钱塘江桐庐渔业资源增殖放流区底栖动物群落结构特征分析
2016-08-18张爱菊刘金殿杨元杰郭爱环顾志敏浙江省淡水水产研究所浙江湖州313001农业部淡水渔业健康养殖重点实验室浙江湖州313001浙江省淡水水产遗传育种重点实验室浙江湖州313001
张爱菊,刘金殿,*,杨元杰,郭爱环,顾志敏,*(1.浙江省淡水水产研究所,浙江湖州313001;2.农业部淡水渔业健康养殖重点实验室,浙江湖州313001;3.浙江省淡水水产遗传育种重点实验室,浙江湖州313001)
钱塘江桐庐渔业资源增殖放流区底栖动物群落结构特征分析
张爱菊1,2,3,刘金殿1,2,3,*,杨元杰1,2,3,郭爱环1,2,3,顾志敏1,2,3,*
(1.浙江省淡水水产研究所,浙江湖州313001;2.农业部淡水渔业健康养殖重点实验室,浙江湖州313001;3.浙江省淡水水产遗传育种重点实验室,浙江湖州313001)
2013年2月—2013年12月,对浙江钱塘江桐庐段渔业资源增殖放流区的底栖动物按季度进行了调查,共采集到底栖动物16种,主要由软体动物、环节动物及节肢动物3大类组成,软体动物种类数占优势。相对重要性指数(IRI)分析结果显示,河蚬(Corbicula fluminea)、湖沼股蛤(Limnoperna lacustris)和齿吻沙蚕(Nephthys sp.)为主要优势种。底栖动物可归纳为4个摄食功能群——滤食收集者、牧食收集者、刮食者和捕食者,其中,滤食收集者的密度和生物量占绝对优势。底栖动物的年均密度和生物量分别为(393.75± 173.72)m-2和(295.72±131.97)g·m-2。方差分析显示,密度和生物量无显著性季节差异,但存在极显著的空间差异。底栖动物群落结构聚类分析表明,设置的6个断面4个季节均可分为A,B两个组群,A组群在春、夏、秋季均包含了断面Ⅰ、断面Ⅵ、断面Ⅴ和断面Ⅳ,B组群包含断面Ⅲ和断面Ⅱ,而冬季略有区别。
底栖动物;渔业增殖放流区;桐庐段;群落结构
底栖动物是淡水生态系统的重要组成部分,在物质循环和能量流动中起着重要的作用。底栖动物对环境污染的反应较灵敏,能较直观地反映水质的变化,是常用的水体状况指示生物,已广泛用于湖泊分类、水质的生物学评价和河流生态系统健康评价[1-4]。同时,底栖动物是鱼类等水生动物的天然优质饵料,对渔业生产有着重要作用[5]。因此,开展河流中底栖动物群落的研究,有助于更加全面地认识河流生态系统,进一步了解水生生物与河流系统的相互关系,对于河流系统的生态保护与生态恢复亦具有参考价值和指导意义[6]。
桐庐县位于浙江省西北部,地处钱塘江中游,介于东经119°10′—119°58′和北纬29°35′—30°05′之间。富春江自南而北贯穿桐庐全境,境内长33 km;分水江是富春江的主要支流之一,从西北部入桐庐境,境内长53 km。据统计,桐庐县大江宜渔面积4 333 hm2,而自2000年开始,大江水体呈污染趋势。研究认为,增殖放流活动具有生态修复的功能[7],因此,桐庐县政府在采取其他措施修复生态环境的同时,在2007年规划了渔业资源增殖放流区,并加大了在该区对花、黄尾密鲴、三角鲂、赤眼鳟、翘嘴红鲌等土著名特优种类的放流力度,至今放流累计2 000余万尾,生态环境得到了明显的改善。但是,目前关于该增殖放流区的水化学、水生生物生态学的研究还未见报道。于2013年对该区的底栖动物群落结构特征进行初步研究,并对部分常规水质理化指标进行检测,以期为水环境保护、水质与渔业管理提供一定的理论依据,并为该区今后底栖动物的后续演替研究提供参照。
1 研究区域与方法
1.1 研究区概况
浙江钱塘江桐庐段增殖放流区是浙江省海洋与渔业局于2007年12月批准设立的省级渔业资源增殖放流区,位于桐庐县富春江和分水江交汇处,具体为桐君街道、旧县街道的分水江段,富春江水库大坝下富春江段和分水江库区等,总面积约1 000 hm2。增殖放流区富春江段以径流作用为主,其径流控制站为富春江电站。江面宽400~1 000 m,多年平均流量为952 m3·s-1。富春江段径流量年际间变幅较大,具有明显的连续丰、枯水年交替变化。径流年内分配具有明显的枯洪之分,每年的3—7月径流量占全年的70%,流域内大洪水主要由梅雨造成。富春江段在秋季大潮期仍受潮汐的影响;分水江段属于分水江下游,宽浅,滩多流急,水位暴涨暴落,具有山溪性河流特点。上游建成分水江水库后,其径流控制站为分水江电站。分水江年最小流量一般出现在10月至翌年2月,多年月平均最小流量为10.6 m3·s-1,历年月平均最小流量仅1.8 m3· s-1[8]。
1.2 野外调查
于2013年5月至2013年12月的5月、7月、10月、12月的20日左右对增殖放流区采样1次。根据该区的地理特点设置6个采样断面(图1),以调查底栖动物群落特征。在地理位上,断面Ⅵ、断面Ⅴ和断面Ⅳ属于钱塘江支流分水江水域,而断面Ⅰ、断面Ⅱ和断面Ⅲ属于钱塘江干流富春江水域。采用改良Peterson采泥器(开口面积为1/16 m2),每断面设置3个采样点,其中2个采样点位于断面两端,另1个采样点位于断面中心,采样点连续采集2次,每断面采样点样品采集后混合,经40目过滤网筛洗后,将底栖生物挑出,保存在8%的福尔马林溶液中。样品带回室内后,在解剖镜、显微镜下进行种类鉴定,以分析天平称取质量[9]。同时,测量测定透明度(SD)、水温(WT)、表层水叶绿素a(Chla)、溶解氧(DO)、总氮(TN)、总磷(TP)、pH值等理化因子[10]。钱塘江桐庐段河床质中60%以上为砾、卵石(d50=2~30 mm),其余为中粗砂(d50=0.25~2 mm),滩地主要是粉砂和黏土。但悬沙中大部分由0.25 mm以下的细砂、粉砂和黏土组成,大于0.25 mm的中粗砂含量甚微[8,11]。
1.3 初级生产力估算
初级生产力(Ps)根据表层水叶绿素a的含量估算,估算公式为:

式(1)中:Ps为表层水(1 m以内)中浮游植物的初级生产力,mg C·m-3·h-1;Ca为表层叶绿素a的含量,mg·m-3;Q为表层水同化系数,为3.7,mg C·mg-1·h-1。
1.4 相对重要性指数计算
由于不同种类底栖动物密度和生物量差异较大,参照韩洁等[12]和吴召仕等[13]的方法,采用相对重要性指数(IRI)来确定各采样断面中的优势种,其计算公式为:

式(2)中:W为某一种类的生物量占总生物量的百分比;N为该种类的密度占总密度的百分比;F为该物种在各断面出现的相对频率。
1.5 群落结构分析
为了减小数据间的差异,先对密度数据进行二次方根转换(transformation:second root),然后计算Bray-Curtis相似性系数,以Bray-Curtis相似性系数为基础构建相似性矩阵,进一步采用类平均法对站位等级进行CLUSTER聚类和非参数多变量排序(MDS),分析底栖动物群落结构特征。采用Two-Way Crossed ANOSIM分析来检验不同断面不同季节间群落结构的差异,并使用SIMPER分析来了解表征群落的物种和造成群落差异的物种[14]。CLUSTER、MDS和SIMPER等分析方法都在PRIMER 5.0软件上进行计算。

图1 采样断面的分布Fig.1 Sampling section distribution in the enhancement and releasing zone
2 结果与分析
2.1 种类组成与优势种
调查共发现底栖动物16种,主要由软体动物、环节动物及节肢动物3大类组成。软体动物9种,占总种数的56.25%,其中,腹足类5种(31.25%),双壳类4种(25.00%);环节动物4种(25.00%),其中,多毛类和寡毛类各2种,分别占总种数的12.50%;节肢动物摇蚊科幼虫3种,占总种数的18.75% 。
相对重要性指数计算发现,河蚬(Corbicula fluminea)的IRI值最高,为136.51,且与其他物种的IRI值相差较大,可见河蚬在该增殖放流区处于最优势地位;湖沼股蛤(Limnoperna lacustris)的IRI值(31.03)仅次于河蚬,为第二优势种;齿吻沙蚕(Nephthys sp.)的 IRI值(4.07)位列第三(表1)。
2.2 现存量
调查发现,增殖放流区底栖动物的年均密度和生物量分别为(393.75±173.72)m-2和(295.72±131.97)g·m-2。从密度来看,增殖放流区双壳类>多毛类>腹足类>水生昆虫类(摇蚊科幼虫)>寡毛类;从生物量来看,双壳类>腹足类>多毛类>寡毛类>水生昆虫类(摇蚊科幼虫)。由此可见,双壳类在密度和生物量上均占绝对优势,所占比例远高于其他类群(表2)。
在季节变化上,调查发现,底栖动物密度呈现夏季>冬季>秋季>春季的趋势(图2),但无显著性季节差异(P>0.05);在空间变化上,方差分析显示底栖动物密度存在极显著差异(P<0.01)。从分水江水域断面来看,断面Ⅵ年均密度最大,断面Ⅴ次之,断面Ⅳ最小;从富春江水域断面来看,断面Ⅲ年均密度最大,其次为断面Ⅱ,断面Ⅰ最小(图3)。多重比较显示,断面Ⅳ、断面Ⅴ和断面Ⅵ与断面Ⅰ之间无显著性密度差异(P >0.05),断面Ⅱ与断面Ⅲ之间无显著性密度差异(P>0.05)。
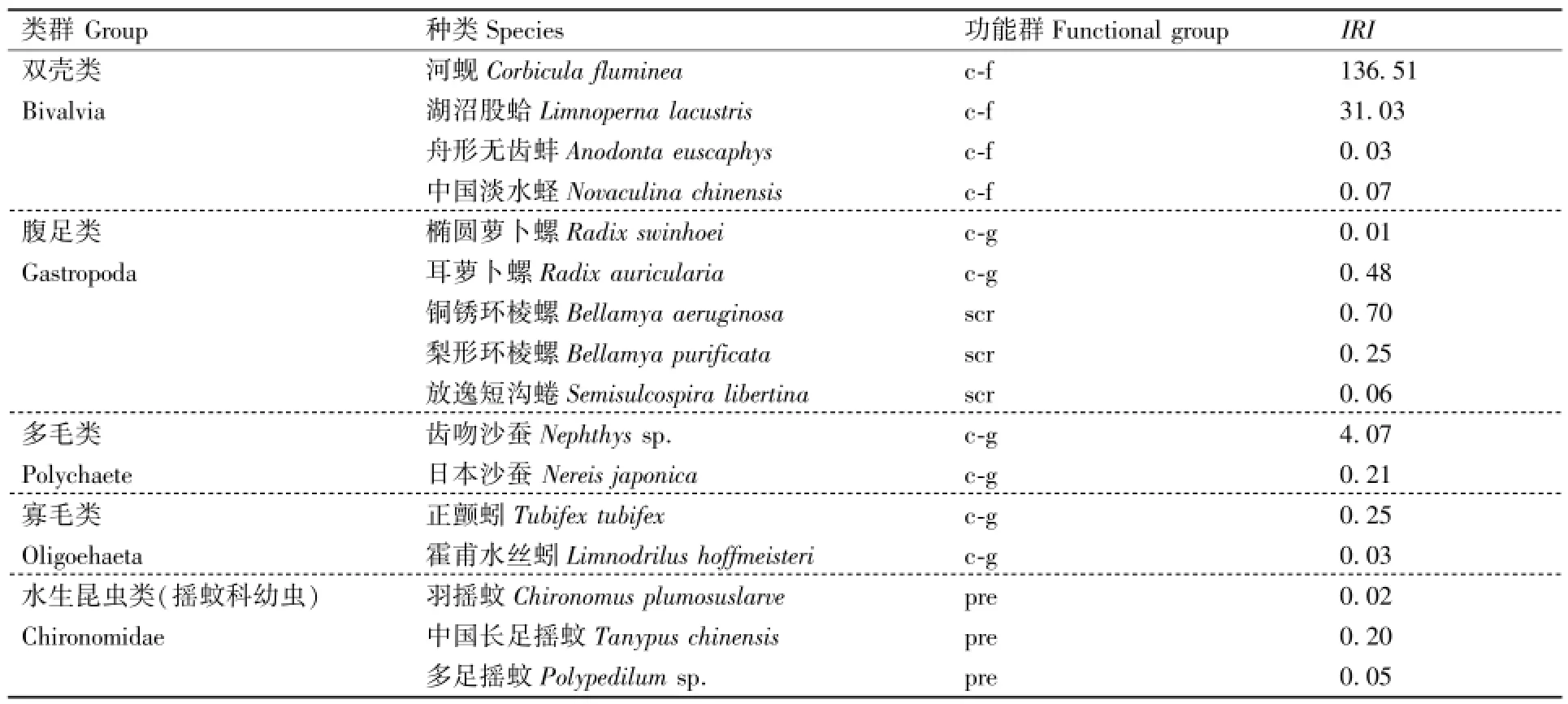
表1 底栖动物的IRITable 1 IRI of the macrozoobenthic species in the enhancement and releasing zone

表2 五种常见类群底栖动物平均密度和平均生物量Table 2 Average density and biomass of the five common macrozoobenthic groups in the sampling sections
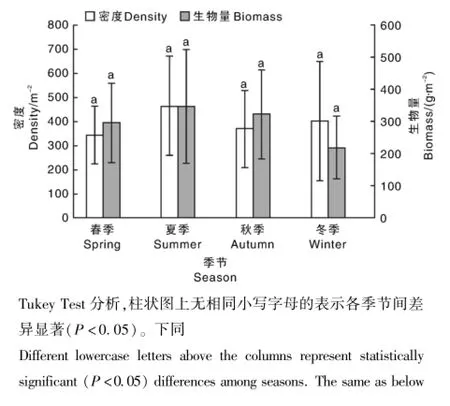
图2 底栖动物密度和生物量的季节变化Fig.2 Seasonal variations of macrozoobenthic density and biomass in the sampling sections

图3 底栖动物密度和生物量的空间分布Fig.3 Spatial distributions of macrozoobenthic density and biomass in the sampling sections
2.3 功能群的划分
底栖动物功能群是具有相同生态功能的底栖动物的组合[15]。根据摄食对象和方式的差异,所鉴别的底栖动物可归纳为4个摄食功能群:滤食收集者(collector-filterer)、牧食收集者(collector-gatherer)、刮食者(scraper)和捕食者(predator)(表1)。牧食收集者物种数最多,为6种,占总种数的37.50%,其次为滤食收集者(4种,25.00%),刮食者和捕食者物种数较少。滤食收集者的密度和生物量占绝对优势,比例分别达到88.63%和94.85%。密度和生物量占第二优势的功能群有所区别,分别为牧食收集者(8.89%)和刮食者(4.60%),捕食者的密度和生物量均为最低(表3)。
2.4 群落结构特征分析
2.4.1 空间变化
根据各个断面底栖动物组成情况,进行群落结构聚类分析和MDS排序分析(表4)。聚类分析表明,4个季节都可分为A、B两个组群,且4个季节的MDS排序分析的胁强系数(Stress)都小于等于0.01。
春季,A类组包含了4个断面(断面Ⅴ、Ⅳ、Ⅵ和Ⅰ),B类组包含了2个断面(断面Ⅱ和Ⅲ)。SIMPER分析的结果显示,A类组的组内平均相似性是65.34%,共有3种对组内平均相似性贡献率之和超过90.0%的底栖动物,分别为河蚬、齿吻沙蚕、正颤蚓(Tubifex tubifex)。B类组的组内平均相似性是65.40%,共有2种对组内平均相似性贡献率之和超过90.0%的底栖动物,分别为河蚬和湖沼股蛤。A和B两个类组之间的平均相异性为61.70%,共有6种底栖动物对A和B组间相异性的累积贡献率达92.82%,分别为河蚬、湖沼股蛤、齿吻沙蚕、正颤蚓、霍甫水丝蚓(Limnodrilus hoffmeisteri)和羽摇蚊(Chironomus plumosuslarve)。
夏季,A类组包含了4个断面(断面Ⅴ、Ⅳ、Ⅵ和Ⅰ),B类组包含了2个断面(断面Ⅱ和Ⅲ)。SIMPER分析的结果显示,A类组的组内平均相似性是68.64%,河蚬和齿吻沙蚕对组内平均相似性贡献率之和超过90.0%。B类组的组内平均相似性是71.91%,河蚬和湖沼股蛤对组内平均相似性贡献率之和超过90.0%。A、B两个类组之间的平均相异性为68.82%,共有6种底栖动物对A和B组间相异性的累积贡献率达91.67%,分别为湖沼股蛤、河蚬、铜锈环棱螺(Bellamya aeruginosa)、齿吻沙蚕、梨形环棱螺(Bellamya purificata)、放逸短沟蜷(Semisulcospira libertina)。

表3 底栖动物的功能摄食群Table 3 Composition of feeding functional group in the sampling sections
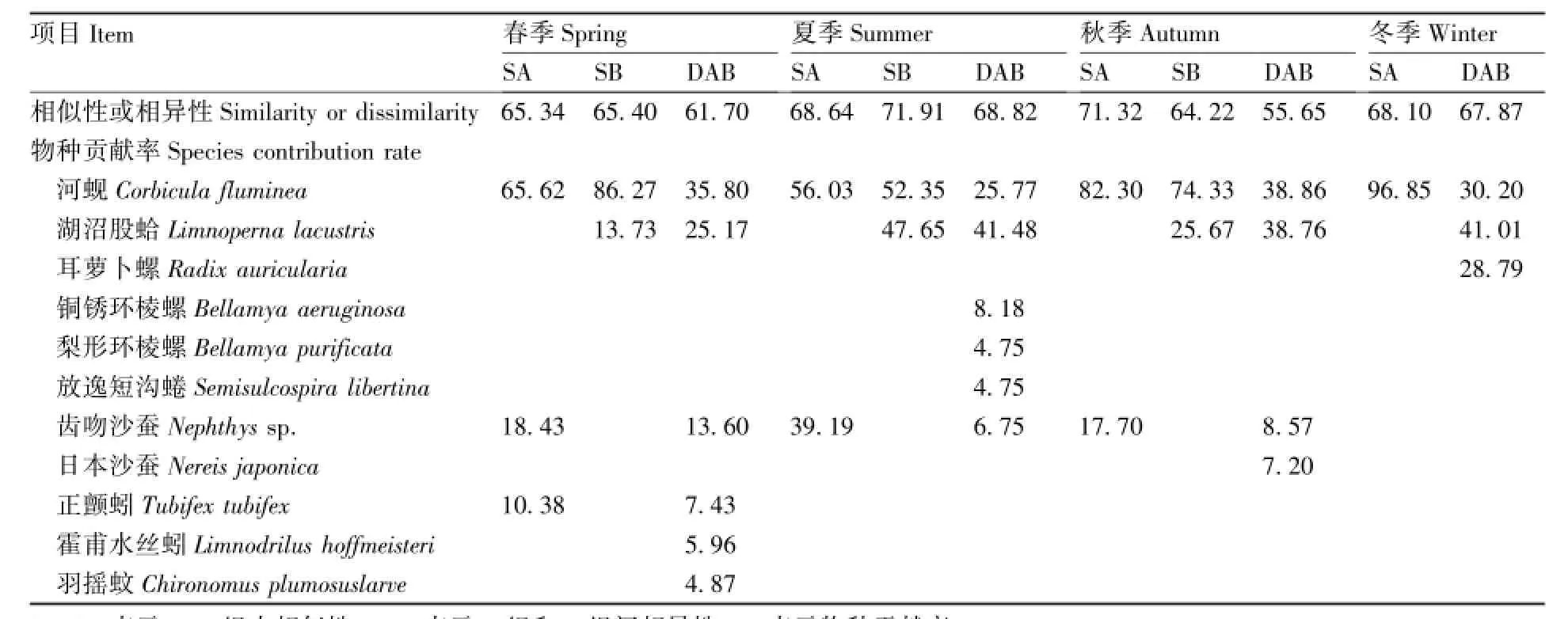
表4 渔业资源增殖放流区水域各季节对A、B组内相似性和组间相异性贡献率较大的底栖动物Table 4 The indicator species yielding more contribution for the similarity and dissimilarity in the waters of enhancement and releasing zone %
秋季,A类组包含了断面Ⅴ、Ⅳ、Ⅵ和Ⅰ等4个断面,B类组包含了断面Ⅱ和Ⅲ 2个断面。SIMPER分析的结果显示,A类组的组内平均相似性是71.32%,河蚬和齿吻沙蚕2种底栖动物对组内平均相似性贡献率之和超过90.00%。B类组的组内平均相似性是64.22%,河蚬和湖沼股蛤对组内平均相似性贡献率之和超过90.00%。A、B两个类组之间的平均相异性为55.65%,共有4种底栖动物对A和B组间相异性的累积贡献率达93.39%,分别为河蚬、湖沼股蛤、齿吻沙蚕和日本沙蚕(Nereis japonica)。
冬季,A类组包含了5个断面(断面Ⅴ、Ⅳ、Ⅵ、Ⅰ和Ⅱ),B类组包含了1个断面(断面Ⅲ)。SIMPER分析的结果显示,A类组的组内平均相似性是68.10%,河蚬对组内平均相似性贡献率超过90.00%。A、B类组之间的平均相异性为67.87%,共有3种底栖动物对A和B组间相异性的累积贡献率达100.00%,分别为湖沼股蛤、河蚬、耳萝卜螺(Radix auricularia)。

表5 渔业资源增殖放流区水域各季节底栖动物的典型种及贡献率Table 5 Representative species for different seasons and their contribution rates in the waters of enhancement and releasing zone %
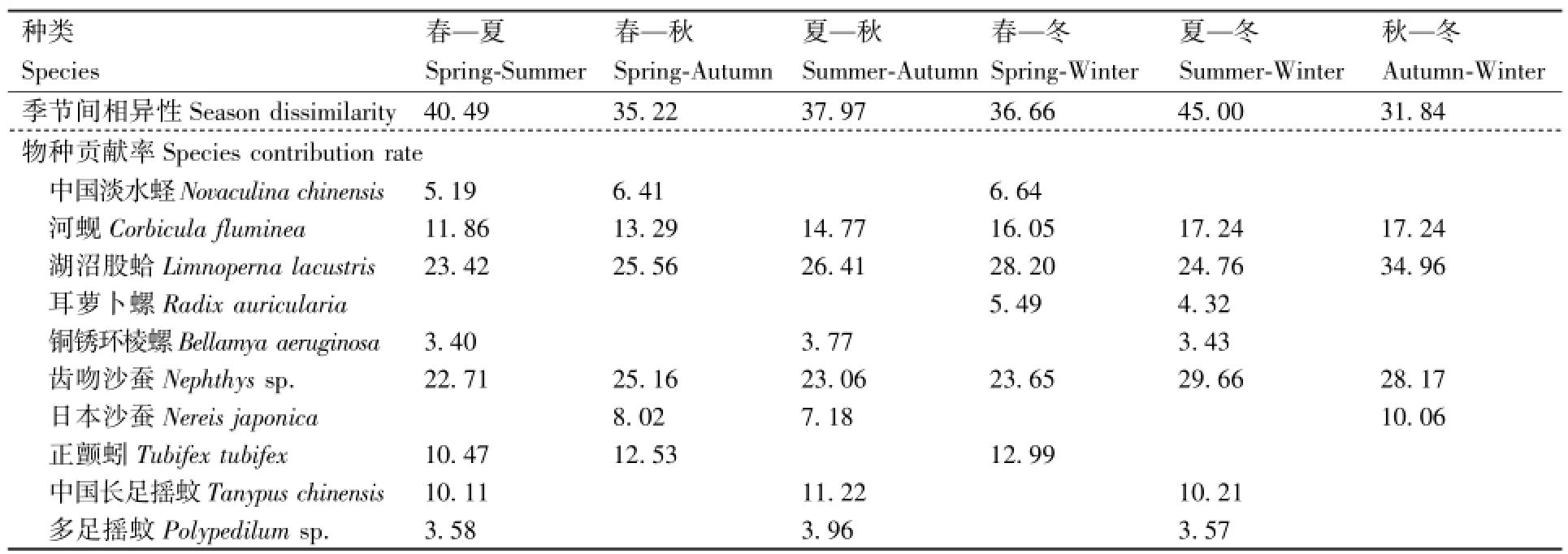
表6 渔业资源增殖放流区水域各季节间的分歧种及对季节间平均相异性的贡献率Table 6 Discriminating species and their contribution rates to the average seasonal dissimilarity in waters of the enhancement and releasing zone %
2.4.2 季节变化
对4个季节的底栖动物组成进行SIMPER分析表明,4个季节的组间相似性分别为60.65%、57.61%、68.58%和74.15%(表5)。将增殖放流区水域4个季节内对组间相似性贡献率累积超过90.00%的底栖动物种类及其贡献百分比列于表5,其中,春季主要典型种包括河蚬、齿吻沙蚕、湖沼股蛤,对春季的平均相似性贡献累积率达到94.07%;夏季主要典型种与春季相似,包括河蚬、齿吻沙蚕、湖沼股蛤,对夏季的平均相似性贡献累积率达到98.04%;秋季主要典型种包括河蚬、齿吻沙蚕,对秋季的平均相似性贡献累积率达到92.52%;冬季主要典型种为河蚬,对冬季的平均相似性贡献率达到93.75%。
表6列出影响各季节间底栖动物群落结构差异的主要种类及其贡献率。夏、冬季间的平均相异性指数最高,达到45.00%,春、夏季间次之,为40.49%,秋、冬间相异性最低,为31.84%。对4个季节相互间底栖动物群落结构相异性贡献率较大的为湖沼股蛤、齿吻沙蚕、河蚬等。
2.5 水体理化因子参数及初级生产力估算
7个理化因子的季度平均值列于表7。单因子分析显示,7个理化因子在分水江和富春江断面间均没有显著差异。值得一提的是,表层水初级生产力在Ⅲ号断面最高,为17.645 mg C·m-3· h-1,其次为Ⅴ号断面,为10.845 mg C·m-3·h-1,其余断面均不超过10 mg C·m-3·h-1。

表7 各采样断面理化因子和初级生产力的年平均值Table 7 Annual average values of physical and chemical parameters and primary production in sampling sections
3 讨论
影响河流中底栖动物群落的环境因子包括物理条件、水化学条件和生物条件等,非常复杂[16-17]。与水库、湖泊等不同,底质和流速可能是影响底栖动物分布的重要环境理化因子。研究表明,底栖动物群落的分布和物种组成在很大程度上取决于底质类型,底质是影响河流底栖动物群落结构最重要的物理因素之一。流速也是影响河流特征和结构的一个重要物理量,流速降低在一定程度上可加快有机碎屑沉积量的增加,而有机碎屑是底栖动物很重要的食物来源,当流速大于1.2 m·s-1时,流速会成为河流底栖动物的限制因子[6]。本次研究中,渔业资源增殖放流区位于河口区,属周期性往复水流,同时两江的上游分建有富春江水库和分水江水库,其泄洪会引起水流、水位、水温等条件的显著变化,且造成水库大坝下游河床底质的极度不稳定,加之该区内的挖沙行为[11]破坏了底质的生态平衡,干扰了底栖动物的生存环境,使许多底栖动物丧失原有的栖息地,降低了其多样性。在所采集的全部样品中,仅发现了16种底栖动物,其中,软体动物在物种数、密度和生物量上占绝对优势。分析发现,本次调查的总物种数(16种)低于分水江水库的37种[18]和富春江水库的20种[19],在优势种和物种多度上也有所差异,分水江水库主要由寡毛纲和摇蚊科物种组成,而富春江水库底栖动物数量上寡毛类占优势。
从时间上看,本次调查底栖动物的密度季节变化总体呈现出夏季>冬季>秋季>春季的趋势,但无显著性季节差异。然而,方差分析显示,底栖动物密度有极显著的空间变化。多重比较显示,断面Ⅳ、断面Ⅴ和断面Ⅵ与断面Ⅰ之间无显著性密度差异(P>0.05),断面Ⅱ与Ⅲ之间无显著性密度差异(P>0.05)。该结论与6个断面底栖动物组成聚类分析结果基本一致。聚类分析结果表明,4个季节中调查断面的组成大致均可以划分为2个类组。A类组在春、夏、秋季均包含断面Ⅰ、断面Ⅵ、断面Ⅴ、断面Ⅳ,B类组包含断面Ⅲ和Ⅱ,而冬季A,B类组断面组成略有区别。结合调查水域实际地理位置,B类组仅包括富春江水域断面,而A类组包含全部分水江水域断面及部分富春江水域断面,且主要为断面Ⅰ。从分水江水域断面来看,断面Ⅵ年均密度最大,断面Ⅴ次之,断面Ⅳ最小;从富春江水域断面来看,断面Ⅲ年均密度最大,其次为断面Ⅱ,断面Ⅰ最小,故均呈现出从上游到下游逐渐下降的趋势,该结论与马徐发等[20]、胡忠军等[21]的研究结果基本一致。分析认为,断面Ⅲ处于富春江上游,相对中下游断面而言,断面Ⅲ水深相对较浅,外源营养盐和有机物来源丰富,表层水初级生产力最高,为17.645 mg C·m-3·h-1,因而有机质沉降量可能较大,底质有机质含量可能较多,底栖动物密度和生物量亦较高。富春江段水体从断面Ⅲ流经中游的断面Ⅱ,与分水江段的水体(上游到下游依次为断面Ⅵ、断面Ⅴ、断面Ⅳ)汇合到达下游的断面Ⅰ,反映在表层水初级生产力上则呈现逐渐下降的趋势,即断面Ⅲ>断面Ⅱ>断面Ⅰ,同时水深的增加可能导致有机质沉降量下降,故底栖动物密度呈现逐渐降低的趋势。调查时也发现,富春江水域的断面Ⅰ和断面Ⅲ附近均有码头,断面Ⅱ位于游客中心上游,分水江水域断面Ⅴ和断面Ⅳ之间有码头,这些因素的存在从一定程度上增加了水体的营养水平,保证了水体中的有机质含量,维持了较高的底栖动物密度和生物量。值得一提的是,本次调查仅是对该增殖放流区底栖动物群落的一次初步研究,希望今后能从多空间和时间尺度上加强对该区渔业生态的研究,为今后渔业增殖放流提供切实可行的指导意见,从而更好地保护钱塘江流域的生态系统。
(References):
[1] 王备新.大型底栖无脊椎动物水质生物评价研究[D].南京:南京农业大学,2003. WANG B X.Water quality bioassessment using benthic macroinvertebrates[D].Nanjing:Nanjing Agricultural University,2003.(in Chinese with English abstract)
[2] 胡忠军,刘其根,陈立婧,等.上海崇明明珠湖摇蚊幼虫群落结构特征及其对水质的指示作用[J].应用生态学报,2009,20(4):929-936. HU Z J,LIU Q G,CHEN L J,et al.Structural characteristics of chironomid community and their indicative significance in bioassessment of water quality in Mingzhu Lake of Chongming Island,Shanghai[J].Chinese Journal of Applied Ecology,2009,20(4):929-936.(in Chinese with English abstract)
[3] 王琴,王海军,崔永德.武汉东湖水网区底栖动物群落特征及其水质的生物学评价[J].水生生物学报,2010,34 (4):739-746. WANG Q,WANG H J,CUI Y D.Community characteristics of the macrozoobenthos and bioassessment of water quality in Lake Donghu district,Wuhan[J].Acta Hydrobiologica Sinica,2010,34(4):739-746.(in Chinese with English abstract)
[4] 蔡永久,刘劲松,戴小琳,等.长荡湖大型底栖动物群落结构及水质生物学评价[J].生态学杂志,2014,33(5):1224-1232. CAI Y J,LIU J S,DAI X L,et al.Community structure of macrozoobenthos and bioassessment of water quality in Lake Changdang,Jiangsu Province[J].Chinese Journal of Ecology,2014,33(5):1224-1232.(in Chinese with English abstract)
[5] KAJAK Z,RYBAK J I,SPODNIEWSKA I,et al.Influence of the planktonivorous fish Hypophthalmichthys molitrix(Val.)on the plankton and benthos of the eutrophic lake[J].Polskie Archiwum Hydrobiologii,1975,22(22):301-310.
[6] 段学花,王兆印,徐梦珍.底栖动物与河流生态评价[M].北京:清华大学出版社,2010.
[7] 谢平.鲢,鳙与藻类水华控制[M].北京:科学出版社,2003:48-51.
[8] 曹新辉.分水江流域水文特性分析[J].黑龙江水专学报,2007,34(4):16-19. CAO X H.Analysis of hydrographical characteristics of Fenshui River[J].Journal of Heilongjiang Hydraulic Engineering College,2007,34(4):16-19.(in Chinese with English abstract)
[9] 金相灿,屠清瑛.湖泊富营养化调查规范[M].2版.北京:中国环境科学出版社,1990:255-257.
[10] 国家环境保护总局.水和废水监测分析方法[M].4版增补版.北京:中国环境科学出版社,2002:88-284.
[11] 陈来华.大规模采砂对富春江桐庐段河床影响及对策[J].人民长江,2006,37(10):70-73. CHEN L H.Impact of large-scale sand mining on the river bed of Tonglu reach of Fuchunjiang River and countermeasures[J].Yangtze River,2006,37(10):70-73.(in Chinese)
[12] 韩洁,张志南,于子山.渤海中、南部大型底栖动物的群落结构[J].生态学报,2004,24(3):531-537. HAN J,ZHANG Z N,YU Z S.Macrobethic community structure in the southern and central Bohai Sea,China[J].Acta Ecologica Sinica,2004,24(3):531-537.(in Chinese with English abstract)
[13] 吴召仕,蔡永久,陈宇炜,等.太湖流域主要河流大型底栖动物群落结构及水质生物学评价[J].湖泊科学,2011,23(5):686-694. WU Z S,CAI Y J,CHEN Y W,et al.Assemblage structure investigation of macrozoobenthos and water quality bioassessment of the main river systems in Taihu Basin[J].Journal of Lake Sciences,2011,23(5):686-694.(in Chinese with English abstract)
[14] BRAY T R,CURTIS J T.An ordination of the upland forest communities of southern Wisconsin[J].Ecological Monographs,1957,27:325-349.
[15] FAUCHALD K,JUMARS P A.The diet of worms:a study of polychaete feeding guilds[J].Oceanography and Marine Biology-Annual Review,1979,17:193-284.
[16] 王丽卿,吴亮,张瑞雷,等.滆湖底栖动物群落的时空变化及水质生物学评价[J].生态学杂志,2012,31(8):1990-1996. WANG L Q,WU L,ZHANG R L,et al.Spatiotemporal variation of zoobenthos community and bio-assessment of water quality in Gehu Lake[J].Chinese Journal of Ecology,2012,31(8):1990-1996.(in Chinese with English abstract)
[17] CAI Y J,GONG Z J,QIN B Q.Benthic macroinvertebrate community structure in Lake Taihu,China:Effects of trophic status,wind-induced disturbance and habitat complexity [J].Journal of Great Lakes Research,2012,38(1):39 -48.
[18] 刘其根,查玉婷,陈立侨,等.浙江分水江水库大型底栖动物群落结构及水质评价[J].应用生态学报,2012,23 (5):1377-1384.LIU Q G,ZHA Y T,CHEN L Q,et al.Macrozoobenthos community structure and its indicative significance in water quality bio-assessment of Fenshuijiang Reservoir,Zhejiang Province of East China[J].Chinese Journal of Applied Ecology,2012,23(5):1377-1384.(in Chinese with English abstract)
[19] 韩轶才,盛海燕,吴洁,等.富春江水库大型底栖动物群落的初步研究[J].环境污染与防治,2011,33(12):23 -26,32. HAN Y C,SHENG H Y,WU J,et al.Preliminary study on macrobenthos community of a large riverine reservoir,Fuchunjiang Reservoir in subtropical China[J].Environmental Pollution&Control,2011,33(12):23-26,32.(in Chinese with English abstract)
[20] 马徐发,熊帮喜,王明学,等.湖北道观湖水库大型底栖动物的群落结构及物种多样性[J].湖泊科学,2004,16(1):49-55. MA X F,XIONG B X,WANG M X,et al.The community structure and biodiversity of macrozoobenthos in Daoguanhe Reservoir,Hubei Province[J].Journal of Lake Sciences,2004,16(1):49-55.(in Chinese with English abstract)
[21] 胡忠军,孙月娟,刘其根,等.浙江千岛湖深水区大型底栖动物时空变化格局[J].湖泊科学,2010,22(2):265 -271. HU Z J,SUN Y J,LIU Q G,et al.Temporal and spatial distribution of profundal macrozoobenthic community in Lake Qiandao,Zhejiang,China[J].Journal of Lake Sciences,2010,22(2):265-271.(in Chinese with English abstract)
(责任编辑 卢福庄)
Analysis of community characteristics of macrozoobenthos in enhancement and releasing zone in Tonglu section of Qiantang River
ZHANG Ai-ju1,2,3,LIU Jin-dian1,2,3,*,YANG Yuan-jie1,2,3,GUO Ai-huan1,2,3,GU Zhi-min1,2,3,*
(1.Zhejiang Institute of Freshwater Fisheries,Huzhou 313001,China;2.Key Laboratory of Healthy Freshwater Aquaculture,Ministry of Agriculture,Huzhou 313001,China;3.Key Laboratory of Freshwater Aquatic Animal Genetic and Breeding of Zhejiang Province,Huzhou 313001,China)
A seasonal investigation was conducted on the macrozoobenthos at 6 sections of enhancement and releasing zone in Tonglu section of Qiantang River from February to December in 2013.A total of 16 species were collected,which belonged to Mollusca,Annelida and Arthropoda,and Mollusca had the largest number of species.According to IRI value,Corbicula fluminea,Limnoperna lacustris and Nephthys sp.were the dominant species.All species were classified to 4 functional feeding groups:collector-filterer,collector-gatherer,scraper and predator,of which the collector-filterer was the predominant group.The mean annual density and biomass of the community were(393.75± 173.72)m-2and(295.72±131.97)g·m-2,respectively.There was no temporal significant difference in the macrozoobenthic density and biomass,but there was a significant variation in space.According to the community struc-ture of macrozoobenthos,the sampling sections could be divided into 2 clusters,cluster A and B.In spring,summer and autumn,cluster A contained sectionⅠ,sectionⅥ,sectionⅤand sectionⅣ,while cluster B contained section Ⅲand sectionⅡ,which had a little difference in winter.
macrozoobenthos;enhancement and releasing zone;Tonglu section;community structure
S932
A
1004-1524(2016)08-1323-09
10.3969/j.issn.1004-1524.2016.08.08
2016-01-07
浙江省科技厅公共科技服务项目(2012F30023)
张爱菊(1981—),女,浙江湖州人,博士研究生,助理研究员,研究方向为水生生物与水污染生态修复。E-mail:zhangaiju2013 @163.com
*
,顾志敏,E-mail:guzhimin2006@163.com;刘金殿,E-mail:jdliu2008@hotmail.com
