±0.000 01,F=10 410,P<0.01).The expression of miR⁃211 was significantly decreased in melanoma tissues compared with nevus tissues(0.17±0.03vs.0.87±0.08,t=9.118,P<0.01).No significant differences were observed in cellular proliferative activity or cell cycle distribution among the miR⁃211 overexpression group,mock⁃vehicle group and negative control group.Compared with the mock⁃vehicle group,the miR⁃211 overexpression group showed significantly suppressed colony formation(0.49±0.05vs.0.85±0.09,t=2.19,P<0.05)and cell migration(0.49±0.06vs.0.82±0.09,t=3.15,P<0.05)abilities,while no significant difference was observed between the mock⁃vehicle group and negative control group.Additionally,the mRNA expression of MMP⁃16 significantly decreased in the miR⁃211 overexpression group compared with the mock⁃vehicle group after transfection(24 hours:0.33±0.02vs.0.91±0.03,t=11.30,P<0.01;48 hours:0.52±0.01vs.0.96±0.02,t=5.02,P<0.05;72 hours:0.71±0.01vs.0.97±0.03,t=3.85,P<0.05),with no significant difference between the mock⁃vehicle group and negative control group at the above time points.Conclusions miR⁃211 was lowly expressed in both malignant melanoma cells and tissues,and it could inhibit both anchorage⁃independent growth and migration of melanoma cells.After up⁃regulation of miR⁃211 expression,the mRNA expression of MMP⁃16 decreased in A375 cells,suggesting that MMP⁃16 may be a downstream target of miR⁃211,and can influence melanoma metastasis.
Melanoma;MicroRNAs;Neoplasm metastasis;Matrix metalloproteinase 16;miRNA211
Zhu Xiaohong,Email:zxh6801@126.com
2015⁃11⁃30)
(本文编辑:吴晓初)
朱小红,Email:zxh6801@126.com
10.3760/cma.j.issn.0412⁃4030.2016.09.006
南京医科大学重点项目(2013NJMU193)
Fund program:Major Project of Nanjing Medical University(2013NJMU193)
