炒酸枣仁对焦虑大鼠基底外侧杏仁核神经元信息编码动态变化的影响*
2021-10-09华伊张正一王艳艳黄莉莉叶晓楠李廷利
华伊,张正一,王艳艳,黄莉莉,叶晓楠,李廷利
(黑龙江中医药大学药学院,哈尔滨 150040)
酸枣仁为鼠李科植物酸枣[ZiziphusjujubaMill.Var.spinosae(Bunge)Hu ex H.F.Chou]的干燥成熟种子,主治虚烦不眠、惊悸多梦等症[1],酸枣仁在临床的应用普遍使用炒制品。现代研究发现炒酸枣仁能减少行为学测试中动物的焦虑样行为,具有一定的抗焦虑作用[2-3]。脑科学领域普遍认为杏仁核(amygdala)是参与调节焦虑相关的情绪、行为和生理反应的一个重要脑区[4-5],杏仁核的基底外侧核团(basolateral amygdala,BLA)是焦虑调节过程中接收刺激信息传入的主要部分[6]。杏仁核功能的改变已被认为是焦虑症的一个潜在条件[7-8],焦虑行为的增加与BLA区的过度活跃有关[9]。近年来,BLA区作为神经回路中治疗焦虑相关状态和行为的关键成分被广泛研究[10-12]。笔者在本文将行为学与神经元在体同步电活动相结合,进行了焦虑大鼠的行为学及BLA区神经元信息编码动态变化的研究,同时考察炒酸枣仁对其的干预作用。
1 材料与方法
1.1实验动物 无特定病原体(SPF)级SD大鼠,雄性,体质量(220±20)g,购于黑龙江中医药大学GLP实验动物中心,实验动物生产许可证号:SCXK(黑)2013-004。动物饲养与实验环境为通风、避光、隔音、电磁屏蔽状态,并通过自动定时光控系统进行12 h/12 h明暗光照周期处理(每日7:00—19:00开启照明,19:00至次日7:00关闭照明);室内温度:20~24 ℃;相对湿度:50%~60%;噪声≤40 dB。实验前所有大鼠在本实验室环境内适应7 d,整个实验过程中所有大鼠不限制饮食和饮水。
1.2药物与试剂 酸枣仁,原产地山西,由黑龙江中医药大学王振月教授鉴定为正品,依照文献[13]方法,制备炒酸枣仁和炒酸枣仁水煎液;0.9%氯化钠溶液(哈尔滨三联药业有限公司,批号:170104D5);戊巴比妥钠(国药集团化学试剂有限公司,批号:WS20140104);义齿基托聚合物(上海贝琼齿材有限公司,批号:170308);义齿基托树脂(上海新世纪齿科材料有限公司,批号:20170408)。
1.3仪器与设备 大鼠独立通气笼IVC(型号:RU25H5,苏州市苏杭科技器材有限公司),分析天平(型号:MS105,梅特勒-托利多仪器上海有限公司,感量:0.01 mg),大鼠旷场实验箱(100 cm×100 cm×40 cm,木制),标准脑立体定位仪(型号:51600,美国Stoelting公司),在体多通道神经记录分析系统(型号:OPX-32D,美国Plexon公司),在体单轴微操作仪(型号:IVM-1000,Microelettrica Scientifica China)。
1.4在体多通道记录电极的埋植 选取SD雄性大鼠20只,进行电极埋植手术前,所有大鼠禁食8 h,不禁水。戊巴比妥钠(40 mg·kg-1)腹腔注射麻醉大鼠后,将其固定在标准脑立体定位仪上。根据《George Paxinos & Charles Watson大鼠脑立体定位图谱》(第3版),定位BLA区位置(坐标AP 2.5~2.8 mm,ML 4.5~4.8 mm,DV 7.5 mm),将16(4×4)通道在体记录电极(铂铱合金,电极丝外周涂有绝缘层,直径35 μm)埋置于大鼠的BLA区域,并用义齿基托树脂将电极部分固定在动物颅顶。术后动物恢复4~5 d,于有机玻璃笼内单独饲养。
1.5动物的分组及给药 将已植入电极并经过术后恢复的大鼠经随机数字表法分为模型对照组与炒酸枣仁组,每组10只。炒酸枣仁组每日灌胃给予大鼠炒酸枣仁水煎液(17.5 g·kg-1),模型对照组给予同等体积纯化水,每日1次,连续7 d。
1.6模型的制备及行为学测试 利用旷场实验复制大鼠焦虑模型,每日给药0.5 h后,将各组大鼠放入旷场箱内底面中心,任其自由探索,利用动物行为学采集和分析系统,记录10 min内大鼠在旷场中的行为学表现,每日1次,连续7 d。测试指标为跨格次数、站立次数、进入中央格次数、中央格运动时间(s)和总运动时间(s),并以中央格运动时间除以总运动时间计算中央格/总运动时间(%)。
1.7神经元在体放电信号的采集与处理 每日给药0.5 h后,利用在体多通道神经记录分析系统,先采集各组大鼠在日常饲养环境中BLA区神经元基础放电信号(10 min),再在旷场实验的同时采集旷场环境中的BLA区神经元旷场放电信号(10 min),每日1次,连续7 d。利用Offline Sorter软件对采集到的神经元放电信号进行处理,过滤低频放电和环境干扰放电,设置信噪比>3:1的信号为所需的动作电位信号,运用主成分分析(principal component analysis,PCA)技术对提取出的动作电位进行分类处理,并计算每一类神经元动作电位的放电频率。将每只大鼠的每一类神经元动作电位在旷场中的放电频率减去其基础放电频率,得出的值定义为神经元动作电位放电频率增加值,以此值进行两组大鼠在旷场中神经元动作电位放电频率变化的对比。

2 结果
2.1炒酸枣仁对焦虑大鼠行为学动态变化的影响 随着测试天数的增加,大鼠跨格次数、站立次数及总运动时间逐渐下降,进入中央格次数变化平稳,中央格运动时间、中央格/总运动时间(%)逐渐上升。与模型对照组比较,炒酸枣仁组大鼠跨格次数在第5天明显减少(P<0.05);站立次数在第2,4,5,7天明显减少(P<0.05或P<0.01);总运动时间1~7 d均无明显变化(P>0.05);进入中央格次数在第1天明显增加(P<0.01);中央格运动时间在第1~第3,第6天明显增加(P<0.05或P<0.01);中央格/总运动时间(%)在第1,第3、第5~第7天明显增加(P<0.05或P<0.01)。见图1。

①与模型对照组比较,P<0.05;②与模型对照组比较,P<0.01。
2.2炒酸枣仁对焦虑大鼠BLA区神经元信息编码动态变化的影响
2.2.1大鼠BLA区神经元动作电位的分类及特征图 在大鼠BLA区记录到的神经元动作电位波形特征为:波长较长,波峰后曲线下降缓慢。按照放电特征的不同,把在BLA区记录到的神经元分为Ⅰ、Ⅱ两类(以下称BLA区Ⅰ类神经元为NeuronⅠ,Ⅱ类神经元为NeuronⅡ)。NeuronⅠ的时长较长、振幅稍低,自相关图中出现典型的尖峰、峰值后表现出快速的指数型下降;NeuronⅡ的时长较短、振幅较高,自相关图中峰值后下降缓慢。两类神经元动作电位放电模式的特征图见图2。

A.NeuronⅠ动作电位;B.NeuronⅡ动作电位;a.波形图;b.放电间隔直方图;c.自相关图。
2.2.2焦虑大鼠BLA区神经元动作电位放电频率的动态变化 与自身的基础放电频率相比,模型对照组和炒酸枣仁组大鼠在旷场中NeuronⅠ、NeuronⅡ动作电位放电频率在1~7 d均有所增加;NeuronⅠ、NeuronⅡ在旷场中动作电位放电频率在1~2 d均有所增加,然后在2~7 d逐渐下降。NeuronⅠ与NeuronⅡ的放电数比例约为2:1。各组放电频率直方图见图3。
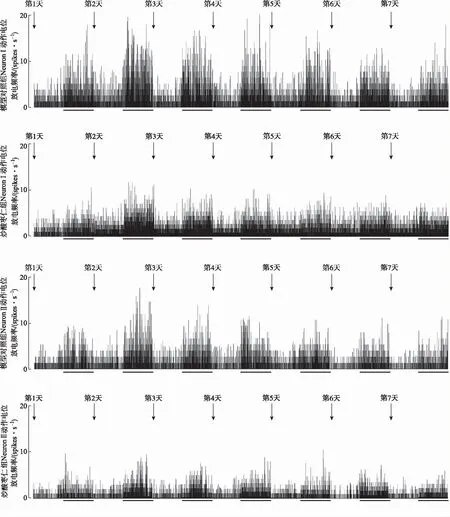
底部未画黑线部分为大鼠处于日常饲养环境中的基础放电,画黑线部分为大鼠处于旷场环境中的旷场放电。
2.2.3炒酸枣仁对焦虑大鼠BLA区神经元动作电位放电频率动态变化的影响 由于与自身的基础放电频率相比,每只大鼠在旷场中的NeuronⅠ、NeuronⅡ动作电位放电频率在1~7 d均有所增加,以神经元动作电位放电频率增加值进行两组大鼠在旷场中动作电位放电频率变化的对比。实验结果表明,与模型对照组比较,炒酸枣仁组大鼠BLA区NeuronⅠ的动作电位放电频率增加值在第1,3~6天明显减少(P<0.05或P<0.01),NeuronⅡ的动作电位放电频率增加值在第1,3,4,6,7天明显减少((P<0.05或P<0.01)。见图4。

①与模型对照组比较,P<0.05;②与模型对照组比较,P<0.01。
3 讨论
啮齿动物面对旷场提供的新环境时,同时产生对空旷环境的恐惧和对新奇环境的探索欲[14],旷场实验利用这种矛盾冲突心理,在造成动物焦虑情绪的同时,进行运动性和焦虑水平的评估[15-16]。本文利用旷场实验制备大鼠焦虑模型,考察了连续7 d大鼠焦虑状态下的行为学动态变化。旷场行为学结果显示,大鼠的运动性相关指标,包括跨格次数、站立次数及总运动时间,随着测试时间的增加均逐渐减少。这一现象反映,随实验时间的增加,大鼠对旷场环境逐渐适应从而减弱它们对旷场的好奇心,降低大鼠的探索行为。
旷场环境的中央格区域对动物意味着潜在威胁,周边格相较而言更安全,动物在中央格区域的探究行为越多,代表动物的焦虑程度越低[17]。连续7 d的行为学实验结果显示,大鼠的焦虑相关指标,包括中央格运动时间及中央格运动时间/总运动时间(%),随着测试时间的增加而逐渐增加,表明动物的焦虑程度逐渐减轻。大鼠的焦虑样行为在初次接触旷场时表现得更为强烈,而焦虑水平随着重复接触旷场而逐渐降低。这些观察结果证明,动物在反复暴露于旷场环境后表现出习惯化行为,与文献[18]报道一致。HSIAO等[19]报道,连续进行4 d的旷场测试,动物在中心格区域运动时间会逐渐减少,即反复接触旷场后焦虑水平会增加,与本文的实验结果有所差异,可能是因为每天动物进行50 min的旷场实验,而本文每日只测试10 min。因此,推测重复暴露于致焦虑环境中的时间过长不会建立习惯化行为,反而会造成焦虑程度增加。
本实验在利用旷场实验制备大鼠焦虑模型的同时,记录连续7 d大鼠BLA区在体神经元的基础放电及在旷场环境中旷场放电情况。本文采集到的BLA区神经元动作电位的波形特征为波长较长,波峰后曲线下降缓慢,与文献[20]报道结果一致。将在BLA区收集到的神经元信号按动作电位的波形和放电特征分为Ⅰ、Ⅱ两类,与基础放电频率相比,所有大鼠的NeuronⅠ、NeuronⅡ在连续7 d的旷场环境中其动作电位放电频率均增加。这一现象表明,旷场环境改变大鼠BLA区的神经元信息编码,NeuronⅠ、NeuronⅡ均参与了旷场对动物的焦虑加工。
通过对BLA区神经元动作电位放电频率增加值变化的观察,发现NeuronⅠ、NeuronⅡ的动作电位放电频率在1~2 d均有所增加,然后在2~7 d持续下降。文献[20]报道小鼠在连续3 d在旷场环境中BLA区神经元动作电位放电频率持续增加,本研究在1~2 d的结果与上述报道相似。这一现象提示,反复暴露于旷场环境中可能会引发BLA区神经元突触可塑性的短期增强,提高BLA区神经元的兴奋性。本研究结果显示从第2天开始BLA区神经元动作电位放电频率呈下降趋势,与WANG等[20]研究存在差异,推测造成此差异的原因与在旷场中的暴露时间有关。WANG等[20]研究中动物每日进行30 min的旷场实验,而本文每日只测试10 min,提示重复接触致焦虑的旷场环境的时间过长不会建立习惯化行为,反而会造成动物焦虑水平的逐渐增加,这与行为学结果的分析相同,但仍需进一步的研究确定。本文第2~7天的结果证明,由于测试次数的不断增加,大鼠建立习惯性行为,逐渐减轻在旷场中的焦虑程度,从而降低了BLA区神经元的放电频率,与行为学测试的结果一致。
本研究在观察焦虑大鼠行为学和神经元在体活动动态变化的同时,给予炒酸枣仁水煎液干预,探讨炒酸枣仁对其的影响作用。行为学结果显示,炒酸枣仁减少大鼠在第5,7天的跨格次数,并减少了第2,4,5,7天的站立次数。这一现象表明炒酸枣仁能降低重复暴露于旷场环境中大鼠的运动性和探索行为。同时,炒酸枣仁增加大鼠第1天的进入中央格次数,增加第1~3,6天的中央格运动时间,并增加了第1,3,5~7天的中央格/总运动时间,表明炒酸枣仁能减轻旷场对大鼠造成的焦虑情绪。由于重复暴露于旷场环境中,大鼠建立了习惯化行为,焦虑水平随着测试次数增加而降低,然而炒酸枣仁仍然能降低重复测试中动物的焦虑水平,提示炒酸枣仁的抗焦虑效果远远超过了动物自身的适应能力。
神经元在体电活动实验结果显示,炒酸枣仁在第1,3~6天减少了大鼠NeuronⅠ动作电位放电频率增加值,在第1,3,4,6,7天减少了大鼠NeuronⅡ动作电位放电频率增加值。表明炒酸枣仁通过抑制BLA区神经元的过度放电改变BLA区神经元信息编码,拮抗旷场对大鼠造成的焦虑状态。此外,在某些测试时间,炒酸枣仁能明显减少BLA区神经元电活动,但动物的焦虑样行为并没有显著变化。推测是由于炒酸枣仁的抗焦虑作用并不是由某一脑区的一类神经元介导,可能是多个脑区或多类神经元共同作用的结果,具体原因还需进一步研究。
