急性精神应激与生理应激对小鼠行为及学习记忆能力的影响对比观察
2016-08-31徐锋董惠郝珂杨诚高蒙熊雅静赖宇航卞国强孟晶刘金东
徐锋,董惠,郝珂,杨诚,高蒙,熊雅静,赖宇航,卞国强,孟晶,刘金东
(1徐州医科大学麻醉学院,江苏徐州221002;2徐州医科大学江苏省麻醉学重点实验室)
急性精神应激与生理应激对小鼠行为及学习记忆能力的影响对比观察
徐锋1,董惠1,郝珂1,杨诚1,高蒙1,熊雅静1,赖宇航1,卞国强1,孟晶2,刘金东2
(1徐州医科大学麻醉学院,江苏徐州221002;2徐州医科大学江苏省麻醉学重点实验室)
目的观察并比较急性精神应激与生理应激对小鼠行为及学习记忆能力的影响。方法昆明种小鼠24只,随机分为生理应激组、精神应激组和对照组,每组各8只。生理应激组小鼠在MG-3Y型迷宫内接受电击,精神应激组小鼠旁观电击过程,对照组小鼠正常饲养。记录各组小鼠排便、排尿及修饰动作总次数。应激2 h后进行旷场实验,记录小鼠中央格爬行次数、外周格爬行次数、中央格百分比、排便粒数、修饰动作发生次数。采用Morris水迷宫进行定位航行实验和空间搜索实验,评价小鼠的学习记忆能力。结果应激过程中生理应激组小鼠的恐惧行为较精神应激组小鼠更为剧烈。在旷场实验中,精神应激组小鼠直立次数、修饰次数、排便粒数少于对照组(P均<0.05);生理应激组小鼠中央格及外周格爬行次数较精神应激组和对照组显著减少(P均<0.05);生理应激组小鼠中央格爬行次数、中央格百分比少于对照组,直立次数、修饰次数、排便粒数多于精神应激组(P均<0.05)。在Morris水迷宫实验中,精神应激组与生理应激组在不同观察时点逃避潜伏期呈现出先延长后缩短趋势,其中第12次训练时精神应激组逃避潜伏期最长,对照组次之,生理应激组最短(P均<0.05)。精神应激组小鼠平台搜索策略多为随机式;生理应激组和对照组小鼠的平台搜索策略多为边缘式和直线式。结论急性精神应激与生理应激对小鼠行为和学习记忆的影响存在较大差异。急性精神应激小鼠表现为抑郁状态,而急性生理应激小鼠表现为空间探索和活动能力减弱。急性精神应激会干扰学习记忆能力,而急性生理应激会增强学习记忆能力。
应激反应;精神应激;生理应激;旷场实验;Morris水迷宫;动物实验
多项研究[1,2]发现,应激可影响观察对象的学习记忆能力。既往建立动物应激反应模型的方法主要有电击足底、限制性应激、束缚应激、强迫游泳等,这些在一定程度上均含有生理与精神应激的成分,不能很好地模拟现实生活中人类的精神应激[3]。上述造模方法未把精神应激与生理应激进行严格区分和比较。2015年6~7月,本研究分别建立了急性精神应激与生理应激小鼠模型,观察并比较了两种应激方式对小鼠行为及学习记忆能力的影响。
1 材料与方法
1.1实验动物与材料同批次昆明种小鼠24只,雌雄不限,由徐州医科大学实验动物中心提供。实验前2 d使小鼠熟悉实验室环境,自由摄食、饮水。MG-3Y型迷宫刺激器及Y型迷宫购自安徽濉溪正华教学实验器械厂,小鼠Morris水迷宫装置、旷场实验装置(由不透明材料制成,长、宽、高各50 cm,底面等分为25个方格,敞箱底部及四周均为黑色)及ANY-maze version4.84系统(serial number:A2ST-3PJW-JS8W-TXKU)购自上海软隆科技发展有限公司。
1.2动物分组与应激方法将24只小鼠随机分为精神应激组、生理应激组和对照组,每组各8只。生理应激组小鼠在MG-3Y型迷宫内接受电击,位于三个通电臂内,给予电压36 V、周期5 s、间歇55 s,小鼠产生惊叫、惊跳、排便、排尿等反应;精神应激组不接受电击,在MG-3Y型迷宫中央旁观生理应激组小鼠遭受电击的过程,通过视觉、听觉、味觉等产生精神应激,每次应激在上午9:00~11:00进行,每次持续1 h、每天1次、共进行2 d;对照组小鼠放入另一个MG-3Y型迷宫,但不给予电刺激。
1.3小鼠行为评估方法精神应激组与生理应激组于最后一次应激过程中(对照组于同一时间)记录小鼠排便、排尿及修饰动作(抓痒、洗脸、舔足等)总次数。急性应激结束2 h后进行旷场实验,评价小鼠在新异环境中的自主行为、探究行为与紧张度。依次将小鼠轻轻放置于旷场实验敞箱正中央格内,观察5 min内活动情况,记录小鼠中央格爬行次数、外周格爬行次数、中央格百分比(中央格数/总格数)、排便粒数、修饰动作发生次数。当小鼠三爪或四爪跨入邻格方能计数,并用ANY-maze version4.84系统监测、记录、分析。
1.4小鼠学习记忆能力评估方法旷场实验结束后进行Morris水迷宫实验[4]。迷宫由圆形水池、平台及图像采集处理系统组成。水池直径80 cm、高30 cm,以两条垂直的直径为界限人为地将圆形水池等分为四个象限,平台位于第三象限中间位置。在第三象限池壁中间放置一白色标记。水温(22±1)℃,水位高出平台1 cm。采用定位航行实验和空间搜索实验评估小鼠的学习记忆能力。①定位航行实验:将小鼠从四个象限中心放入水中,记录小鼠入水至找到平台并停留10 s所用时间(即逃避潜伏期),若在平台停留不足10 s再次入水则继续计时;若小鼠60 s内未找到平台,则引其上平台停留10 s,逃避潜伏期记为60 s;实验第1、2天共进行4个周期训练,每周期训练4次,分别于上午8:00~11:00及下午2:00~5:00进行,记录第4、8、12、16次训练时逃避潜伏期。②空间搜索实验:实验第3天8:00~10:00进行空间搜索实验,取出平台,将第一象限作为入水点把小鼠放入水中,记录小鼠60 s内在水迷宫第一、二、三、四象限的游泳时间。由图像自动采集处理系统生成小鼠游泳路线图,据此分析小鼠搜索平台的策略。搜索策略主要分为边缘式、随机式、直线式、趋向式。以小鼠运动区域中心为圆心,取半径的75%画圆,若小鼠70%以上时间均在该圆外活动,判定为边缘式策略;以小鼠入水点与平台中心连线为中轴,若小鼠所有运动轨迹与中轴距离不超过半径的15%,且在该区域活动时间占总运动时间的70%以上,则判定为直线式策略;以小鼠入水点与平台中心连线为中轴,若小鼠所有运动轨迹与中轴距离不超过半径的50%,且在该区域活动时间占总运动时间的70%以上,则判定为趋向式策略;若实验动物的运动方式与以上三者不同称为随机式策略。痴呆小鼠的搜索策略将发生异常,如只有边缘式→随机式或随机式→边缘式,趋向式、直线式少见。
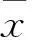
2 结果
2.1各组小鼠行为变化生理应激组小鼠在接受电击时,出现尖叫、挣扎跳跃、排便、排尿等恐惧行为;精神应激组小鼠旁观生理应激组小鼠接受电击时,表现木僵、颤栗、惊恐注视、排便、排尿、频繁修饰等应激行为。生理应激组小鼠的恐惧行为较精神应激组小鼠更为剧烈。将生理应激组小鼠第二次放入电击箱时,小鼠极力挣扎,出现排便、排尿现象,表明小鼠已形成恐惧记忆。生理应激组小鼠排尿次数、修饰次数、排便粒数分别为20次、258次和47粒,精神应激组分别为12次、195次、31粒,对照组分别为11次、161次、32粒,生理应激组、精神应激组小鼠排尿次数、修饰次数较对照组增多,其中生理应激组多于精神应激组(P均<0.05)。旷场试验中,精神应激组小鼠直立次数、修饰次数、排便粒数少于对照组(P均<0.05);生理应激组小鼠中央格及外周格爬行次数较精神应激组和对照组显著减少(P均<0.05);生理应激组中央格爬行次数、中央格百分比少于对照组,直立次数、修饰次数、排便粒数多于精神应激组(P均<0.05)。详见表1。
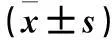
表1 各组小鼠旷场实验结果
注:与对照组相比,#P<0.05,##P<0.01;与精神应激组相比,*P<0.05,**P<0.01。
2.2各组小鼠学习记忆能力变化精神应激组与生理应激组小鼠第4、8、12、16次训练时逃避潜伏期呈现出先延长后缩短的趋势,其中第12次训练时精神应激组逃避潜伏期最长,对照组次之,生理应激组最短(P均<0.05)。见表2。空间搜索实验结果显示,三组小鼠在水迷宫不同象限游泳时间差异无统计学意义(见表3)。精神应激组小鼠平台搜索策略多为随机式;生理应激组和对照组小鼠的平台搜索策略多为边缘式和直线式。

表2 应激后不同时间各组小鼠逃避潜伏期比较±s)
注:与对照组相比,*P<0.05;与精神应激组相比,#P<0.05。
3 讨论
精神应激因素为情感、心理方面,生理应激因素侧重躯体方面[5]。生理应激因素有别于精神应激因素[3],以往常用的精神应激模型制作方法未把精神应激与生理应激进行严格区分和比较。急性应激状态下,海马形态结构不发生明显变化,神经元结构功能未受损伤[6]。急性生理应激对中枢神经系统的影响可能不涉及到器质性改变;但较为强烈而持久的慢性应激足以损害海马组织结构,严重影响学习记忆能力[7,8]。Jeon等[9]对旁观电击鼠模型进行研究发现,在旁观学习中,有前扣带回皮层(ACC)及杏仁体参与。Kim等[10]也指出“旁观恐惧学习”的过程受ACC支配。以上研究结果表明,急性精神应激与生理应激的神经内分泌、神经解剖、神经电生理等机制均存在明显差异[11~13]。

表3 各组小鼠在水迷宫不同象限游泳次数(次,±s)
本研究分别通过不同方式制作生理应激与精神应激小鼠模型,发现生理应激组小鼠的恐惧行为较精神应激组小鼠更为剧烈。生理应激组、精神应激组小鼠排尿次数、修饰次数较对照组增多,其中生理应激组多于精神应激组。这说明旁观电击造模可以使小鼠产生精神应激和焦虑行为,而这种精神应激模型的应激强度和焦虑水平都远低于生理应激模型。在旷场实验中,精神应激组小鼠直立次数、修饰次数及排便粒数较少,提示精神应激小鼠处于一种抑郁状态,对于陌生环境(旷场实验箱)并未表现出较多的焦虑行为。生理应激组小鼠中央格爬行次数、外周格爬行次数、中央格百分比较少,提示急性生理应激小鼠的空间活动和探索能力减弱,更趋向在外周格活动。
我们利用Morris水迷宫实验,从四个时点观察应激小鼠学习记忆能力的变化,结果显示,在定位航行实验中,各组小鼠的逃避潜伏期呈现出先延长后缩短的趋势,其中第12次训练时精神应激组逃避潜伏期最长,对照组次之,生理应激组最短;在空间搜索实验中,各组小鼠在水迷宫不同象限的游泳时间无统计学差异。这些结果提示应激对小鼠学习记忆能力的影响有一定时间依赖性,应激水平随时间延长会出现衰减,从而减弱或消除对小鼠学习记忆能力的干扰;同时,急性生理应激可增强小鼠的学习记忆能力,而急性精神应激会干扰学习记忆能力。我们还发现,生理应激组及对照组小鼠的平台搜索策略较为规则有效,而精神应激组小鼠的搜索策略较不规则,这也进一步验证了急性精神应激对小鼠学习记忆能力的干扰作用,与之前的研究[14,15]结果一致。
结合上述研究结果,我们认为,急性精神应激与生理应激对小鼠行为和学习记忆的影响存在较大差异。急性精神应激小鼠更多表现为一种抑郁状态,而急性生理应激小鼠表现为空间探索和活动能力减弱。急性精神应激会干扰学习记忆能力,而急性生理应激会增强学习记忆能力。
[1] Mika A,Mazur GJ,Hoffman AN,et al.Chronic Stress Impairs Prefrontal Cortex-Dependent Response Inhibition and Spatial Working Memory[J].Behav Neurosci,2012,126(5):605-619.
[2] Rei D,Mason X,Seo J,et al.Basolateral amygdala bidirectionally modulates stress-induced hippocampal learning and memory deficits through a p25/Cdk5-dependent pathway[J].Proc Natl Acad Sci U S A,2015,112(23):7291-7296.
[3] 李功迎,李凌江,马洪霞.精神应激与生理应激对大鼠焦虑行为的影响[J].中国行为医学科学,2008,17(12):1067-1068.
[4] Wise LE,Long KA,Abdullah RA,et al.Dual Fatty Acid Amide Hydrolase and Monoacylglycerol Lipase Blockade Produces THC-Like Morris Water Maze Deficits in Mice[J].ACS Chem Neurosci,2012,3(5):369-378.
[5] Stults-Kolehmainen MA,Sinha R.The Effects of Stress on Physical Activity and Exercise[J].Sports Med,2014,44(1):81-121.
[6] Whitehead G,Jo J,Hogg EL,et al.Acute stress causes rapid synaptic insertion of Ca2+-permeable AMPA receptors to facilitate long-term potentiation in the hippocampus[J].Brain,2013,136(12):3753-3765.
[7] Ferland CL,Harris EP,Lam M,et al.Facilitation of the HPA axis to a novel acute stress following chronic stress exposure modulates histone acetylation and the ERK/MAPK pathway in the dentate gyrus of male rats[J].Endocrinology,2014,155(8):2942-2952.
[8] 程娟,王涛涛,唐伟,等.慢性复合应激致老年小鼠学习记忆损伤及机制研究[J].安徽医科大学学报,2013,48(12):1466-1500.
[9] Jeon D,Kim S,Chetana M,et al.Observational fear learning involves affective pain system and Cav1.2 Ca2+channels in ACC[J].Nat Neurosci,2010,13(4):482-488.
[10] Kim S,Mátyás F,Lee S,et al.Lateralization of observational fear learning at the cortical but notthalamic level in mice[J].Proc Natl Acad Sci U S A,2012,109(38):15497-15501.
[11] Radley JJ,Kabbaj M,Jacobson L,et al.Stress risk factor and stress-related pathology: Neuroplasticity,epigenetics and endophenotypes[J].Stress,2011,14(5):481-497.
[12] 李凌江,陈树林.精神应激的生物学致病机制研究-Ⅲ神经解剖学机制[J].中国行为医学科学,2005,14(3):193-195.
[13] Vitlic A,Lord JM,Phillips AC.Stress,ageing and their influence on functional,cellular and molecular aspects of the immune system[J].Age (Dordr),2014,36(3):1169-1185.
[14] Trammell JP,Clore GL.Does Stress Enhance or Impair Memory Consolidation[J].Cogn Emot,2014,28(2):361-374.
[15] 邹涛,程明,朱熊兆,等.急性心理应激对小鼠记忆的影响[J].中国行为医学科学,2015,14(1):23-25.
江苏省高等学校大学生创新创业训练计划项目(201510313075X)。
孟晶(E-mail:mengjingmz@sina.com)
10.3969/j.issn.1002-266X.2016.19.013
R395.1
A
1002-266X(2016)19-0039-03
2015-12-15)
