活化TLR9的牛种布氏菌基因外重复回文序列筛选及活性检测
2016-07-28张雅娴白丽云王占黎
张雅娴,白丽云,王占黎,王 英,于 慧
活化TLR9的牛种布氏菌基因外重复回文序列筛选及活性检测
张雅娴1.4,白丽云2,王占黎2,王英2,于慧3
1.包头医学院,包头014040;2.包头医学院第一附属医院, 包头014040;3.包头医学院第二附属医院, 包头014040;4.赤峰市疾控中心, 赤峰024000
摘要:目的筛选具有活化Toll样受体9(TLR9)的牛种布氏菌基因外重复回文序列(Repetitive Extragenic Palindrome Squence, REPs)并检测其活性,为布氏菌病的治疗提供新思路。方法基于Brucella abortus A13334基因组序列,利用生物信息学技术识别其REPs后,合成序列。将合成的天然骨架的脱氧寡核苷酸(ODNs)转染小鼠单核巨噬细胞株RAW264.7,以ELISA检测IFN-α的分泌水平。采用TLR9-siRNA沉默RAW264.7中的TLR9,将上述诱导IFN-α分泌增加的阳性ODN序列转染RAW264.7,ELISA方法检测IFN-α的分泌变化。结果筛选出1 857条牛种布氏菌REPs,选择2级茎环结构较好的5条ODNs序列进行合成,ELISA方法检测显示ODNs M4、M5介导IFN-α分泌量显著高于阴性对照(P<0.05),且阳性ODN M5所介导的IFN-α分泌可以被TLR9-siRNA显著抑制。结论布氏菌基因组中存在可以活化TLR9信号通路的REPs,此结果有助于对布氏菌致病和免疫机制的认识。
关键词:牛种布氏菌;基因外重复回文序列;巨噬细胞;Toll样受体9;α-干扰素
布氏菌病(简称布病)是由布氏菌(Brucella)引起的人畜共患传染病,在全世界都有流行,尤其是在没有适当预防和控制措施的发展中国家和地区,包括拉丁美洲、亚洲、非洲、地中海地区等,每年新增病例 50 余万人[1]。近年来,我国布病疫情持续上升。据卫生部网站统计,2011 年共报告人间布病发病42 654例,发病率为3.18/10万,与2010年比较上升21.16%[2]。2013年我国布病发病人数46 089人,为历年新高,2014年发病率达3.21/10万,布病的防控与治疗工作刻不容缓。
布氏菌进入机体后,感染的靶细胞主要是巨噬细胞等免疫细胞[3]。免疫细胞内的Toll样受体 (Toll-like receptors, TLRs)对布氏菌发挥重要免疫防御作用[4]。TLRs是免疫细胞表面的模式识别受体, 参与病原相关分子模式 (pathogen-associated molecular patterns, PAMPs) 的识别,从而诱导机体免疫防御反应。最新研究发现,与其他TLRs成员相比,TLR9更能有效清除感染动物体内的布氏菌。Oliveira等研究发现,与野生型小鼠相比,TLR9基因敲除小鼠更易感染布氏菌[5]。TLR9主要通过胞内髓样分化因子88(Myd88)招募一系列信号分子,活化核因子NF-κB, 进而引起IFN-α等炎性因子级联反应,导致机体炎症反应。最近研究发现,大肠杆菌、大肠埃希氏菌、沙门氏菌、假单胞菌属假单胞菌和脑膜炎奈瑟氏球菌等革兰性阴性菌来源的基因外重复回文序列(Repetitive Extragenic Palindrome Squence, REPs)可经TLR9活化机体免疫系统,从而产生大量IFN-α[6]。此外,参与REPs刺激TLR9产生免疫作用在用B6.129P2 - TLR9tm1Aki基因敲除的小鼠身上中也得到了证实。在大肠埃希氏菌,沙门氏菌,假单胞菌属假单胞菌,和脑膜炎奈瑟氏球菌中,甲基化的REPs或删除CpG基序后无刺激作用进一步证实了对于天然细菌DNA的免疫刺激作用是依赖在未甲基化CpG基序。而REPs是包含CpG基序的DNA序列。与其他革兰氏阴性菌相似,布氏菌基因组中也存在REPs。但是,布氏菌DNA中REPs是否引起TLR9介导的免疫活化效应,目前尚不清楚。本研究旨在寻找牛种布氏菌 (Brucella abortus)DNA中能够活化TLR9的REPs,研究结果对于探讨布氏菌感染引起的免疫失衡具有重要意义。
1材料与方法
1.1主要试剂高糖DMEM培养基、胎牛血清(FBS)及减血清培养液(Opti-MEM) 购自美国Gibico公司,青霉素、链霉素购自美国Sigma公司,天然骨架脱氧寡核苷酸(ODNs)由上海生工生物工程股份有限公司合成,Lipofectamine 3000由Invitrogen公司提供,ELISA试剂盒购自美国Affymetrix公司。
1.2主要仪器二氧化碳细胞培养箱购自美国Thermo公司,倒置相差显微镜购自日本Nikon公司,移液器均购自德国Eppendorf公司,电子天平购自德国Sartorius公司,96孔细胞培养板购自Corning 公司。
1.3牛种布氏菌REPs的筛选以国内外公开报道的BrucellaabortusA13334基因组信息为研究对象(chromosome 1: NC_016795.1, GI:363399402. chromosome 2: NC_016777.1, GI: 363401588)。采用EMBOSS Cpgplot软件定位CpG岛,利用自主研发的C++程序进一步搜索REPs,采用软件Primer premier 5.0分析上述REPs的回文结构。
1.4天然骨架ODNs的合成针对上述获得的牛种布氏菌DNA的REPs,采用DNA合成仪合成天然磷酸二酯骨架ODNs来替代天然REPs,经HPLC纯化后,260 nm紫外定量后真空干燥,-20 ℃保存备用(上海生工生物工程股份有限公司)。
1.5细胞培养和脂质体转染参照文献[7]方法培养小鼠巨噬细胞RAW264.7,按1.0×104个/孔接种于96孔板中。至细胞生长状态良好,密度约为80%时,采用脂质体Lipofectamine 3000 (Opti-MEM配制) 转染ODNs,每个样本设3个复孔。在37 ℃、5%CO2培养箱中培养6 h后,更换为10%FBS高糖DMEM培养基继续培养,48 h后收集上清。
1.6ELISA检测IFN-α水平收集上述96孔板细胞培养上清,空白孔加入等体积的样品稀释液,按产品说明书步骤操作,酶标仪检测IFN-α的浓度。
1.7TLR9-siRNA转染小鼠TLR9-siRNA序列为5′-CCA ACA UCC UGG UUC UAG AUG CUA A-3′(购自Invitrogen公司)。按照上述方法将细胞RAW264.7接种于96孔板中,至细胞生长状态良好,密度约为80%时,采用脂质体Lipofectamine 3000转染TLR9-siRNA,并以阴性siRNA作为对照。在37 ℃、5%CO2培养箱中培养6 h后,更换为10%FBS高糖DMEM培养基继续培养24 h,按照上述方法转染ODNs,ELISA检测IFN-α水平。
1.8统计学分析采用SPSS 17.0对统计数据进行分析,两组数据的比较采用t检验,P<0.05有统计学意义。
2结果
2.1牛种布氏菌基因组DNA中REPs的筛选采用生物信息学技术共搜索到1857个REPs,并从中选择出回文结构较好的5条序列进行合成,见表1。M1-M5为合成的样本ODNs,MC1为阳性对照ODN 2216(A型人TLR9激动剂),MC2为阴性对照。
表1合成的ODNs序列Tab.1Synthetic ODN sequences

名称序列(5'—3')长度/bpNameSequence(5'—3')length(bp)M1GTAGATTTGACGGCAAAT18M2GTAGCGCCCGGACTTCAC-CGGGCG24M3GTAGCGGAAGTTTTCCG17M4GTAGGTTTCGATGAAAC17M5GTAGCCGCCCCAAGGGCG18MC1GGGGGACGATCGTCGGGGGG20MC2GGGGGAGCATGCTAGGGGGG20
2.2脂质体转染前后RAW264.7的细胞状态培养小鼠巨噬细胞RAW264.7,取对数增长期细胞铺板,脂质体转染合成的ODNs,转染6 h后细胞状态,见图1。

(A) Before transfection; (B) Transfection of ODNs for 6 h
图1脂质体转染前后RAW264.7的细胞状态。(A) 脂质体转染前;(B) 脂质体转染6h后
Fig.1Morphology of RAW264.7 cells before and after transfection of ODNs with Lipofectamine
2.3牛种布氏菌REPs对巨噬细胞分泌IFN-α的影响ELISA结果显示,M1、M2、M3、M4、M5共5个序列的IFN-α分泌浓度大于阴性序列MC2(图2)。采用t检验分别比较上述序列与阴性对照组之间IFN-α浓度的差异,结果显示,M4、M5序列与阴性对照序列相比,IFN-α浓度差异有统计学意义(t=4.649,P=0.010;t=16.572,P=0.000),提示上述序列可引起炎性因子IFN-α分泌的增加。
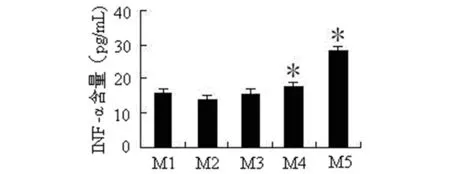
注:数字代表不同序列分泌IFN-α含量,*代表与MC2对照序列相比,IFN-α含量差异有统计学意义(P<0.05)。
The numbers represent the IFN-α production induced by REP sequences, *P<0.05 as compared with control sequence MC2.
图2不同REPs转染细胞后诱导IFN-α的分泌
Fig.2Induction of IFN-αproduction by REP sequences fromBrucellaabortus
2.4TLR9-siRNA对牛种布氏菌REPs所介导的IFN-α分泌的影响阳性ODN序列M5+TLR9-siRNA组与阴性对照MC2组相比所介导的IFN-α分泌无统计学意义(t=0.975,P=0.385),与M5+阴性siRNA组比较,M5+TLR9-siRNA组IFN-α分泌显著降低(t=16.525,P=0.000)。提示IFN-α分泌可以被TLR9-siRNA显著抑制。与M5序列相比,M5+阴性siRNA组所介导的IFN-α分泌没有明显影响(t=2.034,P=1.112),与阴性对照MC2相比,M5+阴性siRNA组IFN-α分泌有统计学意义(t=14.835,P=0.000)(图3)。这些结果提示,牛种布氏菌REPs在巨噬细胞中是经TLR9信号通路影响IFN-α的分泌,进而影响巨噬细胞的免疫应答。

注:与M5+阴性siRNA组相比,IFN-α分泌含量P<0.05;与MC2相比,IFN-α分泌含量P<0.05。
*P<0.05 as compared with M5+ negative siRNA; #P<0.05 as compared with control sequence MC2.
图3TLR9-siRNA对牛种布氏菌REPs所介导的IFN-α分泌的影响
Fig.3Effects of TLR9-siRNA on the levels of IFN-α mediated by REPs fromBrucellaabortusDNA
3讨论
布氏菌是革兰氏染色阴性胞内寄生菌,进入机体后主要以巨噬细胞与胎盘滋养层细胞为宿主细胞。在布氏菌细胞免疫机制的研究中,过去大多针对脂多糖和外膜蛋白等引起免疫应答的抗原进行探讨。Magnusson等发现,革兰氏阴性细菌的REPs序列具有免疫活化功能[6]。研究还发现,含GTAG基序的REPs对人和大部分家畜的免疫细胞刺激作用较强[8]。然而,布氏菌基因组DNA中是否存在具有免疫活性的REPs,目前未见报道。本文采用生物信息学方法识别牛种布氏菌DNA中的含有GTAG基序的REPs,通过转染培养的小鼠巨噬细胞RAW264.7,发现2个REPs序列(M4、M5)能够诱导RAW264.7细胞分泌IFN-α,提示其具有免疫活化功能。
TLRs是天然免疫中重要的模式识别受体,其中,TLR9是TLRs家族成员之一,存在于全身各组织中。TLR9主要通过识别未甲基化的CpG 基序来激活免疫细胞[9]。其识别病原菌DNA后胞内髓样分化因子88(Myd88)c一端与TIR区发生同源性相互作用,DD端募集含有“死亡”功能域的相关激酶(IRAKs)家族,导致IRAKs磷酸化进入胞浆,激活NF-κB,激酶抑制物(IKKs),使得核转录因子NF-κB的抑制成分IKB磷酸化而降解并与NF-κB分离,使核因子NF-κB激活而转入细胞核中,诱导相关基因的表达,分泌炎症因子(如IFN-α,IL.6,IL.8),化学趋化因子等,发挥转录调控作用。REPs含有丰富的非甲基化CpG基序,并且是回文结构(即茎环二级结构),在DNA生理学和基因组可塑性中起着重要作用[10]。因此,我们猜测,牛种布氏菌DNA来源的REPs序列可能通过TLR9诱导巨噬细胞IFN-α的分泌。为了证实上述猜测,我们使用TLR9-siRNA沉默小鼠巨噬细胞(RAW264.7)TLR9表达后,发现M5介导的IFN-α分泌被显著抑制,提示TLR9信号通路介导了REPs诱导的巨噬细胞IFN-α的分泌。
目前,化学合成的寡核苷酸作为TLR9激动剂已广泛应用于肿瘤、感染性疾病、哮喘等疾病的治疗[11]。从牛种布氏菌DNA发现具有活化TLR9活性的REPs序列,不仅有助于深入探讨布氏菌致病机制与细胞免疫机制,还有助于发现新的TLR9激动剂先导结构,为布鲁士菌病的防治提供新的思路。
参考文献:
[1] Galińska EM, Zagórski J. Brucellosis in humans—etiology, diagnostics, clinical forms[J]. Ann Agric Environ Med, 2013, 20(2): 233-238.
[2] Li TF,Liu FQ,Wang Y,et al.Surveillance results analysis of human brucellosis in 2011[J]. Chin J Ctrl Endem Dis,2013,28(1),46-47.(in Chinese)
李铁锋,刘凤岐,王赢等.2011 年全国布鲁杆菌病监测结果分析[J].中国地方病防治杂志,2013,28(1),46-47.
[3] Celli J. Surviving inside a macrophage: the many ways ofBrucella[J]. Res Microbiol, 2006, 157(2): 93-98. DOI:10.1016/j.resmic.2005.10.002
[4] Gomes MT, Campos PC, de Almeida LA, et al. The role of innate immune signals in immunity toBrucellaabortus[J]. Front Cell Infect Microbiol, 2012, 2: 130. DOI: 10.3389/fcimb.2012.00130
[5] Oliveira SC, de Almeida LA, Carvalho NB, et al. Update on the role of innate immune receptors duringBrucellaabortusinfection[J]. Vet Immunol Immunopathol, 2012, 148(1/2): 129-135. DOI: 10.1016/j.vetimm.2011.05.036
[6] Magnusson M, Tobes R, Sancho J, et al. Cutting edge: natural DNA repetitive extragenic sequences from gram-negative pathogens strongly stimulate TLR9[J]. J Immunol, 2007, 179(1): 31-35. DOI:10.4049/jimmunol.179.1.31
[7]Zhu J,Hu D,Liu LN,et al.Research on the role of capsular sialic acid inStreptococcussuisactivate macrophage TLR2-AKT-NF-κB signaling pathway[J].Microbiol China,2013, 40(6): 1058-1067. (in Chinese)
朱静,胡丹,刘丽娜等.荚膜唾液酸对猪链球菌激活巨噬细胞TLR2-AKT-NF-κB 信号通路影响的研究[J]. 微生物学通报,2013,40(6):1058-1067.
[8] Di Nocera PP, De Gregorio E, Rocco F. GTAG- and CGTC-tagged palindromic DNA repeats in prokaryotes[J]. BMC Genomics, 2013, 14: 522. DOI: 10.1186/1471-2164-14-522
[9] Manuja A, Manuja BK, Kaushik J, et al. Immunotherapeutic potential of CpG oligodeoxynucleotides in veterinary species[J]. Immunopharmacol Immunotoxicol, 2013, 35(5): 535-544. DOI: 10.3109/08923973.2013.828743
[10] Jaroslav N, Tereza H, Irena L. Identification and characterization of repetitive extragenic palindromes (REP)-associated tyrosine transposases: implications for REP evolution and dynamics in bacterial genomes[J]. BMC Genomics, 2010, 11: 44. DOI: 10.1186/1471-2164-11-44
[11] Gosu V, Basith S, Kwon OP, et al. Therapeutic applications of nucleic acids and their analogues in Toll-like receptor signaling[J]. Molecules, 2012, 17(11): 13503-13529. DOI: 10.3390/molecules171113503
DOI:10.3969/j.issn.1002-2694.2016.05.006
通讯作者:于慧,Email:huiyu2008@hotmail.com
中图分类号:R183.7
文献标识码:A
文章编号:1002-2694(2016)05-447-04
Corresponding author:Yu Hui, Email: huiyu2008@hotmail.com
收稿日期:2015-08-04修回日期:2016-02-22
Screening and detection of repetitive extragenic palindromic sequences with TLR9 agonistic activity fromBrucellaabortusDNA
ZHANG Ya-xian1.2, BAI Li-yun2, WANG Zhan-li2, WANG Ying2, YU Hui3
(1.BaotouMedicalCollege,Baotou014040,China;2.TheFirstAffiliatedHospitalofBaotouMedicalCollege,Baotou014040,China;3.TheSecondAffiliatedHospitalofBaotouMedicalCollege,Baotou014040,China;4.ChifengCityforDiseaseControlandPrevention,Chifeng024000,China)
Abstract:The aim of this study was to screen natural DNA sequences named repetitive extragenic palindromic sequences (REPs) with TLR9 agonistic activity from Brucella abortus DNA. Complete genome sequence of Brucella abortus A13334 was available in GenBank. A total of 1 857 REP sequences in Brucella abortus was identified. Bioinformatics technology was used to identify Brucella abortus REPs. Five REP sequences were synthesized for further detecting the TLR9 agonistic activity. Phosphodiester ODNs representing the selected ODNs were then synthesized. Murine macrophage RAW264.7 were cultured and transfected with ODNs mediated by lipofectamine 3000. The content of IFN-α in the supernatant was then measured by ELISA. We found that IFN-α expression was significantly increased in RAW264.7 treated with M4 and M5. In addition, TLR9-siRNA was used to down regulate TLR9 expression. IFN-α levels were examined in RAW264.7 transfected with TLR9-siRNA upon ODN stimulation. Results showed TLR9-siRNA could significantly reduce the levels of IFN-α in RAW264.7 treated with M5. This work demonstrated that REP sequences present in Brucella abortus DNA were able to produce macrophages stimulation through TLR9.
Key words:Brucella abortus; repetitive extragenic palindrome sequence; macrophages; toll-like receptor 9; IFN-α Supported by the the Natural Science Foundation of China(No.81360242) and China National Funds for Distinguished Young Scientists(No.2014JQ04) and the Natural Science Foundation of Inner Mongolia(Nos.2014MS0808,2015 MS0810)
国家自然科学基金(No.81360242);内蒙古杰出青年培育基金(No.2014JQ04)和内蒙古自治区自然科学基金(No.2014MS0808,No.2015MS0810)联合资助
