睾丸Leydig细胞发育生物学研究进展*
2016-05-07马学综述董强审校
马学 综述 董强 审校
睾丸Leydig细胞发育生物学研究进展*
马学1综述 董强2审校
(1. 四川大学华西医院小儿泌尿外科;2. 四川大学华西医院泌尿外科, 成都 610041)
Leydig细胞是位于哺乳动物睾丸间质中的一种内分泌细胞,在促进胚胎期生殖器官的分化发育、男性第二性征的发育和维持、精子发生、激发性欲以及维持性功能和促进人体的新陈代谢等方面发挥重要作用。因此,针对Leydig细胞的起源、分化发育机制的研究已经成为当今国际男性生殖健康研究领域的前沿和热点。本文对该领域近年来的研究进展进行综述。
Leydig细胞;睾酮;发育生物学
Leydig细胞是哺乳动物睾丸间质中一种内分泌细胞,具有合成和分泌睾酮(Testosterone, T)的功能,是男性体内雄激素的最主要来源,其功能受下丘脑-垂体性腺轴的调控[1]。其分泌的睾酮通过血液循环被运输到全身各处的靶器官, 再通过与雄激素受体结合进而在促进胚胎期生殖器官的分化发育、男性第二性征的发育和维持、精子发生、激发性欲以及维持性功能和促进人体的新陈代谢(如促进蛋白质合成、骨骼生长及红细胞生成等)等方面发挥重要的作用[1,2]。几十年来,伴随着工业化社会的发展,越来越多的与环境污染密切相关的内分泌干扰物质可以通过影响胚胎Leydig细胞的结构和功能干扰胚胎期生殖器官的发育,从而导致泌尿生殖系统先天性畸形疾病(例如隐睾、尿道下裂、真/假两性畸形等)的发生率逐年上升,给众多患儿及其家庭和社会带来了生理、心理上的巨大痛苦和沉重的经济负担。与发达国家一样,发展中国家的人口老龄化亦在迅速推进,因此,以迟发性睾丸功能减退(Late onset hypogonadism in males,LOH)为代表的老龄男子的健康和生活质量问题已引起全社会,尤其是医学界的广泛关注[3]。研究表明,老年男性睾丸Leydig细胞数量的减少和分泌功能的减退是导致老年男性部分雄激素缺乏的主要原因。因此,针对Leydig细胞的起源、分化发育机制的研究已经成为当今国际男性生殖健康研究领域的前沿和热点。对Leydig细胞发育生物学的进一步全面了解将有助于我们更加深入地理解和认识男性生殖器官的分化发育、男性生殖生理以及雄激素紊乱相关疾病的发病机制,为临床预防和治疗此类疾病提供理论基础和开辟新的途径。本文从以下两方面综述该领域的研究进展。
1 Leydig细胞分化发育模式
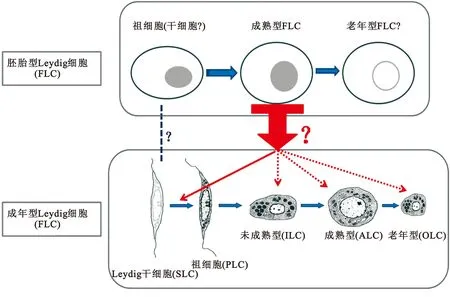
图1 Leydig细胞系发育模式图
Figure 1 Developmental modes of Leydig cell lineage
根据目前的研究结果,多数学者认为Leydig细胞系在个体发育过程中有两种不同的类型: 胚胎型Leydig细胞(Fetal Leydig cell,FLC)和成年型Leydig细胞(Adult Leydig cell, ALC)。胚胎型Leydig细胞的主要功能是分泌睾酮,促进胚胎期胎儿的性别分化和泌尿生殖器官的发育,在出生后睾丸中的FLC开始减少并逐渐消亡[4,5]。成年型Leydig细胞的主要功能是合成和分泌睾酮,其在促进男性第二性征的发育和维持、精子发生、激发性欲、维持性功能以及促进人体的新陈代谢等方面起着重要的作用[6]。其中,关于FLC与ALC的关系问题存在不同的观点:一方面,部分学者认为FLC在个体出生后逐渐消亡,与ALC是两种完全不同的细胞类型,两者之间没有相互影响;另有学者认为,FLC在个体出生后并没有完全消亡,而是以某种形式沉默或通过某种方式转分化为ALC细胞系[7-10]。
1.1 胚胎型Leydig 细胞(Fetal Leydig cell,FLC) 小鼠FLC 在胚胎第12.5天左右即开始在睾丸间质中形成,大鼠大约在第14.5 天,而人类大约在第7~8周。当FLC分化完全后,FLC的形状逐渐由纺锤梭状形向椭圆形转变,并且获得合成类固醇激素的能力。与成年型Leydig细胞一样,FLC具有丰富的滑面内质网和脂滴,唯一不同的是的其高尔基体发育不完全,细胞膜表面有非常多的突起。此外,FLC易成簇,周围包裹有胶原和层粘连蛋白;出生后,成簇的FLC逐渐开始分解[11,15]。
有关FLC的起源现在仍有很多争论,大体上认为FLC可能有4种起源即:肾上腺-性腺原基(中胚层),中胚层神经嵴,中肾,体腔上皮。FLC分化完成后,其数目急剧增加。以大鼠为例,在胚胎第17 天时,每个睾丸FLC的平均数目是2.5×104个,而在第21 天时则激增至1×105个[12]。在FLC产生的一系列分子中,雄激素和胰岛素样因子3(INSL3)至关重要。Alfred Jost首先发现了睾丸起源的雄激素对午非氏管发育的重要性。他利用兔胚胎证实,去势可以导致午非氏管退化,但这种退化可以被睾酮所抑制。FLC可以产生胰岛素样因子3(insulin-like factor 3, INSL3),后者可与其受体相结合,是促使睾丸降入阴囊的关键因子;在哺乳动物中,精子生成所需要的温度相对较低,因此,睾丸下降进入阴囊对于精子的生成至关重要[16-21]。
1.2 成年型Leydig细胞(Adult Leydig cell,ALC) 目前的研究认为,成年型Leydig细胞(Adult Leydig cell,ALC)是由Leydig干细胞(stem Leydig cell,SLC)分化而来;ALC的分化包括4个阶段,即Leydig干细胞、Leydig祖细胞(Progenitor Leydig cell,PLC)、 未成熟型Leydig细胞(immature Leydig cells,ILC)以及成熟型Leydig细胞(ALC)[1]。在Leydig细胞系分化发育过程中,LH 的结合能力、滑面内质网的含量以及睾酮的产生能力均逐渐增加。SLC 在出生时就已经存在于睾丸的间质中,在出生后不久开始定向分化成Leydig细胞系并且进一步增殖,最终分化为成熟的ALC。由于缺乏特异性的细胞表面标记,对SLC的分离鉴定方面的研究进展缓慢。许多候选的SLC表面分子标记物都是一些激素的受体,包括白血病抑制因子(Leukemia inhibitory factor,LIF),血小板衍生生长因子α(Platelet-derived growth factor receptor-α,PDGFRα)以及黄体生成素受体(LHR)。假定的Leydig干细胞应该具有如下特征:具有自我更新能力,能够在体外扩增而不分化;具有分化潜能,能够表达类固醇合成相关的酶类,例如3β-HSD等,最终可以合成睾酮;当被移植到受体的睾丸间质中时,可以存活并且分化成Leydig细胞系。有研究者在大鼠出生后第7 天的睾丸中分离获得了一些具有上述特征的细胞,并认为这些细胞就是Leydig干细胞。这些细胞可以在体外培养扩增6个月保持不分化,经诱导后可以表达Leydig细胞的一些分化标志,例如LHR、CYP-11B1、3β-HSD、CYP-17A1等;当把这些细胞移植入缺失Leydig细胞的睾丸中时,其可以存活并且表达3β-HSD等[6]。
2 Leydig细胞发育过程调控
2.1 FLC发育的分子调控 FLC开始出现在胚胎期的睾丸中,所以认为其分化受到性染色体组分或睾丸特殊机制的调控。Burgogne通过带有XX和XY 的杂交鼠胚胎发现,XX和XY都可以形成FLC[16]。Y染色体上的性别决定基因(SRY)在Leydig细胞中不表达,这说明FLC的分化间接受到SRY基因的调控[14-16]。Dhh基因对Leydig细胞分化起着决定性作用, Dhh基因敲除可导致Leydig细胞的减少,从而导致雄激素分泌严重不足,出现隐睾,性器官萎缩等临床疾病;人类的Dhh基因突变也会出现上述变化。Dhh以旁分泌的形式刺激Leydig前体细胞的分化,特别是刺激SF-1和P450scc表达。然而,当Leydig细胞分化完全时,Dhh基因将被抑制。PDGFRα也可刺激Leydig细胞的增殖,当其基因被敲除后也会导致Leydig细胞数目的下降[17-19]。Dhh和PDGFRα两种细胞信号途径呈平行式调控Leydig细胞的分化。胰岛素生长因子1(IGF-1)可以刺激Leydig细胞增殖,引起类固醇产生;IGF-1基因突变可导致Leydig细胞数目的下降,导致类固醇产生的降低。上述发现说明,Leydig细胞分化的调控是一个网状系统,但其具体作用方式尚未阐明。
除了旁分泌的因子以外,多种转录因子也参与了对FLC分化的调控。Pod1(Capsulin/Epicardin/ Tcf21)基因成螺旋-环螺旋状分子,能够促进侧链切割酶(Scc)阳性细胞的增加,但当Pod1基因被敲除后,将导致雄激素分泌不足或缺如。有研究发现,Pod1可以防止FLC分化不成熟,当FLC增加时,控制类固醇的产生。在Arx(X-linked aristaless-related homeobox gene)基因突变的小鼠中发现FLC分化障碍,在小鼠中Arx不在FLC中表达,这说明Arx间接调控FLC的分化。FLC中也有雌激素受体(Estrogen receptor,ER)的表达,当ER失活时,FLC出现肥大,并可产生大量的睾酮,这表明外源性雌激素可以通过雌激素受体来调控FLC产生类固醇[20]。但是,这些转录因子之间究竟是如何相互作用以及它们与旁分泌因子之间的关系和机制仍不清楚。与成年型Leydig细胞不同的是,FLC的发育不直接受到促黄体生成激素(LH)的调控,因为直到其分化完成后48~72 hFLC才表达LH受体,同时也是在FLC分化完成后72 h以后才能在血清中检测到LH,而ALC则受LH的调控;而且在LH及LH受体敲除的小鼠中FLC的分化正常。
2.2 ALC发育特征及其分子调控 在成年睾丸组织中,Leydig干细胞始终存在,可随时分化为成熟的ALC;以大鼠为例,在出生后第11 ~ 28 天之间,Leydig干细胞开始进入Leydig细胞系的分化状态,形成PLC并开始表达类固醇生成酶系和LH受体。Leydig祖细胞个体较小,外观呈纺锤梭状形,PLC与SLC比较不同的是,PLC可表达LH受体和类固醇生成酶系(如3β-HSD);但是PLC 表达类固醇生成酶的能力比较低。在PLC中,还可见到PDGFRα、LIFR和c-kit等的表达,这表明Leydig祖细胞还保留有一些Leydig干细胞的特征。Leydig祖细胞逐渐变大变圆,增殖活性逐渐降低,开始退出细胞周期;并逐步表达P450scc、3β-HSD和P450c17,PLC基本上不表达或极少量表达17β-HSD3。PLC可以继续分化生成为未成熟的Leydig细胞(ILC)。ILC在大鼠出生后28~56 天开始出现,并大量表达3β-HSD和LH受体。ILC内含有大量的滑面内质网和脂滴,这些脂滴在分化成ALC 后逐渐消失。在ILC时段,睾酮生成酶P450scc、3β-HSD和P450c17活性大大增加,但是17β-HSD3在第56 天后才开始表达。ILC在出生后56 天分化成为ALC,后者含有丰富的滑面内质网、很少的脂滴和全部的类固醇生成酶。ALC 主要功能是分泌睾酮,而ILC的主要功能则是分泌3α-Diol。ALC中5α-还原酶显著降低。ALC是成熟睾丸中的主要细胞群体,但其前体细胞ILC 始终存在于成熟睾丸内部[1,22]。
多种调控因子密切地调控Leydig细胞和类固醇的生成。促性腺激素(LH)是调控Leydig细胞分化最主要因子,但是在早期SLC阶段,LH受体并不表达,所以还有其他因子参与调控。白血病抑制因子(Leukemia inhibitory factor,LIF)是白介素-6(IL-6)家族成员,其受体由其特异结合亚基gp190和转膜信号传导亚基gp130组成[23]。LIF是胚胎干细胞保持其多能性的一个关键因子。在大鼠睾丸中,LIF在胚胎期第13.5 天就可被检测到,它主要在围绕生精小管周围的管周细胞中表达。分化后的Leydig细胞中LIF的表达升高,则会导致类固醇生成降低,主要原因是LIF减少P450scc酶的底物产生。LIF可促进胚胎Leydig细胞的增殖,与LIF相关性很大的因子IL-6在不成熟睾丸中也有表达。IL-6受体主要在PLC和ILC 中表达[24]。因此LIF和IL-6都可以调控胚胎型Leydig细胞和早期Leydig细胞。血小板衍生因子(Platelet derived growth factor, PDGFs)是一个二硫键连接的同源二聚体蛋白家族,其主要功能是在胚胎早期对未分化细胞促进有丝分裂作用,其也在组织重塑中起作用[25]。PDGF受体α对FLC的分化起着决定性作用,同时也可促进ALC的分化。在PDGFAA配体敲除的小鼠中,ALC 不能分化。在胚胎期PDGF的表达先于LHR,因此,PDGF受体α信号途径可能是促进胚胎型Leydig细胞向ALC分化的最早因子。Kit受体在A1型精原细胞和Leydig细胞中表达。Kit在小鼠出生后7 天就可被检测到。与ILC和ALC相比,Kit在PLC中表达量最高。除Sertoli细胞外,SLC也产生Kit配体。Kit配体的主要功能是促进生殖细胞的增殖和存活。由此推测,Kit配体对Leydig细胞也具有类似的功能。在Kit 敲除的转基因鼠中发现,在成年期睾丸中Leydig细胞增生和睾酮生成下降,这预示着Kit调控Leydig细胞增殖[26,27]。
促黄体生成激素(LH)是Leydig细胞合成睾酮至关重要的调节因子。LH与LH受体结合,激活cAMP信号级联效应,动员胆固醇,提高类固醇酶活性。LH信号中断,则可导致类固醇酶活性降低,类固醇生成细胞数目减少并萎缩[28,29]。在GnRHhpg转基因鼠中,LH缺陷,Leydig细胞的数目明显下降,与阳性对照组相比,只有对照组的10%。在体外大鼠ILC培养中,LH可以促进ILC的DNA合成,但这种增加是有限的,但当加入IGF-1因子时,其作用被显著增强。上述结果说明,在LH促进Leydig细胞增殖过程中需要其他的因子参与。胰岛素样生长因子1(Insulin-like growth factor 1, IGF-1)的mRNA及其蛋白和受体都存在于Leydig细胞中,在出生后第4 天的大鼠睾丸中就可以检测到由IGF-1、LH和hCG刺激所导致的IGF-1分泌,并可上调IGF-1受体的表达。IGF-1可刺激Leydig细胞的增殖,并且当与LH协同作用时,这种增殖作用可以被扩大。IGF-1与LH共同作用可以促进Leydig细胞的分化和成熟。在体外实验中,IGF-1可以刺激Leydig细胞的成熟,提高类固醇酶的活性,进而促进睾酮的产生[31]。在IGF-1基因敲除的小鼠中,睾酮的水平严重不足,睾丸变小,Leydig细胞的数目降低。卵泡刺激素(Follicle stimulating hormone, FSH)可提高Sertoli细胞的功能,反过来也可间接地作用于Leydig细胞。目前对FSH 与Leydig细胞发育影响的具体机制还不清。有研究发现,给LH分泌不足的小鼠注射FSH可以促进Leydig细胞的分化和提高类固醇生成活性。但在LH分泌的正常小鼠中,,FSH的作用也不十分显著。FSH基因突变后,Leydig细胞的数目并没有太大的改变[30]。
雄激素受体存在于Leydig细胞所有阶段,但在PLC和ILC中雄激素受体呈高表达;雄激素受体的存在说明雄激素直接参与调控Leydig细胞的发育和功能。当把PLC与LH和双氢睾酮共培养3 天时,PLC产生睾酮水平提高10倍。LH 受体在PLC中含量很少,这说明雄激素可以提高PLC对LH的反应强度。雄激素可以促进PLC中LH受体、雄激素受体和3β-HSD的蛋白活性的增加;雄激素受体发生突变时可以导致Leydig细胞数目的降低和Leydig细胞分化不完全[32,33]。
3 小结
目前,在Leydig细胞发育生物学方面仍然存在许多亟待阐明的问题,主要集中于:①胚胎型Leydig细胞和成年型Leydig细胞之间有无发育上的联系。②Leydig细胞系从原始细胞阶段向成熟细胞阶段分化发育的调控机制。③Leydig干细胞的起源(其定位及其与管周间叶细胞的关系等)。④FLC与ALC是否来源于同一干细胞系等。对Leydig细胞发育生物学的进一步研究将有助于更加深入地理解和认识雄激素及其靶器官的生理病理机制,为临床预防和治疗雄激素相关疾病提供新的思路。
[1]Haolin Chen, Renshan Ge, Barry R.Zirkin. Leydig cells: from stem cells to aging[J]. Mol Cell Endocrinology 2009,306:9-16.
[2]Ge R, Chen G, Hardy MP. The role of the Leydig cell in spermatogenic function[J]. Adv Exp Med Biol. 2008,636:255-269.
[3]Lunenfeld B, Saad F, Hosel CE. ISA, ISSAM and EAU recommend-dations for the investigation, treatment and monitoring of late-onset hypogonadism in males: Scientific background and rationale[J]. Aging Male. 2005,8:59-74.
[4]S.L. Griswold, R.R. Behringer. Fetal Leydig cell origin and development[J]. Sexual Development, 2009,3:1-15.
[5]V.Rouiller-Fabre, C.Levacher, R.Habert. Development of the fetal and neonatal testis[J]. Andrologia, 2003,35:79-83.
[6]Ren-Shan Ge, Qiang Dong, Matthew P. Hardy. In search of rat stem Leydig cells: Identi- fication, isolation, and lineage-specific development[J]. PNAS,2006,103:2719-2724.
[7]Xiufeng Wu, Shengqin Wan, Mary M.Lee. Key factors in the regulation of fetal and postnatal Leydig cell development[J]. J Cell Physiology, 2007,213:429-433.
[8]Griffin DK, Ellis PJ, Dunmore B, Bauer J, Abel MH, Affara NA. Transcriptional Profiling of Luteinizing Hormone Receptor-Deficient Mice Before and after Testosterone Treatment Provides Insight into the Hormonal Control of Postnatal Testicular Development and Leydig Cell Differentiation[J]. Biol Reprod, 2010,82(6):1139-1150.
[9]Lei Dong, Scott A. Jelinsky, Ren-shan Ge. Gene expression during development of fetal and adult Leydig Cells[J]. Ann. N.Y. Acad. Sci, 2007,1120: 16-35.
[10] Barsoum IB, Yao HH. Fetal Leydig cells: progenitor cell maintenance and differentiation[J]. J Androl, 2010,31(1):11-15.
[11] Kerr JB. and C.M. Knell, The fate of fetal Leydig cells during the development of the fetal and postnatal rat testis[J]. Development, 1988,103(3): 535-544.
[12] Kuopio T, Tapanainen J, Pelliniemi LJ,etal. Developmental stages of fetal-type Leydig cells in prepubertal rats[J]. Development, 1989, 107(2): 213-220.
[13] Brennan J, C. Tilmann, and B. Capel, Pdgfr-alpha mediates testis cord organization and fetal Leydig cell development in the XY gonad[J]. Genes Dev, 2003, 17(6): 800-810.
[14] Nishino K, Yamanouchi K, Naito K,etal. Characterization of mesonephric cells that migrate into the XY gonad during testis differentiation[J]. Exp Cell Res, 2001, 267(2): 225-232.
[15] Schmahl J, Eicher EM, Washburn LL,etal., Sry induces cell proliferation in the mouse gonad[J]. Development, 2000, 127(1): 65-73.
[16] Burgoyne PS, Buehr M, Koopman P,etal. Cell-autonomous action of the testis-determining gene: Sertoli cells are exclusively XY in XX----XY chimaeric mouse testes[J]. Development, 1988,102(2): 443-450.
[17] Clark AM, KK Garland, and L.D. Russell, Desert hedgehog (Dhh) gene is required in the mouse testis for formation of adult-type Leydig cells and normal development of peritubular cells and seminiferous tubules[J]. Biol Reprod, 2000, 63(6): 1825-1838.
[18] Yao HH, W. Whoriskey, and B. Capel, Desert Hedgehog/Patched 1 signaling specifies fetal Leydig cell fate in testis organogenesis[J]. Genes Dev, 2002, 16(11): 1433-1440.
[19] Canto P, Söderlund D, Reyes E,etal. Mutations in the desert hedgehog (DHH) gene in patients with 46,XY complete pure gonadal dysgenesis[J]. J Clin Endocrinol Metab, 2004, 89(9): 4480-4483.
[20] Delbes G, Levacher C, Duquenne C,etal., Endogenous estrogens inhibit mouse fetal Leydig cell development via estrogen receptor alpha[J]. Endocrinology, 2005,146(5): 2454-2461.
[21] Kumagai J, Hsu SY, Matsumi H,etal. INSL3/Leydig insulin-like peptide activates the LGR8 receptor important in testis descent[J]. J Biol Chem, 2002,277(35): 31283-31286.
[22] Mendis-Handagama, SM, and H.B. Ariyaratne, Differentiation of the adult Leydig cell population in the postnatal testis[J]. Biol Reprod, 2001,65(3): 660-671.
[23] Malaval L, Liu F, Vernallis AB,etal. GP130/OSMR is the only LIF/IL-6 family receptor complex to promote osteoblast differentiation of calvaria progenitors[J]. J Cell Physiol, 2005, 204(2): 585-593.
[24] Piquet-Pellorce, C, Dorval-Coiffec I, Pham MD,etal. Leukemia inhibitory factor expression and regulation within the testis[J]. Endocrinology, 2000, 141(3): 1136-141.
[25] Hoch, RV. and P. Soriano, Roles of PDGF in animal development[J]. Development, 2003,130(20): 4769-4784.
[26] Ge RS, Dong Q, Sottas CM,etal. Gene expression in rat leydig cells during development from the progenitor to adult stage: a cluster analysis[J]. Biol Reprod, 2005,72(6): 1405-1415.
[27] Kissel H, Timokhina I, Hardy MP,etal. Point mutation in kit receptor tyrosine kinase reveals essential roles for kit signaling in spermatogenesis and oogenesis without affecting other kit responses[J]. EMBO J, 2000,19(6): 1312-1326.
[28] Lei ZM, Zou W, Mishra S,etal. Epididymal phenotype in luteinizing hormone receptor knockout animals and its response to testosterone replacement therapy[J]. Biol Reprod, 2003, 68(3): 888-895.
[29] Scott IS, Charlton HM, Cox BS,etal. Effect of LH injections on testicular steroidogenesis, cholesterol side-chain cleavage P450 mRNA content and Leydig cell morphology in hypogonadal mice[J]. J Endocrinol, 1990,125(1): 131-138.
[30] Baker PJ, Pakarinen P, Huhtaniemi IT,etal. Failure of normal Leydig cell development in follicle-stimulating hormone (FSH) receptor-deficient mice, but not FSHbeta-deficient mice: role for constitutive FSH receptor activity[J]. Endocrinology, 2003,144(1):138-145.
[31] Gelber SJ, Hardy MP, Mendis-Handagama SM,etal. Effects of insulin-like growth factor-I on androgen production by highly purified pubertal and adult rat Leydig cells[J]. J Androl, 1992, 13(2): 125-130.
[32] Shan L, Hardy DO, Catterall JF,etal. Effects of luteinizing hormone (LH) and androgen on steady state levels of messenger ribonucleic acid for LH receptors, androgen receptors, and steroidogenic enzymes in rat Leydig cell progenitors in vivo[J]. Endocrinology, 1995, 136(4): 1686-1693.
[33] Murphy L, IA Jeffcoate, and P.J. O′Shaughnessy, Abnormal Leydig cell development at puberty in the androgen-resistant Tfm mouse[J]. Endocrinology, 1994,135(4): 1372-1377.
Advances in research of developmental biology of Leydig Cells
MA Xue1reviewingDONG Qiang2checking
(1.DepartmentofPediatricUrology,WestChinaHospital,SichuanUniversity,Chengdu610041,China;2.DepartmentofUrology,WestChinaHospital,SichuanUniversity,Chengdu610041,China)
Leydig cells are endocrine cell located in a mammalian testicular interstitial that play important roles in promoting genital differentiation of embryonic development, the development and maintenance of male secondary sex characteristics, spermatogenesis, maintain sexual function and the body′s metabolism. Therefore, the origin, mechanism of differentiation and development of Leydig cells has become the highlights of male reproductive health research. Progress in the field has been discussed in this review.
Leydig cell; Testosterone; Developmental biology
四川省科技厅支撑计划(2014SZ0031);四川省西部精神医学协会科研基金(Wcpafund2014-10)
董强,教授,本刊常务编委,E-mail:dqiang@gmail.com.
R
A
10.3969/j.issn.1672-3511.2016.02.043
2015-10-08; 编辑: 张文秀)
