果寡糖对急性氨氮应激下团头鲂非特异性免疫指标的影响
2016-09-23张春暖任洪涛张纪亮吴秋珏高小蝉刘文斌
张春暖,任洪涛,张纪亮,吴秋珏,高小蝉,刘文斌
(1.河南科技大学动物科技学院,河南洛阳 471003;2.南京农业大学动物科技学院,南京 210095)
果寡糖对急性氨氮应激下团头鲂非特异性免疫指标的影响
张春暖1,任洪涛1,张纪亮1,吴秋珏1,高小蝉1,刘文斌2
(1.河南科技大学动物科技学院,河南洛阳471003;2.南京农业大学动物科技学院,南京210095)
为研究氨氮应激条件下果寡糖对团头鲂(Megalobramaamblycephala)非特异免疫指标的影响,试验选择360尾初重为13.5 g的团头鲂随机分为3组,每组4个重复,每缸30尾鱼,对照组投喂基础日粮,试验组分别投喂果寡糖(FOS)添加量为0.4%和0.8%的日粮,饲养8周,养殖试验结束后,用10 mg/L的NH4Cl对24尾鱼进行氨氮应激试验。试验结果显示,氨氮应激前,0.4% FOS组的血液谷丙转氨酶(GPT)活性显著低于对照组和0.8% FOS组,在氨氮应激后,0.4% FOS组的GOT活性有降低的趋势,但仅在24 h时显著低于0.8%组,0.8%组和对照组之间并无显著差异;血清酸性磷酸酶、补体3(C3)、补体4(C4)、酚氧化酶都出现先升高后降低的趋势,分别在应激后的3、24、12、12 h达到最大值。此外,应激前0.4% FOS组补体C3、C4、酚氧化酶的活性都显著高于对照组,应激后0.4%FOS组C3在3 h处显著高于对照组,但是在其它时间点各组差异并不显著,果寡糖添加水平和采样时间对这些免疫指标都无显著的交互作用。结果表明饲料中添加0.4%的果寡糖能够提高团头鲂的免疫力指标活性,提高了团头鲂抗氨氮应激的能力。
团头鲂(Megalobramaamblycephala);果寡糖;氨氮应激;免疫
氨氮是水产养殖环境中重要的污染胁迫因子之一,在水体中以离子氨(NH4+)和非离子氨(NH3)2两种形态存在,它们之间可以相互转换,其中非离子氨因为不带电荷,具有较强的脂溶性,能够穿透细胞膜,表现出毒性效应[1]。水体中过高的氨氮和亚硝酸盐氮能对鱼虾产生直接毒害作用,因为外界高浓度氨氮,会使鱼类血液中氨氮浓度迅速升高而对鱼类产生很大的毒害[2]。池水积累一定量非离子氨会对鱼鳃表皮细胞造成损伤而使鱼的免疫力降低[3],许多学者研究认为在低于致死浓度的条件下,氨氮对鱼、虾鳃组织和生理功能(如氧消耗、氨排泄、渗透调节等)具有显著影响[4],近年来,由于高密度、集约化的养殖模式,水产动物排泄物的氨化作用增强,饵料利用率低,殘饵剩余较多,再加上硬骨鱼类对氨氮毒性又非常敏感[5],容易造成免疫力下降,病原菌感染加剧,死亡率升高。
果寡糖(GPT)是蔗糖分子以β-1,2糖苷键结合几个(8个以下)D-果糖而形成的寡糖,作为有益微生物的一种养分来源,它们可通过选择性刺激肠道中原有或外来菌种的生长和活性来影响宿主,对动物胃肠道微生物区系、免疫等功能有重要影响,在对三角鲂(Megalobramaterminalis)、异育银鲫(Carassiusauratusgibelio)等研究的结果表明果寡糖能显著提高其白细胞吞噬活性、血清溶菌酶活力、血清超氧化物歧化酶活力和补体含量[6-8],但研究果寡糖在应激条件下,对鱼类免疫状况的影响至今还是空白。本实验以团头鲂为研究对象,研究果寡糖在急性氨氮胁迫下对团头鲂(Megalobramaamblycephala)非特异性免疫反应的影响。为团头鲂养殖水环境调控和病害防治提供科学依据。
1 材料与方法
1.1试验饲料
基础日粮饲料配方及养分水平如表1所示。对照组投喂基础日粮,试验组饲料是在基础日粮中分别添加0.4%、0.8%的果寡糖,其中,果寡糖由日本明治集团提供,果寡糖由蔗果三糖(GF2)、蔗果四糖(GF3)、蔗果五糖(GF4)组成,且GF2+GF3+GF4≥95%,其它成分<5%;饲料原料购自江苏帅丰饲料公司,原料粉碎后,均过60目筛,用逐级扩大法混匀后制成直径为1.5 mm的颗粒饲料,自然风干后,放置于﹣20 ℃冰箱中保存备用。
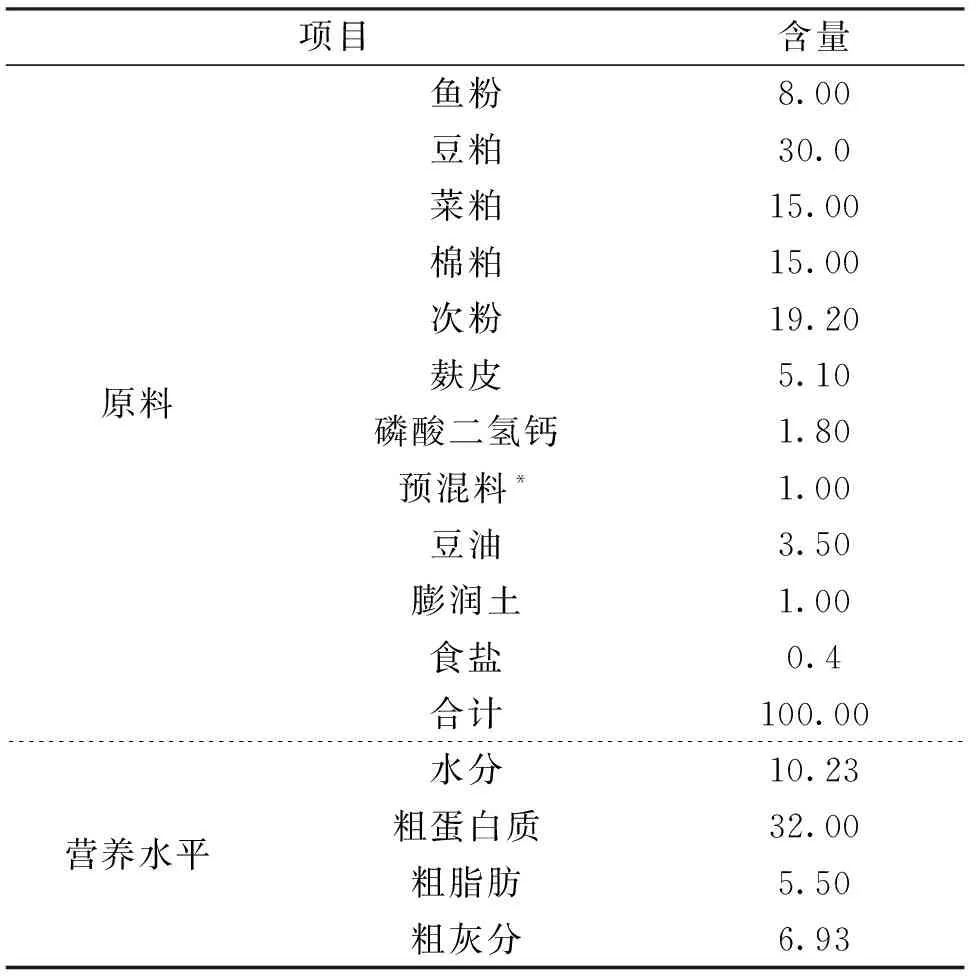
表1 基础日粮组成及营养水平(风干基础)Tab.1 Composition and nutrient levels of basal diets(air dry basis) %
注:每千克饲料包含以下矿物质和维生素:CuSO4·5H2O,20 mg;FeSO4·7H2O,250 mg;ZnSO4·7H2O,220 mg;MnSO4·4H2O,70 mg;Na2SeO3,0.4 mg;KI,0.26 mg;CoCl2·6H2O,1 mg;VA,9 000 IU;VD,2 000 IU;VE,45 mg;VK3,2.2 mg;VB1,3.2 mg;VB2,10.9 mg;烟酸 28 mg;VB5,20 mg;VB6,5 mg;VB12,0.016 mg;VC,50 mg;泛酸,10 mg;叶酸,1.65 mg;胆碱,600 mg。
1.2试验鱼与饲养管理
试验用鱼团头鲂由江苏南京浦口基地提供。试验鱼在网箱中驯养15 d 后,选择体质健壮,规格一致的幼鱼,初始体质量(13.5±0.5)g的团头鲂360尾,随机分成3组,其中1组为对照组,另外2组为试验组,每组4重复,每个重复30尾鱼,分别饲养于室外网箱(规格:1 m×1 m×1 m)中,每天投饵量为鱼体重的3%~6%,根据吃食情况进行适度调整,每天投喂3次,投喂时间分别为7:30、12:30、17:30,试验期间保持水体pH 7.0~8.0,溶氧>5.0 mg/L,氨氮<0.3 mg/L,亚硝酸盐<0.1 mg/L,以保证水质优良。每天对天气、水温、摄食、死鱼等情况进行记录,养殖周期为8周。
1.3氨氮应激试验
饲养结束后,把鱼转移到水族箱中,稳定2 d后,用氯化铵调制水中氨氮质量浓度10 mg/L,氨氮浓度的测定用次溴酸盐氧化法,每隔2 h测一次水中氨氮浓度和pH值,保证其基本稳定,分别于应激前0 h和应激后3、6、12、24、48 h进行采样。
1.4样品采集与测定
分别在应激前后的各个时间点随机从每缸中取3尾鱼,用MS-222(100 mg/L)进行麻醉,尾静脉采血,放入用一定浓度的肝素钠润过的离心管中。血样于4 ℃冰箱中静置2 h,然后在4 ℃,3000 g离心10 min,上清液移置-20 ℃冰箱中保存备用。
谷丙转氨酶(GPT)和谷草转氨酶(GOT)的测定采用葡萄糖氧化酶法,碱性磷酸酶(AKP)和酸性磷酸酶(ACP)的测定采用比色法,补体3(C3)、补体4(C4)和酚氧化物酶(PO)含量的测定采用的酶联免疫吸附法,以上试剂盒均购自南京建成生物有限公司,详细操作步骤见说明书。
1.5数据处理
试验数据用SPSS 16.0软件进行处理,用双因素方差分析(Two-way ANOVA)的方法分析果寡糖水平和采样时间是否存在交互作用,若交互显著则采用Tukey′s多重比较法检验各组间差异,P<0.05表示差异显著。实验结果以平均值±标准误(Mean±SEM)表示。
2 结果
2.1果寡糖对团头鲂血清谷丙转氨酶和谷草转氨酶活性的影响
由图1,2可知,在氨氮应激下,各组团头鲂血清谷丙转氨酶(GPT)和谷草转氨酶(GOT)的活性呈升高趋势。在应激前,各组GOT水平的差异不显著,GPT活性 0.4% FOS组显著低于对照组和0.8%FOS组。应激后,0.4%添加组的GPT活性有的趋势,但仅在24 h时显著低于0.8%组,0.8%组和对照组之间并无显著差异,另外,FOS添加水平和采样时间不存在交互作用。

图1 氨氮应激下果寡糖对团头鲂血液 谷草转氨酶活性的影响Fig.1 Effects of FOS on plasma GOT levels of blunt snout bream under ammonia stress
大写字母表示同一个组在不同时间点的变化情况,小写字母表示同一时间点各组之间的差异。(注释下同)

图2 氨氮应激下果寡糖对团头鲂血液 谷丙转氨酶活性的影响Fig.2 Effects of FOS on plasma GPT levels of blunt snout bream under ammonia stress
2.2果寡糖对团头鲂血清碱性磷酸酶和酸性磷酸酶活性的影响
由图3,4可知,在氨氮应激下,各组血清碱性磷酸酶出现显著降低趋势,并在24~48 h几乎趋于稳定;而酸性磷酸酶出现先升高后降低的趋势,在3~6 h达到最高;饲料中添加FOS后,AKP、ACP活性都有所升高,但各组在采样前后差异均不显著,且双因素分析得出果寡糖添加水平和采样时间并无交互作用。
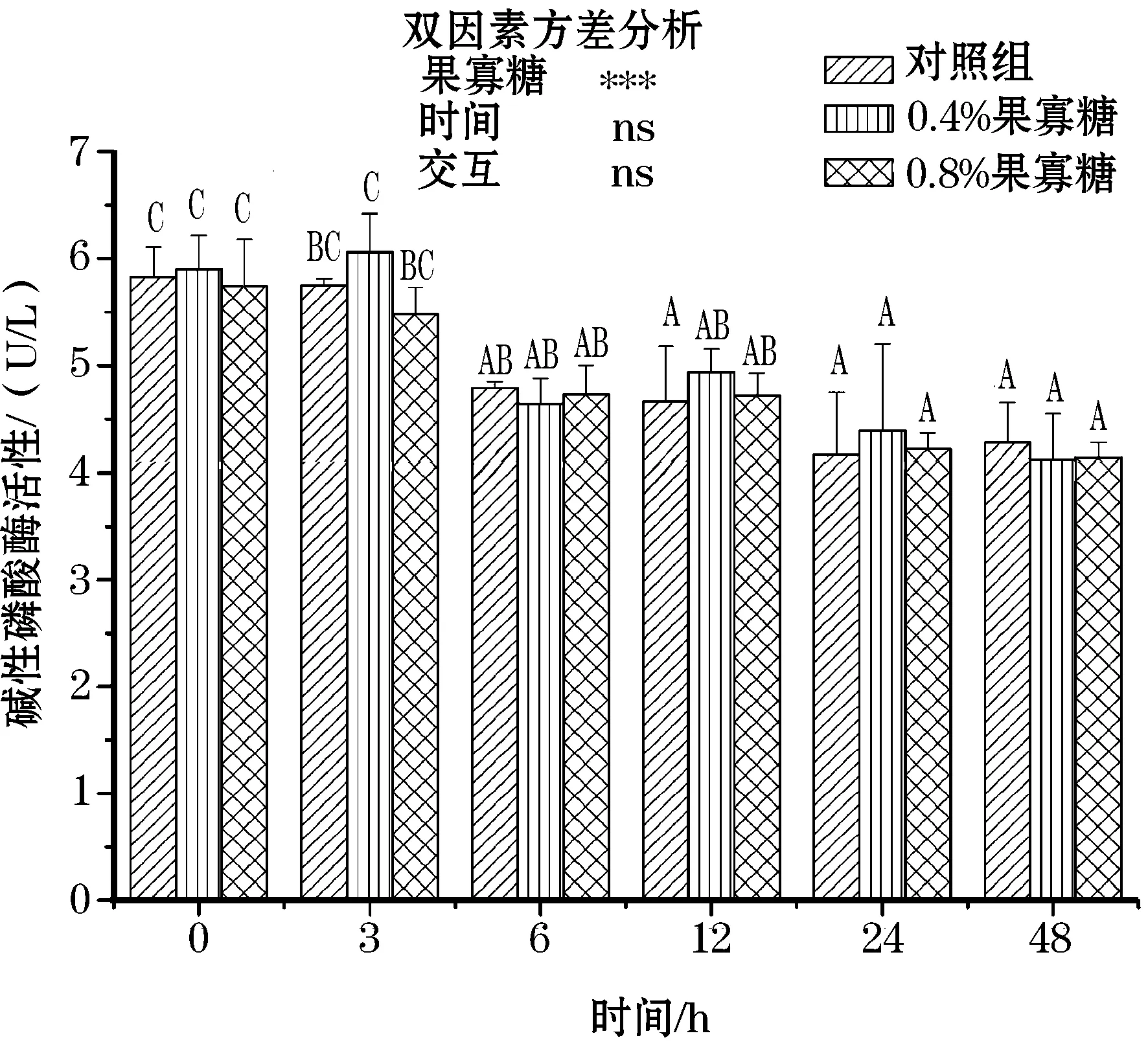
图3 氨氮应激下果寡糖对团头鲂血液 碱性磷酸酶含量的影响Fig.3 Effects of FOS on plasma AKP levels of blunt snout bream under ammonia stress
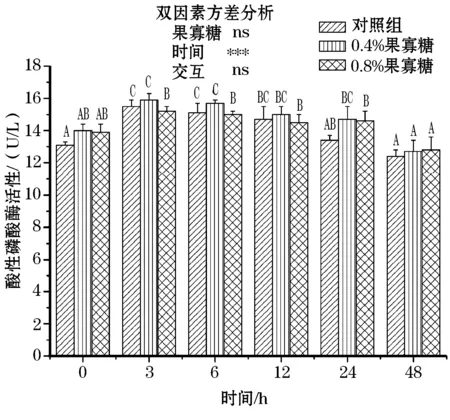
图4 氨氮应激下果寡糖对团头鲂血液 酸性磷酸酶活性的影响Fig.4 Effects of FOS on plasma ACP levels of blunt snout bream under ammonia stress
2.3果寡糖对团头鲂血清补体C3、C4和酚氧化物酶的影响
由图5,6可知,在氨氮应激下各组血清补体C3在0~24 h呈升高趋势,而C4在0~12 h呈升高趋势,之后出现了降低趋势;添加0.4% FOS组在应激前C3、C4都显著高于对照组,且0.8% FOS组C3出现相似结果,氨氮应激后添加0.4%FOS组C3、C4都有升高趋势,但是各组差异并不显著;由图7得出,氨氮应激前,酚氧化物酶(PO)在0.4%组显著高于对照组,而0.8%组和对照组无显著差异,氨氮应激后对照组呈先升高后降低的趋势,但其它两组并无显著变化,且FOS水平和采样时间对C3、C4和PO都无交互作用。

图5 氨氮应激下果寡糖对团头鲂血液补体C3的影响Fig.5 Effects of FOS on plasma C3 levels of blunt snout bream under ammonia stress

图6 氨氮应激下果寡糖对团头鲂血液补体C4的影响Fig.6 Effects of FOS on plasma C4 levels of blunt snout bream under ammonia stress
3 讨论
3.1果寡糖对团头鲂血清谷丙转氨酶和谷草转氨酶活性的影响
谷丙转氨酶和谷草转氨酶是在动物组织细胞内参与氨基酸合成与分解的重要氨基酸转移酶,在谷氨酸和丙氨酸的转化中起着催化剂的作用,对蛋白质的代谢起着重要作用,它们的变化在一定程度上能够反映机体肝细胞和心肌细胞的受损程度[9],机体在正常情况下,血清中只有很少量的谷丙转氨酶,但是但机体的肝组织受到损伤时,细胞膜的通透性会随之增加,细胞浆内的谷丙转氨酶释放到血液中,本研究得出谷丙转氨酶和谷草转氨酶都随着氨氮应激时间的延长呈现先增高后降低的趋势,并在应激后的12 h达到最大值,这可能是由于高浓度的氨氮胁迫导致机体脂质过氧化物增多,肝细胞受到损伤,影响了肝细胞的正常的生理功能,Jeney等[10]研究表明鲤(Cyprinuscarpio)受到高浓度氨氮胁迫后,血清谷丙转氨酶活力出现类似的趋势,胡毅等[11]的研究指出当用10 mg/L或20 mg/L的氨氮胁迫青鱼时,其血清也出现谷丙转氨酶升高的趋势。这可能是因为随着应激时间的延长,破坏鱼类血清酸碱平衡和血清离子的衡态,影响了鱼体内环境的稳定。非特异性免疫系统遭到破坏,免疫力下降,病原菌的易感染性也增强。但是在添加果寡糖0.4%组谷丙转氨酶和谷草转氨酶的活性与对照组相比,都有一定程度的降低,这表明果寡糖在氨氮应激下有保护肝细胞和心肌细胞作用。

图7 氨氮应激下果寡糖对团头鲂血液酚氧化物酶的影响Fig.7 Effects of FOS on plasma PO levels of blunt snout bream under ammonia stress
3.2果寡糖对氨氮应激下团头鲂血清酸性磷酸酶和碱性磷酸酶活性的影响
酸性磷酸酶在体内直接参与磷酸基团的转移和代谢,间接增强机体的非特异性免疫能力,碱性磷酸酶是生物体内的一种重要的代谢调控酶,直接参与磷酸基团的转移和钙磷代谢,在鱼类对营养物质的吸收与利用过程中发挥着重要作用。另外,碱性磷酸酶能够改变病原体的表面结构,从而增强机体对病原体的识别和吞噬能力[12]。本实验得出血清中酸性磷酸酶的活性呈先升高后下降的变化趋势。碱性磷酸酶一直处于降化可能是因为鱼生活在氨氮浓度较高的环境中,机体会进行一些适应性调节。在适当的环境胁迫下,可能会刺激一些酶活性的表达,酶的活性增强,但是随着应激时间的延长,某些酶的构象会发生变化,不利于酶与底物的结合,从而导致活力下降,这说明机体对外界不良刺激的抵抗力下降,机体很容易受到外界不利因素的刺激而引发疾病,环境因子对组织中酶活性的影响及其作用机制还有待于进一步研究。本研究得出在应激前后血清酸性磷酸酶和碱性磷酸酶活性在0.4%果寡糖组显著高于对照组和0.8%组,这表明果寡糖可以提高团头鲂的非特异性免疫,增强其抵抗氨氮应激的能力。这可能与果寡糖促进了益生菌比如芽孢杆菌和乳酸菌的生长和增殖有关[8],而这些细菌的细胞壁的组成成分比如一些脂多糖等都具有增强免疫功能并能刺激机体的免疫反应,提高免疫力,另外,这些微生物能够促进免疫因子和免疫球蛋白的分泌,增强机体的免疫活性[13]。
3.3果寡糖对氨氮应激下团头鲂血清补体C3、C4和PO活性的影响
血清补体活性已被证实是一项非常重要的非特异性防御指标,能够保护机体抵御病原菌的感染[14],补体C3、C4是补体系统中的固有成分,有调节机体非特异性免疫的功能[15],本研究发现随着应激时间的延长,补体C3、C4都出现先升高后降低的趋势,这可能是因为鱼类在受到氨氮胁迫后,引起了鱼体的急性应激反应[9],其通过大量分泌皮质醇来促进糖的合成、脂肪降解以获得能量,并通过增加特定蛋白(如溶菌酶、补体和C反应蛋白等)的水平来增强机体的免疫力[9,16],共同抵抗病原菌的侵袭,从而导致血清中某些蛋白的水平升高,当应激强度超出鱼体防御机能时,这些蛋白的合成能力就会减弱,从而表现为某些酶的活性降低。周显青等[17]研究结果表明,应激能够降低中华幼鳖补体活性和含量,鱼类受到氨氮应激后,鱼体的免疫机能会受到影响,但是影响程度还会因种类的不同而有所差异[18]。
酚氧化酶是重要的免疫应答因子,在异物识别中起到重要作用。它在机体受到外界异物侵袭或者外界环境发生变化时,细胞会通过胞吐作用把酚氧化酶原释放到周围介质中,从而使酚氧化酶被激活,活性升高[19]。本实验结果表明酚氧化酶随着应激时间的延长呈现先升高后降低的趋势,在应激后的12 h出现最大值,这表明氨氮急性应激刺激机体发生应激反应,神经内分泌系统分泌生物胺等内分泌因子,诱发启动了proPO系统,酚氧化酶原系统释放到周围的细胞间质中,出现酶活性升高的现象,这与投喂芽孢杆菌的斑节对虾在注射嗜水气单胞菌后血液酚氧化酶活性有升高的现象相似。但是随着应激时间的增长,酶活性出现下降趋势,这可能是由于长时间的氨氮应激对酚氧化酶原的颗粒细胞造成了损伤[20],导致PO活性的降低,黄鹤忠等[21]研究表明高浓度的氨氮随着胁迫时间的延长,中华绒螯蟹(Eriocheirsinensis)血液中溶菌酶、酚氧化酶和超氧化酶的活性都出现下降的趋势,机体的非特异免疫系统遭到损害,机体的细胞和组织都受到一定程度的损伤,由以上结果可以得出,氨氮对酚氧化酶的影响是一个比较复杂的过程,在一定程度上反映了机体的健康状况,其作用机制还需进一步的研究。
饲料中0.4%的果寡糖提高了血清中C3、C4的含量和酚氧化物酶的活性,增强了团头鲂的免疫功能,并提高了抗氨氮应激的能力。
[1]姜令緖,潘鲁青,肖国强.氨氮对凡纳对虾免疫指标的影响[J].中国水产科学,2004,11(6):537-541.
[2]Ip Y,Chew S,Leong I,et al.The sinensis(Family Eleotridae) stores glutamine and reduces ammonia Production during aerial exposure[J].J ComP Physiol Bioehem Syst Environ Physiol,2001,171( 5):357-367.
[3]Colt J E,Armstrong D A.Nitrogen toxicity to crustaceans,fish,and mollusks[J].American Fisheries Society and North East Soeiety of Conservation Engineers,1981,12(2):34-47.
[4]Chen J C,Lin C Y.Oxygen consumption and ammonia-N excretion ofPenaeuschinensisjuveniles exposed to ambientammonia at different salinity levels[J].Comp Biochem Physiol,1992,102(2):287-291.
[5]Handy R D,Poxton M G.Nitrogen pollution in mariculture:toxicity and excretion of nitrogenous compounds by marine fish[J].Rev Fish Biol Fisher,1993,3(3):205-241.
[6]Soleimani N,Hoseinifar S H,Merrifield D L,et al.Dietary supplementation of fructooligosaccharide (FOS) improves the innate immune response,stress resistance,digestive enzyme activities and growth performance of Caspian roach (Rutilusrutilus) fry[J].Fish Shellfish Immunol,2012,32(2):316-321.
[7]Zhang C N,Li X F,Xu W N,et al.Combined effects of dietary fructooligosaccharide andBacilluslicheniformison innate immunity,antioxidant capability and disease resistance of triangular bream (Megalobramaterminalis)[J].Fish Shellfish Immunol,2013,35(5):1380-1386.
[8]Zhang Q,Ma H,Mai K S,et al.Interaction of dietaryBacillussubtilisand fructooligosaccharide on the growth performance,non-specific immunity of sea cucumber,Apostichopusjaponicus[J].Fish Shellfish Immunol,2010,29(2):204-211.
[9]Yuan W,Li X,Kang J Y,et al.Effect of beta-cypermethrin on GPT and GOT activities of crucian serum[J].Agricul Sci Technol,2005,1(13):198-203.
[10]Jeney G,Nemcsok J,Jenet Z,et al.Acute effect of sublethal ammonia concentrations on common carp (CyprinuscarpioL.).II.Effect of ammonia on blood plasma transaminases (GOT,GPT),G1DH enzyme activity,and ATP value[J].Aquaculture,1992,104(1-2):149-156.
[11]胡毅,黄云,钟蕾,等.氨氮胁迫对青鱼幼鱼鳃丝 Na+/K+-ATP 酶,组织结构及血清部分生理生化指标的影响[J].水产学报,2012,36(4):538-545.
[12]明建华,谢骏,徐跑,等.大黄素,维生素C及其配伍对团头鲂抗拥挤胁迫的影响[J].水生生物学报,2011,35(3):400-413.
[13]Yasui H,Ohwaki M.Enhancement of immune response in peyer's patch cells cultured withBifidobacteriumbreve[J].J Dairy Sci,1991,74(4):1187-1195.
[14]Tort L,Gomez E,Montero D,et al.Serum haemolytic and agglutinating activity as indicators of fish immunocompetence:their suitability in stress and dietary studies[J].Aquacult Int,1996,4(1):31-41.
[15]Holland M C H,Lambris J D.The complement system in teleosts[J].Fish Shellfish Immunol,2002,12(5):399-420.
[16]Mock A,Peters G.Lysozyme activity in rainbow trout,Oncorhynchusmykiss (Walbaum),stressed by handling,transport and water pollution[J].J Fish Biol,1990,37(6):873-885.
[17]周显青,牛翠娟,孙儒泳.黄芪和酸应激对中华鳖幼鳖血清补体C3和C4含量的影响[J].动物学研究,2002,23(2):177-180.
[18]Bowden T J,Thompson K D,Morgan A L,et al.Seasonal variation and the immune response:a fish perspective[J].Fish Shellfish Immunol,2007,22(6):695-706.
[19]Sung H H,Hwang S F,Tasi F M.Responses of giant freshwater prawn (Macrobrachiumrosenbergii) to challenge by two strains ofAeromonasspp[J].J Invertebr Pathol,2000,76(76):278-284.
[20]Sequeira T,Vilanova M,Baldaia L,et al.Flow cytometric analysis of molt-related changes in hemocyte type in male and femalePenaeusjaponicus[J].Biolo Bull,1995,189(3):376-380.
[21]黄鹤忠,李义,宋学宏,等.氨氮胁迫对中华绒螯蟹(Eriocheirsinensis)免疫功能的影响[J].海洋与湖沼,2006,37(3):198-205.
(责任编辑:邓薇)
Effects of fructooligosaccharide on nonspecific immunity of blunt snout bream (Megalobrama amblycephala) under high ammonia stress
ZHANG Chun-nuan1,REN Hong-tao1,ZHANG Ji-liang1,WU Qiu-jue1,GAO Xiao-chan1,LIU Wen-bin2
(1.CollegeofAnimalScienceandTechnology,HenanUniversityofScientificandTechnology,Luoyang471003,Henan,China;2.CollegeofAnimalScienceandTechnology,NanjingAgriculturalUniversity,Nanjing210095,China)
This study aimed to determine the effects of fructooligosaccharide (FOS) on immune response of blunt snout bream (Megalobramaamblycephala) under high ammonia stress.A total of 360 fish were randomly distributed into three groups (each with four replicates) and were fed three levels of FOS (0,0.4% and 0.8%) for 8 weeks.After the feeding trial,24 fish per tank were exposed to ammonia at 10 mg/L.From the results,we found that Glutamic Pyruvic Transaminase (GPT) levels of fish fed 0.4% FOS were all significantly lower than that of the other groups at 0h and 24 h.After stress,Plasma acid phosphatase (ACP),complement 3 (C3),complement 4 (C4) and phenoloxidase (PO) activities all increased significantly with the maximum levels being attained at 3,24,12 and12 h,respectively.Thereafter,these parameters all decreased significantly.In addition,fish fed 0.4% FOS showed higher C3,C4and PO compared to that of control group before stress.Whereas,only the C3of fish fed 0.4% FOS showed higher than that of control group at 3 h after stress.There was no significant difference at other times and there was no significant interaction between the FOS levels and the time.The results indicated that the supplementation of 0.4% FOS could increase the non-specific immunity of blunt snout bream,and enhance its resistance to high ammonia stress.
Megalobramaamblycephala;fructooligosaccharide;ammonia stress;immunity
2015-12-16;
2016-06-20
国家大宗淡水鱼类产业体系-鲤和团头鲂营养与饲料岗位(CARS-46-20)
张春暖(1987-),女,河南平顶山人,讲师,从事水产动物营养与饲料的研究。E-mail:zhangchunnuan12@163.com
刘文斌。E-mail:wbliu@njau.edu.cn
S963.73
A
1000-6907-(2016)05-0064-06
