长江江豚TRAIL基因的克隆、体外表达及生物学功能分析
2016-07-04裴丽丽章纬菁卢佳黄芳曹倩倩任文华
裴丽丽,章纬菁,卢佳,黄芳,曹倩倩,任文华
1 南京师范大学 生命科学学院 江苏省分子医学重点实验室,江苏 南京 2100462 南京医科大学 康达学院,江苏 连云港 222000
长江江豚TRAIL基因的克隆、体外表达及生物学功能分析
裴丽丽1,2,章纬菁1,卢佳1,黄芳1,曹倩倩1,任文华1
1 南京师范大学 生命科学学院 江苏省分子医学重点实验室,江苏 南京 210046
2 南京医科大学 康达学院,江苏 连云港 222000
裴丽丽, 章纬菁, 卢佳, 等. 长江江豚TRAIL基因的克隆、体外表达及生物学功能分析. 生物工程学报, 2016, 32(5): 610–620.
Pei LL, Zhang WJ, Lu J, et al. Molecular cloning, in vitro expression and bioactivity of TRAIL (TNFSF10) gene from finless porpoises. Chin J Biotech, 2016, 32(5): 610–620.
摘 要:构建可溶性肿瘤坏死因子相关凋亡诱导配体 (TRAIL) 基因的表达体系,研究其蛋白表达产物对肿瘤细胞凋亡的影响,为以后江豚免疫系统的研究奠定基础。通过RT-PCR技术从江豚Neophocaena phoconoides血液总RNA中反转录扩增出肿瘤坏死因子相关凋亡诱导配体 (简称fTRAIL) 的全长cDNA序列,并将fTRAIL的胞外可溶性 (简称fsTRAIL) 片段连接入表达载体pET43.1a中,在大肠杆菌BL21 (DE3) 中表达并纯化,Western blotting对产物Nus-His-fsTRAIL蛋白进行鉴定。体外用MTT法、台盼蓝拒染法及流式细胞术检测Nus-His-fsTRAIL蛋白对Jurkat细胞和HeLa细胞的影响。成功构建了fTRAIL胞外可溶性片段 (简称fsTRAIL) 与pET43.1a组成的表达载体,并获得Nus-His-fsTRAIL蛋白。体外实验表明,Nus-His-fsTRAIL蛋白能够以剂量依赖的方式抑制Jurkat和HeLa细胞的增殖并诱导其凋亡。Nus-His-fsTRAIL表达产物具有对Jurkat和HeLa细胞体外抗肿瘤活性的作用。
关键词:TRAIL,江豚,细胞凋亡,Jurkat细胞,HeLa细胞
Received: October 10, 2015; Accepted: March 14, 2016
Supported by: National Natural Science Foundation of China (No. 31370401).
国家自然科学基金 (No. 31370401) 资助。
肿瘤坏死因子 (TNF) 是1975年Carswell等发现的一种能使肿瘤细胞凋亡、坏死的细胞因子。细胞凋亡是一个内在基本的细胞程序化死亡过程,在多细胞有机体的发展和组织稳态的维持中起着关键的作用[1]。TRAIL是在其序列同源性的基础上,于1995年由Wiley首次发现[2],与肿瘤坏死因子配体家族的其他成员一样,TRAIL对体外的细胞系包括一些肿瘤细胞系具有诱导细胞凋亡的作用。与其他TNF家族成员相比,TRAIL具有两个特性:一是TRAIL在大多数组织中都能表达,而其他TNF家族成员的表达受到严格调控并在活化的细胞中暂时表达[3]。第二,TRAIL是一个促凋亡配体,目前发现在一些肿瘤和炎症细胞中能够高表达,如小儿恶性肿瘤[4]、肾脏疾病[5]、类风湿关节炎、骨关节炎和脊柱关节病等[6],但其对正常细胞无明显毒性[7],这意味着,TRAIL在癌症治疗中具有潜在的治疗作用[8-11]。TRAIL在与其受体结合时形成三聚体,并诱导其受体也形成三聚体而相互结合,从而选择性地诱导肿瘤细胞发生凋亡。这些特性预示着TRAIL在治疗肿瘤方面有着广泛的应用前景。
1 材料与方法
1.1材料
1.1.1实验动物、细胞和宿主菌
江豚新鲜外周血从安徽铜陵繁育中心淡水长江江豚体检期间获得;实验用细胞系Jurkat细胞与HeLa细胞由本实验室保存;大肠杆菌E. coli Top10、DH5α、BL21和pET-43.1a由本实验室保存;pMD19-T载体购于TaKaRa公司。
1.1.2试剂
动物血液RNA提取试剂盒、PrimeScriptTM1st Strand cDNA合成试剂盒、pMD19-T载体试剂盒、Taq DNA聚合酶、感受态细胞制备试剂盒、蛋白Marker、荧光定量PCR试剂盒、rTaq酶和限制性内切酶等均购自TaKaRa公司;割胶回收试剂盒及质粒小提试剂盒均购自Axygen公司;T4 DNA连接酶、碱性磷酸酶、磷酸激酶和预染蛋白Marker等均购自Fermentas公司;鼠His6一抗和辣根过氧化酶标记的山羊抗鼠IgG购自TIANGEN公司;FITC标记的羊抗鼠IgG抗体 (R&D Systems Inc.,USA) 和IPTG购自Promega公司;所有引物合成及测序由上海英骏公司完成;其他试剂均为国产分析纯。
1.2方法
1.2.1江豚外周血总RNA的提取
使用动物血液RNA提取试剂盒 (RNeasy Mini Kit, Qiagen) 提取江豚外周血总RNA,实验方法步骤按照试剂盒中的说明书进行。
1.2.2江豚TRAIL (fTRAIL) 基因的克隆
通过BLAST,在NCBI数据库中获得牛(GenBank Accession No. AC_000158.1)、羊(GenBank Accession No. ADL27922) 和猪(GenBank Accession No. ABV21385) 的TRAIL基因[12],根据同源性比对利用Primer 5.0软件[13]设计一对简并引物:正义引物fTRAIL F1和反义引物fTRAIL F2 (表1)。用上述简并引物进行RT-PCR试验。扩增参数:95 ℃ 5 min;35个循环 (94 ℃ 30 s,56 ℃ 30 s,72 ℃ 1 min);最后72 ℃孵育7 min。RT-PCR产物用1%的琼脂糖凝胶电泳分离后,观察并记录结果。按Axygene公司的DNA胶回收试剂盒操作说明进行割胶回收约870 bp的DNA片段。
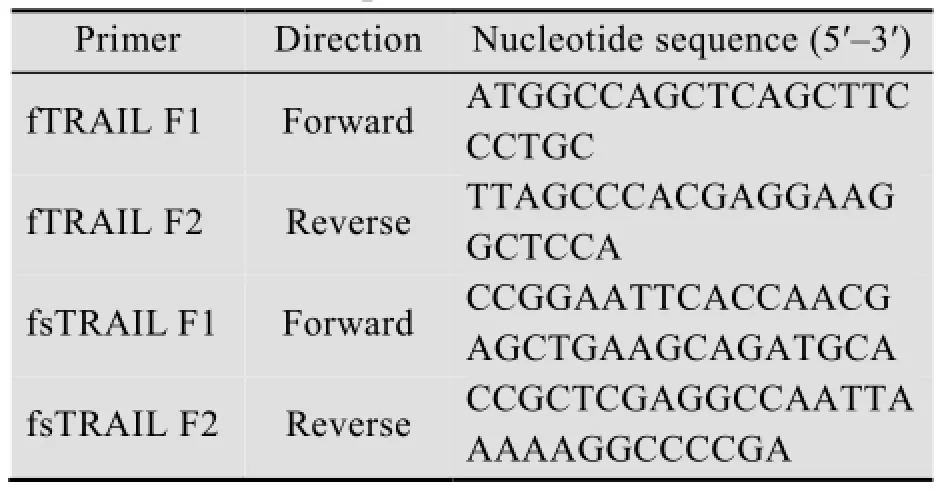
表1 引物序列Table 1 Primer sequences
1.2.3生物信息学分析
为克隆fTRAIL cDNA序列并对所得序列进行生物信息学分析,使用以下软件和网站:1) 从NCBI网站获得TRAIL基因序列、氨基酸序列,以及fTRAIL的开放阅读框 (Open reading frame, ORF),并对相应的氨基酸序列进行预测;2) 使用3D-JIGSAW和SWISS-MODEL服务器预测三级结构,并用RasMol软件查看;3) 使用Clastal W软件[14]进行多重比对分析;4) 在序列多重比对的基础上,使用MEGA 5.0 软件中的Maximum Likelihood法构建系统树。
1.2.4融合表达载体pET43.1a-fsTRAIL的构建
研究表明,TRAIL的可溶型是其功能发挥的主要形式,根据fTRAIL的胞外可溶区(fsTRAIL) 设计一对含有pET43.1a酶切位点的基因特异性引物:fsTRAIL F1 (含EocRⅠ位点) 与fsTRAIL F2 (含XhoⅠ位点) (表1)。PCR反应程序:94 ℃ 5 min ;35个循环 (94 ℃ 30 s;56 ℃30 s;72 ℃ 1 min);最后72 ℃延伸7 min。将PCR反应产物经电泳、纯化回收PCR产物(870 bp左右)。以EcoRⅠ、XhoⅠ酶切PCR产物与pET43.1a载体,然后进行pET43.1a载体的去磷酸化与fsTRAIL目的片段的磷酸化,两者进行连接反应,构建pET43.1a-fsTRAIL表达载体。
1.2.5连接产物的转化、表达及纯化鉴定
原核表达载体pET43.1a-fsTRAIL在大肠杆菌中的表达。首先将原核表达载体pET43.1afsTRAIL转入大肠杆菌BL21 (DE3),挑取含表达质粒的转化菌株,于LB培养基 (含Amp+)中,37 ℃、220 r/min培养至A600达0.4–0.6时,加入1 mol/L IPTG至终浓度为0.2 mmol/L,16 ℃、100 r/min振荡培养24 h,诱导pET-43.1a-fsTRAIL重组蛋白可溶性的表达。培养液在冰上超声破碎后,分离上清和沉淀,用SDS-PAGE检测蛋白的表达。然后用镍柱亲和层析纯化可溶性重组蛋白Nus-His-fsTRAIL,取出少量蛋白进行SDS-PAGE电泳,将其余蛋白质溶液放于4 ℃冰箱备用。用1× PBS (pH 8.0)对收集的蛋白液进行透析浓缩后根据需要保存于–70 ℃。按同样的方法纯化空载体pET-43.1a的表达产物,即Nus-His标签蛋白。通过Western blotting鉴定纯化的蛋白表达产物。
1.2.6重组可溶性蛋白生物活性测定
对纯化的Nus-His-fsTRAIL蛋白进行生物学活性检测,并以Nus-His标签蛋白作为对照。采用倒置显微镜及台盼蓝拒染法观察不同浓度上述两种蛋白在不同时间对白血病T细胞(Junkat细胞) 和人宫颈癌细胞 (HeLa细胞) 形态的影响;MTT法检测在不同浓度作用下,重组蛋白对肿瘤细胞的抑制作用;经MTT法和台盼蓝拒染法检测后,选取适宜浓度的重组蛋白及适宜的时间梯度,使用流式细胞术检测重组蛋白对Junkat细胞的促凋亡作用。
2 结果与分析
2.1江豚全长fTRAIL cDNA和预测氨基酸序列分析
江豚TRAIL的开放阅读框为864 bp,编码一个288个氨基酸的fTRAIL蛋白质。克隆所得全长cDNA序列和预测的氨基酸序列如图1A所示。fTRAIL的cDNA序列已提交GenBank数据库 (GenBank Accession No. KF151167)。江豚TRAIL序列含预测的细胞质结构域,一个跨膜结构域 (氨基酸18–38) 和一个由250个氨基酸组成的胞外结构域。它具有TNF结构域、一个保守的半胱氨酸残基和预测的糖基化位点。fTRAIL是典型的II型跨膜蛋白 (图1B)。在氨基酸水平上,用Clustal软件分析显示,江豚和马、人、狗及牛之间的序列同源性分别为86%、79%、82%和80% (图1B)。
2.2江豚和人TRAIL胞外区氨基酸序列比对及其蛋白质空间结构分析
人TRAIL (hTRAIL) 胞外区与受体结合部位为C端的114–281位氨基酸,江豚TRAIL (fTRAIL) 胞外区与受体结合部位为C端的119–289位氨基酸,如图2所示,hTRAIL (114–281) 与fTRAIL (119–289) 氨基酸序列比对,相似度为83.3%以上。尽管二者在氨基酸水平上存在一定的差异,但预测fTRAIL的胞外功能区3D结构 (c, d) 和hTRAIL 胞外区3D结构(a, b) 非常相似。这表明江豚的TRAIL与人TRAIL可能具有相似的生物学功能。

图1 fTRAIL基因的cDNA序列和氨基酸序列Fig. 1 The nucleotide and deduced amino acid sequences of fTRAIL. (A) The nucleotide and deduced amino acid sequences of fTRAIL. The initiation codon and the stop codon are boxed, and the stop codon is indicated by an asterisk (*). The putative transmembrane domain is underlined. (B) Amino-acid sequence alignment of TRAIL from several species: finless porpoise, human, dog, cow, and horse were performed using Clustal W software. Identical amino acids among all sequences are indicated by “*”, whereas those with high or low similarity are indicated by “:”and “.” respectively. The putative transmembrane domain is marked by gray shadings. The TNF superfamily signature is boxed. The conserved cysteine and asparagine site are marked by red shadings.

图2 fsTRAIL (aa119-289) 与hsTRAIL (aa 114-281) 胞外区氨基酸序列比对及其蛋白质空间结构比较Fig. 2 Amino acid sequence alignment and homology modelling of the fsTRAIL monomer (aa 119-289) based on the human structure (aa 114-281). (A) The amino acid sequence alignment of fsTRAIL and human sTRAIL. (B) The homology modelling of the fsTRAIL and human sTRAIL. For cartoons display and sphere display of structure models: yellow represents β strands, red represents α-helix, blue represents turns and whites represent other residues. (a) and (b) represent the cartoon display and sphere display of the 3D structure of human sTRAIL, respectively. (c) and (d) represent the cartoon and sphere displays of the 3D structure of fsTRAIL, respectively.
2.3系统进化树分析
为确定江豚TRAIL基因在进化中的位置,运用Maximum Likelihood法构建了20个脊椎动物的TRAIL序列的系统进化树。如图3A所示,系统树主要分两个分支,一个是鱼类的分支,一个是哺乳类和鸟类的分支。江豚TRAIL与哺乳动物的偶蹄目在进化上接近,而与其他非哺乳动物的TRAIL序列较远。为进一步确定江豚在进化上的位置,我们又用GILT基因构建了系统进化树,如图3B所示,江豚与偶蹄目哺乳动物在进化上依然比较接近。以上结果表明江豚可能与偶蹄目哺乳动物具有共同的原始祖先。
2.4重组蛋白的表达、纯化和鉴定
为检测Nus-His-fsTRAIL蛋白的生物学活性,将重组pET43.1a-fsTRAIL导入大肠杆菌BL21 (DE3) 中表达。经SDS-PAGE检测 (图4)显示,在大约90 kDa处为目的蛋白,即Nus-HisfsTRAIL蛋白,经IPTG诱导24 h达到最大表达量。经Ni2+-NTA柱纯化后,收集的蛋白经SDS-PAGE分析和Western blotting检测,结果见图4。
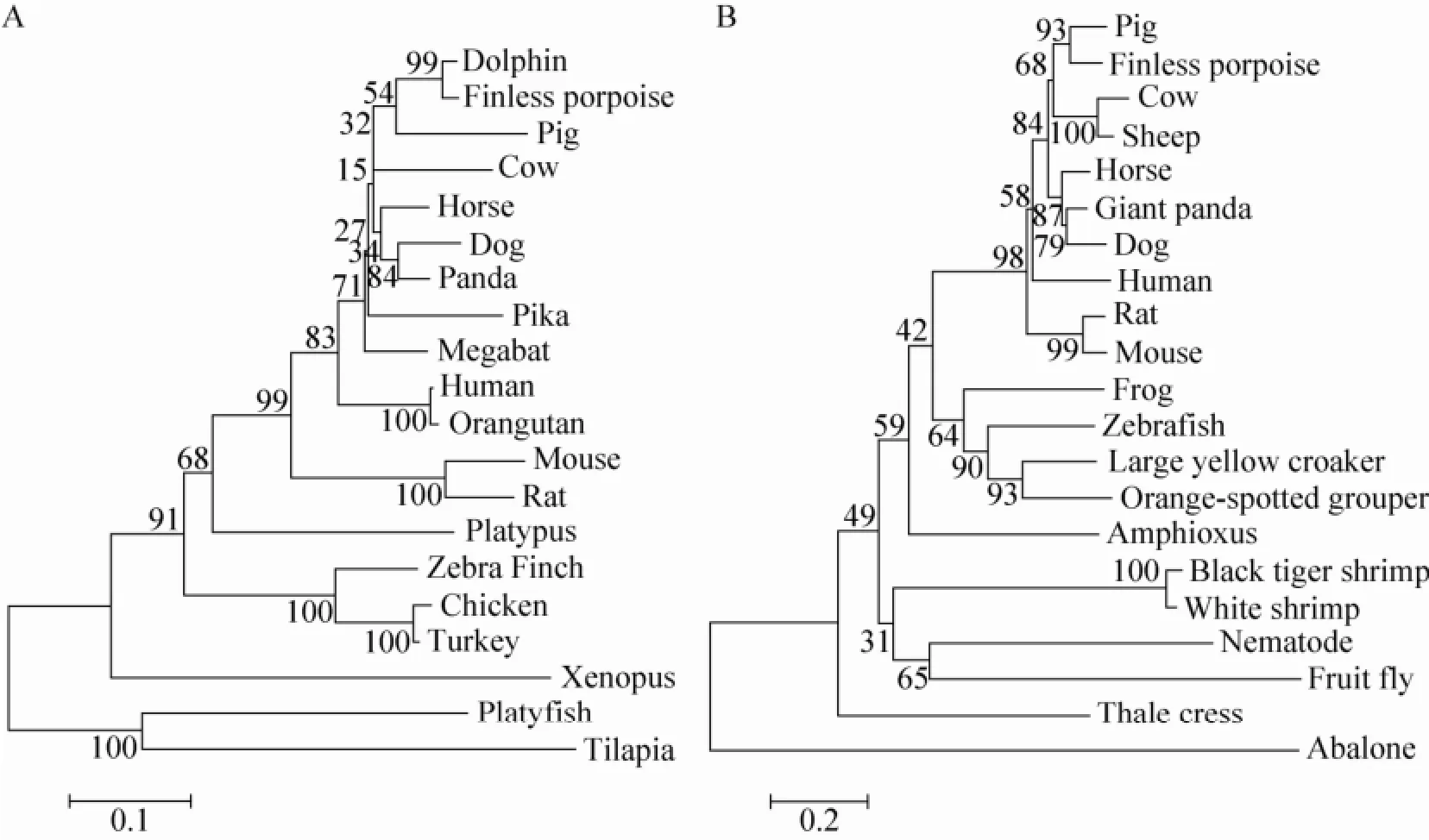
图3 最大似然法构建系统进化树Fig. 3 The phylogenetic tree was constructed by the Maximum Likelihood method using MEGA 5.0 packages and was bootstrapped 1 000 times. (A) Phylogenetic tree showing the relationship between fTRAIL DNA sequences and other identified TRAIL sequences. (B) The phylogenetic tree showing the relationship of GILT sequences from different species.
2.5MTT法检测Nus-His-fsTRAIL蛋白
采用MTT法检测Nus-His-fsTRAIL蛋白对Jurkat与HeLa肿瘤细胞的作用。从图5中可以看出,以Nus-His标签蛋白作为对照,在不同浓度下,Nus-His-fsTRAIL蛋白表现出抑制细胞增殖的作用,而且这种作用呈剂量依赖性。
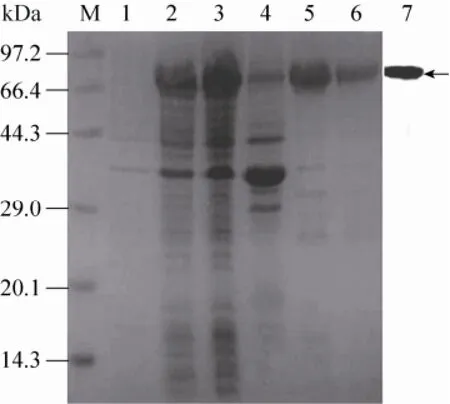
图4 SDS-PAGE分析Nus-His-fsTRAIL蛋白在大肠杆菌中的表达Fig. 4 SDS-PAGE analysis of Nus-His-fsTRAIL protein expressed in E. coli BL21 (DE3). M: low molecular weight marker. Lane 1: cell lysates of bacteria transformed with pET43.1a-fsTRAIL without IPTG induction; Lane 2: induced recombinant BL21; Lane 3: the recombinant BL21 supernatant; Lane 4: the recombinant BL21 precipitation; Lane 5: the fusion protein purified by the Ni-IDA affinity chromatography; Lane 6: the secondary chromatography from Lane 5 protein by the Ni-IDA affinity chromatography; Lane 7: the Western blotting analysis of purified Nus-His-fsTRAIL fusion protein using mAb against His6-tag. The arrow represents the location of Nus-His-fsTRAIL fusion protein.
2.6Nus-His-fsTRAIL蛋白对Jurkat和HeLa细胞作用的形态观察
采用不同浓度梯度,在光学显微镜下观察Nus-His-fsTRAIL蛋白对Jurkat和HeLa细胞作用,并选择反应36 h后进行台盼蓝拒染实验。从图6中可以看出随着时间和浓度的增加,细胞发生了明显的破裂和死亡。
2.7流式细胞仪检测Nus-His-fsTRAIL蛋白的促Jurkat细胞凋亡作用
Nus-His标签蛋白与Nus-His-fsTRAIL蛋白分别对Jurkat细胞作用36 h后,进行流式细胞仪检测。结果显示,Nus-His标签蛋白对Jurkat细胞作用后,在凋亡区FITC (+)/PI(–) 检测到少量细胞(5.67%) (图7A);Nus-His-fsTRAIL对Jurkat细胞作用后,在早期凋亡区FITC (+)/PI(–) 检测到少量细胞 (7.71%),在晚期凋亡区FITC (+)/PI(+) 检测到较多的细胞 (40.97%) (图7B)。

图5 Nus-His-fsTRAIL蛋白对Jurkat和HeLa细胞活性的影响Fig. 5 Effect of Nus-His-fsTRAIL protein on the viability of Jurkat and HeLa cell lines by MTT assay. (A) The Jurkat cells were treated with different concentrations of the Nus-His-fsTRAIL or Nus-His tag protein at indicated amounts. (B) HeLa cells were treated with different concentrations of the Nus-His-fsTRAIL or Nus-His tag protein at indicated amounts. The cell viability was examined after treatment with MTT 24 h. Each treatment was performed in triplicate. Values ares of three independent experiments.

图6 Nus-His-fsTRAIL蛋白对Jurkat和HeLa细胞作用后形态学的改变Fig. 6 Morphology of Jurkat and HeLa cell was visualized under a Nikon Eclipse TE2000-U inverted phase-contrast microscope (200×) equipped with digital camera. (A) and (B) The morphology of Jurkat cells and HeLa cells were photographed under an inverted microscope after treating with different concentrations and different time of the Nus-His-fsTRAIL protein or Nus-His tag protein at indicated amounts, respectively. (C) and (D) Jurkat cells and HeLa cells were stained by Trypan Blue after treating with different concentrations at 36 h of the Nus-His-fsTRAIL protein or Nus-His tag protein at indicated amounts, respectively.

图7 流式细胞仪检测Nus-His-fsTRAIL蛋白对Jurkat细胞的促凋亡作用Fig. 7 The apoptotic effect of fsTRAIL on the Jurkat cell was examined by flow cytometry. (A) Jurkat cells were treated with 800 ng/mL of the Nus-His tag protein. (B) Jurkat cells were treated with 800 ng/mL of the Nus-His-fsTRAIL protein.
3 讨论
与陆生哺乳动物一样,水生哺乳动物的免疫系统包括天然免疫和适应性免疫。目前,许多脊椎动物的肿瘤坏死因子相关凋亡诱导配体(TRAIL) 的基因已成功克隆[15-16]。然而,水生哺乳动物TRAIL仍未见报道。本实验成功克隆了江豚TRAIL的CDS序列并进行了测定。这是第一个被克隆并鉴定的水生哺乳动物TRAIL基因。克隆策略是依据江豚与其他哺乳动物序列之间的同源性进行的。
江豚TRAIL (fTRAIL) 也具有TRAIL蛋白的典型特征。首先,fTRAIL蛋白也是一个Ⅱ型跨膜蛋白,有跨膜区且没有启动子;其次,fTRAIL具有N端非保守区,而C端不同物种之间具有较大的保守性;第三,人TRAIL (hTRAIL)蛋白的Cys230是TRAIL发挥功能所必需的,通过形成分子间二硫键诱导凋亡[17],而fTRAIL也具有Cys230。据报道,鲸类与有蹄类动物 (偶蹄目) 非常相似[18]。我们的研究结果表明,与其他氨基酸序列相比,江豚TRAIL与偶蹄目动物TRAIL比如猪有88%相似、与牛相似度有80% (图2.1b)。但fTRAIL与非洲爪蟾和岩鸽只有46%的相似性,与其他无脊椎动物 (包括果蝇、海鞘和鲍鱼) 只有15%–20%的相似性,这也预示着fTRAIL可能与偶蹄目动物具有相同的祖先,可以为鲸类免疫进化的研究提供一定参考。虽然氨基酸序列比对显示fTRAIL序列与偶蹄目序列具有高度相似性,但在某些氨基酸位点上也存在明显差异,这表明,与陆生哺乳动物相比,鲸类哺乳动物为了适应水中的环境可能进行了某些适应性进化。
通过氨基酸序列比对构建系统进化树,获得了江豚与其他脊椎动物的亲缘关系,结果表明江豚与偶蹄目哺乳动物的亲缘关系最近。这一观点同时也被江豚同其他物种的GILT氨基酸序列比对所验证[19]。
人TRAIL与受体结合的胞外区为C端的114–281位氨基酸,为TRAIL的活性部分,有一个 N-糖基化位点并有蛋白酶作用位点,可以被蛋白酶从膜上剪切下来,形成可溶性分子sTRAIL[8]。因此,我们将人sTRAIL (114–281)与江豚的fsTRAIL (119–289) 进行了氨基酸序列比对,结果表明,它们的相似度在83.3%以上,即与受体结合部位的氨基酸序列基本一致。通过蛋白质的3D结构预测分析,人sTRAIL与江豚的fsTRAIL蛋白具有类似的三维结构。重要空间结构的氨基酸是非常保守的,这表明fsTRAIL可能也具有人sTRAIL类似的功能,能够与人肿瘤细胞上的相应受体相结合,这为后续对江豚TRAIL胞外可溶片段的表达产物的活性检测奠定基础。此外,因为我们很难获得江豚的新鲜组织材料,TRAIL基因在各种器官组织 (心脏、肾脏、脾脏、肝脏、肺和肠等) 的表达水平无法检测,但根据目前的研究表明,TRAIL基因的mRNA在脾、肝和肾中有高水平表达[20]。
为了研究fsTRAIL的活性,我们曾将其连接入pET28表达载体,并在BL21 (DE3) 中表达。结果蛋白存在于包涵体中,包涵体复性后没有获得可溶性蛋白。为了增强蛋白的可溶性,我们将其构建入pET43.1a表达载体,并进行低温诱导 (16 ℃),得到了较高水平的可溶性表达。
文献报道人、 红鳍东方鲀或鳜鱼的sTRAIL 能够诱导人肿瘤细胞系的凋亡[2,20-21],因此推测fsTRAIL也具有相似作用。为了检测fsTRAIL诱导肿瘤细胞凋亡的能力,我们首先用MTT法检测其对Jurkat和Hela细胞作用,实验结果表明Nus-His-fsTRAIL蛋白能以剂量依赖的方式抑制细胞增殖,不难发现Nus-His-fsTRAIL蛋白对Jurkat细胞的抑制能力强于对Hela细胞的抑制能力,这可能是由于Nus-His-fsTRAIL蛋白与两种细胞上TRAIL受体结合能力不同所致。此外,经Nus-His-fsTRAIL蛋白处理后,Jurkat和HeLa细胞的形态学观察以及台盼蓝染色后的形态学观察表明fsTRAIL可以剂量依赖的方式诱导Jurkat和HeLa细胞死亡。为了进一步确定Nus-His-fsTRAIL蛋白的肿瘤细胞毒理效用,我们运用了流式细胞术进一步验证了Nus-HisfsTRAIL蛋白可以诱导肿瘤细胞凋亡,而对照Nus-His标签蛋白则未见此效应。综上所述,我们可以得出结论,fsTRAIL可抑制Jurkat细胞和HeLa细胞增殖及诱导凋亡。但fsTRAIL能否诱发其他人类肿瘤细胞株的凋亡还有待进一步研究。
总之,本次实验首次对水生哺乳动物TRAIL蛋白进行报道,发现其在进化上与偶蹄目动物的TRAIL具有同源性,也发现fTRAIL能抑制Jurkat和HeLa细胞增殖并诱导细胞凋亡,既为水生哺乳动物从陆生到水生的进化研究提供了依据,也为江豚甚至是水生哺乳动物的免疫机制和物种保护的研究奠定了基础。
REFERENCES
[1] Steller H. Mechanisms and genes of cellular suicide. Science, 1995, 267(5203): 1445–1449.
[2] Wiley SR, Schooley K, Smolak PJ, et al. Identification and characterization of a new member of the TNF family that induces apoptosis. Immunity, 1995, 3(6): 673–682.
[3] Goetz FW, Planas JV, MacKenzie S. Tumor necrosis factors. Dev Comp Immunol, 2004, 28(5): 487–497.
[4] Gasparini C, Vecchi Brumatti L, Monasta L, et al. TRAIL-based therapeutic approaches for the treatment of pediatric malignancies. Curr Med Chem, 2013, 20(17): 2254–2271.
[5] Lorz C, Benito A, Ucero AC, et al. Trail and kidney disease. Front Biosci (Landmark Ed), 2009, 14: 3740–3749.
[6] Mustafa MA, Mohamed UK, Bastawisy A. TNF-related apoptosis-inducing ligand levels in rheumatoid arthritis, osteoarthritis, and spondyloarthritis. Egypt J Int Med, 2012, 24(3): 72–78.
[7] Yang SB, Wu XZ. Identification and functional characterization of a human sTRAIL homolog, CasTRAIL, in an invertebrate oyster Crassostrea ariakensis. Dev Comp Immunol, 2010, 34(5): 538–545.
[8] Pitti RM, Marsters SA, Ruppert S, et al. Induction of apoptosis by Apo-2 ligand, a new member of the tumor necrosis factor cytokine family. J Biol Chem, 1996, 271(22): 12687–12690.
[9] Balkwill F. Tumour necrosis factor and cancer. Nat Rev Cancer, 2009, 9(5): 361–371.
[10] Fox NL, Humphreys R, Luster TA, et al. Tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) receptor-1 and receptor-2 agonists for cancer therapy. Expert Opin Biol Ther, 2010, 10(1): 1–18.
[11] Gerspach J, Pfizenmaier K, Wajant H. Therapeutic targeting of CD95 and the TRAIL death receptors. Recent Pat Anticancer Drug Discov, 2011, 6(3): 294–310.
[12] Glenney GW, Wiens GD. Early diversification of the TNF superfamily in teleosts: genomic characterization and expression analysis. JImmunol, 2007, 178(12): 7955–7973.
[13] Lalitha S. Primer premier 5. Biotech Software Internet Rep, 2000, 1(6): 270–272.
[14] Larkin MA, Blackshields G, Brown NP, et al. Clustal W and Clustal X version 2.0. Bioinformatics, 2007, 23(21): 2947–2948.
[15] Allen JE, El-Deiry WS. Regulation of the human TRAIL gene. Cancer Biol Ther, 2012, 13(12): 1143–1151.
[16] Chang MX, Nie P, Xie HX, et al. Characterization and expression analysis of TNF-related apoptosis inducing ligand (TRAIL) in grass carp Ctenopharyngodon idella. Vet Immunol Immunopathol, 2006, 110(1/2): 51–63.
[17] Seol DW, Billiar TR. Cysteine 230 modulates tumor necrosis factor-related apoptosis-inducing ligand activity. Cancer Res, 2000, 60(12): 3152–3154.
[18] Gatesy J. Whales and even-toed ungulates (Cetartiodactyla)//Hedges SB, Kumar S, Eds. The Timetree of Life. Oxford, UK: Oxford University Press, 2009: 511–516.
[19] Zhou LD, Yan WL, Yang L, et al. Isolation of gamma-interferon-inducible lysosomal thiol reductase (GILT) from the Yangtze finless porpoise. Dev Comp Immunol, 2013, 41(4): 652–656.
[20] Li JF, Ai HX, Zhang J, et al. Molecular cloning, functional characterization and phylogenetic analysis of TRAIL in Japanese pufferfish Takifugu rubripes. J Fish Biol, 2011, 79(3): 747–760.
[21] Gao Y, Chang MX, Sun BJ, et al. TRAIL in the mandarin fish Siniperca chuatsi: gene and its apoptotic effect in HeLa cells. Fish Shellfish Immunol, 2008, 24(1): 55–66.
(本文责编 郝丽芳)
环境生物技术
Molecular cloning, in vitro expression and bioactivity of TRAIL (TNFSF10) gene from finless porpoises
Lili Pei1,2, Weijing Zhang1, Jia Lu1, Fang Huang1, Qianqian Cao1, and Wenhua Ren1
1 Jiangsu Province Key Laboratory for Molecular and Medical Biotechnology, College of Life Sciences, Nanjing Normal University, Nanjing 210046, Jiangsu, China
2 Kangda College of Nanjing Medical University, Lianyungang 222000, Jiangsu, China
Abstract:To construct soluble TNF related apoptosis inducing ligand (TRAIL) expression system and investigate the effect of the expression product on tumor cell. It may provide valuable information for research into the immune system of the finless porpoise. The full-length cDNA of TRAIL (designated fTRAIL) was cloned from the total RNA of the finless porpoises blood using RT–PCR techniques and then the extracellular soluble fragments of fTRAIL (designated fsTRAIL) was ligated into pET43.1a. Recombinant soluble fTRAIL (pET43.1a-fsTRAIL) fused with Nus-his tag was efficiently expressed in Escherichia coli BL21 (DE3) and the Nus-His-fsTRAIL protein was purified. The expression of Nus-His-fsTRAIL was verified by Western blotting. In vitro, the effects of the purified Nus-His-fsTRAIL protein on Jurkat and HeLa cells were etected by the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrasodium bromide (MTT) assay, TrypanBlue and Flow Cytometry analysis. The expression system pET43.1a-fsTRAIL was constructed and Nus-His-fsTRAIL protein was expressed successfully. In vitro, the Nus-His-fsTRAIL protein was able to inhibit the proliferation and induce apoptosis of Jurkat and HeLa cells in a dose-dependent manner. The Nus-His-fsTRAIL protein has anti-tumor activity against Jurkat and HeLa cells in vitro.
Keywords:TRAIL, finless porpoises (Neophocaena phoconoides), apoptosis, Jurkat cells, HeLa cells
Corresponding author:Wenhua Ren. Tel/Fax: 86-25-85891163; E-mail: spidervenom@163.com
