盐胁迫下G蛋白在拟南芥种子萌发和根生长发育中的作用
2016-04-26王绍杰周
王绍杰周 索
(1.河南天冠企业集团有限公司,河南 南阳 473000;2.南阳师范学院,河南 南阳 473061)
盐胁迫下G蛋白在拟南芥种子萌发和根生长发育中的作用
王绍杰1周 索2
(1.河南天冠企业集团有限公司,河南 南阳 473000;2.南阳师范学院,河南 南阳 473061)
以拟南芥的野生型(WS)、异三聚体G蛋白α亚基基因GPA1缺失突变体(gpα1-1,gpα1-2)和超表达突变体(wGα,cGα)的种子为材料,在添加不同浓度NaCl盐溶液的MS基本培养基上培养,观察种子的萌发及根生长情况。结果表明:随着盐胁迫程度的增加,不同基因型拟南芥种子萌发率、存活率、主根长度及侧根数目均明显呈下降趋势;在同一浓度下,缺失型突变体的萌发率和存活率高于超表达突变体和野生型;缺失型突变体的主根长度和侧根的数目也大于野生型和超表达突变体。初步说明了植物异三聚体G蛋白α亚基在拟南芥种子萌发及根生长发育过程中起负调控作用。
拟南芥;盐胁迫;种子萌发;异三聚体G蛋白
拟南芥(Arabidopsis thaliana),因其基因组简单,突变体众多,生长发育指标资料全面等特点而被广泛使用,成为一种典型的模式植物。NaCl对拟南芥种子萌发、存活及根生长有抑制作用[1]。但NaCl抑制过程中的信号转导途径目前还不清楚。
高等真核生物中,异三聚体G蛋白(简称G蛋白)参与的跨膜信号转导途径是一种常见且保守的作用机制,胞外信号通过与质膜相结合的受体和质膜内表面的异三聚体G蛋白,将信号传递给效应器并产生胞内第二信使,再由后者调节下游效应器及酶活性。大量的药理学及分子细胞生物学证据表明,植物异三聚体G蛋白参与了诸多发育过程,如气孔开关、细胞周期调节、根的发育等,并在植物感知环境信号反应中起着重要的作用。异三聚体G蛋白由α、β、γ3个亚基组成,目前认为G蛋白α亚基(Gα)在信号转导过程中发挥主导作用。目前,国际上通用的研究G蛋白α亚基功能的材料是拟南芥G蛋白α亚基基因GPA1缺失突变体(gpα1-1,gpα1-2)和超表达突变体(wGα,cGα)。
为了探索异三聚体G蛋白α亚基在盐胁迫下对拟南芥种子萌发及根生长发育过程中的调控作用,本试验以拟南芥野生型(WS)、超表达突变体(wGα,cGα)、缺失突变体(gpα1-1,gpα1-2)5种基因型拟南芥种子的萌发率、存活率、主根长度、侧根数目等指标进行测量,以期为G蛋白α亚基是否参与盐胁迫下种子萌发、根生长发育调节提供科学的依据。
1 材料与方法
1.1 材料
拟南芥(Arabidopsis thaliana)的野生型(WS)和异三聚体G蛋白α亚基基因GPA1缺失突变体(gpα1-1,gp1-2)和超表达突变体(wGα,cGα)5种基因型种子。
1.2 试验方法
1.2.1 培养基配制和分装。培养无菌拟南芥种子的培养基为MS基本培养基,添加一定量的NaCl,使最终浓度分别达到0、40、80、120、160 mmol/L和200 mmol/L(经预备试验反复证明当NaCl浓度大于200 mmol/L时,5种基因型都不萌发),每配方6皿,共需30个培养皿。按常规先将配制好的培养基放入锥形瓶中,每瓶100 mL共5瓶,2种超表达突变体基因型的培养基必须准确每瓶25 mL(培养基要加激素100 μL/L)共10瓶,高压灭菌(121℃,20 min),将每个100 mL锥形瓶分装为4个9.0 cm直径的培养皿(培养皿也经高压灭菌),做好标记,放入超净工作台中,以便接种。
1.2.2 种子的消毒和接种。拟南芥种子经表面(75%乙醇,30 s)和深层(HgCl2,4 min,无菌水洗3次)消毒后,用移液枪将拟南芥种子点种于相应的培养皿上,在培养室(22℃、光强6 000~8 000 Lux、16 h/8 h光暗周期,相对湿度70%)培养,获得无菌苗,每天观察萌发情况,10 d后统计萌发数,存活率,测量主根长度和侧根数目3次,取平均值。
1.2.3 拟南芥种子萌发率的测定。待种子长出芽时,对每个培养皿中拟南芥种子的萌发数目进行统计并算出萌发率。萌发率=萌发种子数/接种种子总数。
1.2.4 拟南芥种子存活率的测定。待4周幼苗长出三四片叶子后,统计各培养皿中正常生长的幼苗,并算出拟南芥的存活率。存活率=正常生长幼苗数目/接种种子总数。
1.2.5 拟南芥主根长度和侧根数目测量。待4周后,取6张足够大的黑纸,放在灯光下。取同一盐浓度处理下的不同基因型拟南芥幼苗各45株,将其平行摆放在黑纸上,测量主根长度及观察侧根数目,并记录主根长度和侧根的数目。
2 结果与分析
2.1 盐胁迫下5种基因型拟南芥种子的萌发率
由表1、图1可以看出,盐胁迫对拟南芥种子的萌发率具有浓度依赖。5种基因型的拟南芥种子萌发率随着盐浓度的增加而呈下降趋势,160~200 mmol/L盐胁迫条件对种子萌发有较大的影响,种子几乎不萌发,表明随着盐浓度的增加,各基因型拟南芥种子的萌发受到的抑制增大;同一浓度下不同基因型拟南芥的萌发率相比,缺失型突变体的萌发率大于超表达突变体和野生型,表明缺失型突变体拟南芥种子萌发受到的抑制小于超表达突变体和野生型。
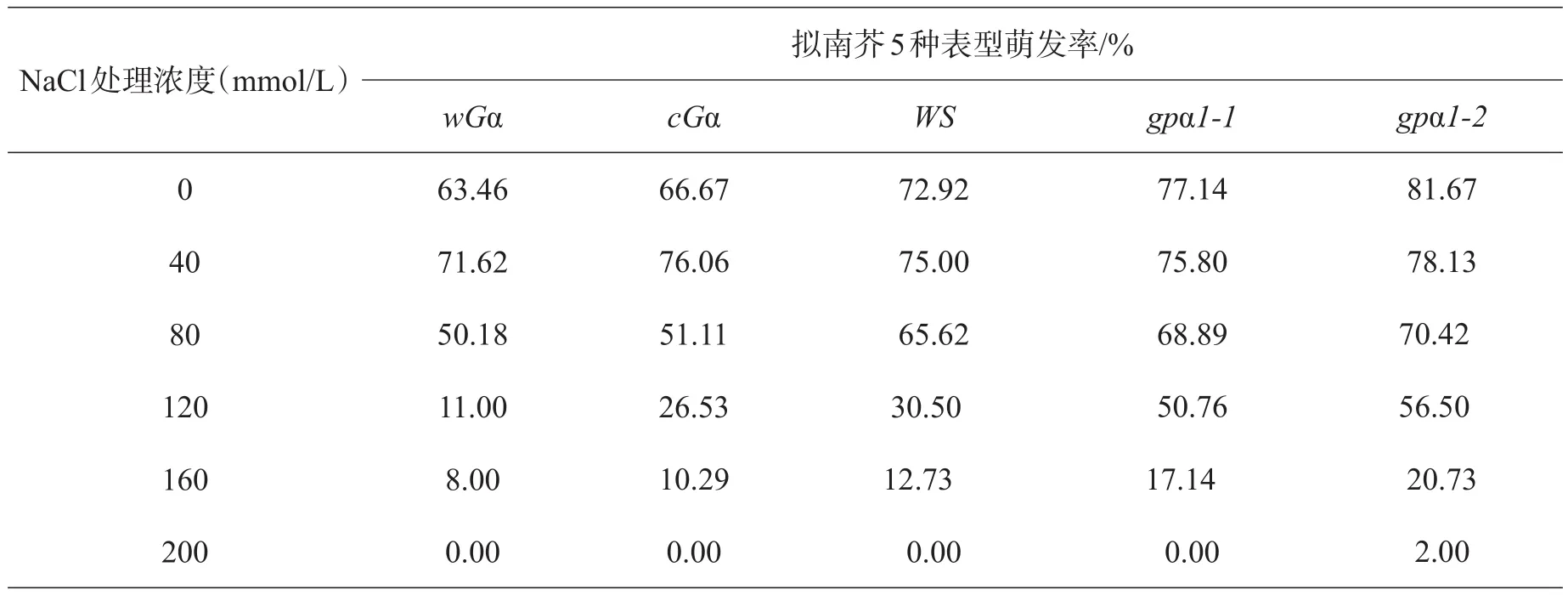
表1 NaCl对不同基因型拟南芥种子萌发率的影响
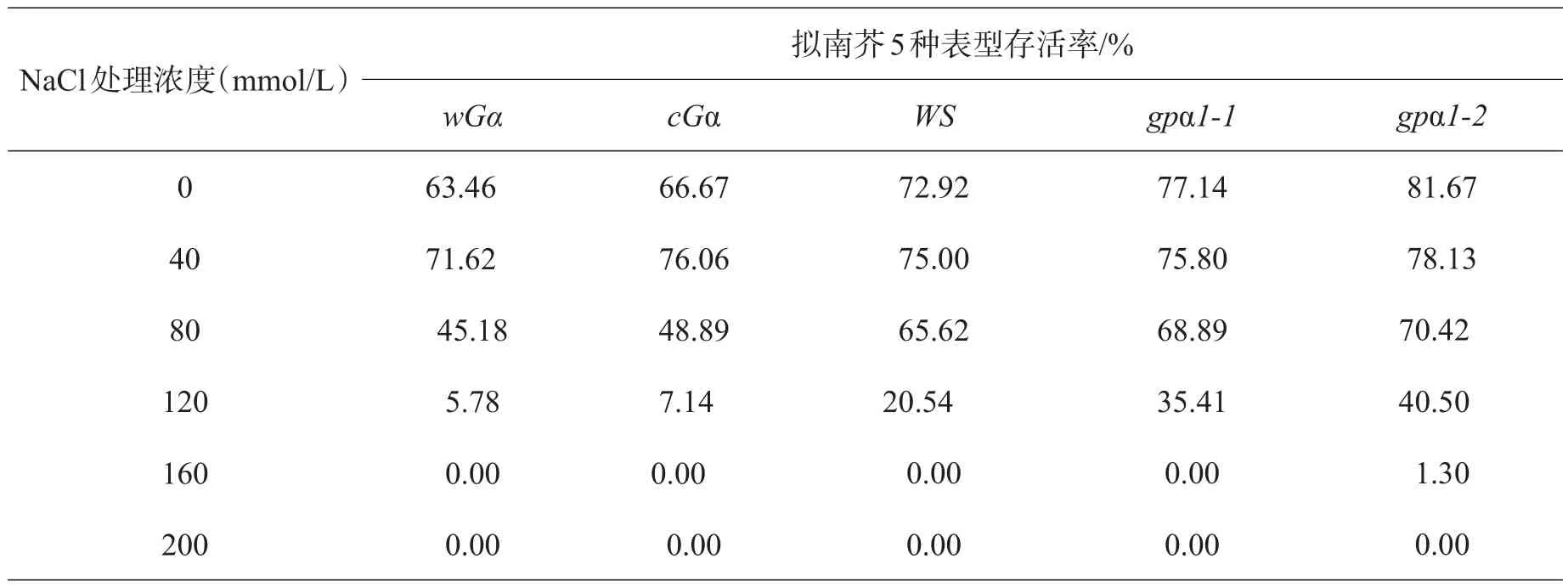
表2 NaCl对不同基因型拟南芥种子存活率的影响
图1 NaCl对不同基因型拟南芥种子萌发率的影响
2.2 盐胁迫下5种基因型拟南芥幼苗的存活率盐胁迫对不同基因型拟南芥种子的存活都有抑制作用,第5天40~120 mmol/L盐胁迫的拟南芥都长出叶子,而160 mmol/L盐胁迫的拟南芥萌发的种子长出子叶后就逐渐枯萎,不能进一步生长真叶。将能够生长出真叶的幼苗作为能够存活的依据,统计后发现,盐胁迫培养皿中,盐浓度超过160 mmol/L的培养基中,拟南芥种子萌发出子叶后,随着时间的延长,最终均陆续死亡。由表2、图2可知,5种基因型的拟南芥随着盐浓度的升高,其存活率呈下降趋势,受到的抑制逐渐增大,同一盐浓度下缺失型突变体的存活率大于超表达突变体和野生型,表明缺失型突变体受到的抑制低于超表达突变体和野生型。

图2 NaCl对不同基因型拟南芥种子存活率的影响
2.3 盐胁迫下5种基因型拟南芥幼苗的主根长度
种子萌发到第10天,测量不同盐浓度培养下的5种基因型拟南芥幼苗的主根长度。由表3、图3可以看出,在盐胁迫下,随着NaCl浓度的增加,拟南芥幼苗的主根长度明显降低,同一浓度下缺失型突变体的主根长度大于超表达突变体和野生型,野生型拟南芥的根也稍长于超表达型,说明在同一浓度下,不同基因型拟南芥受到的抑制情况为超缺失型<野生型<表达型。
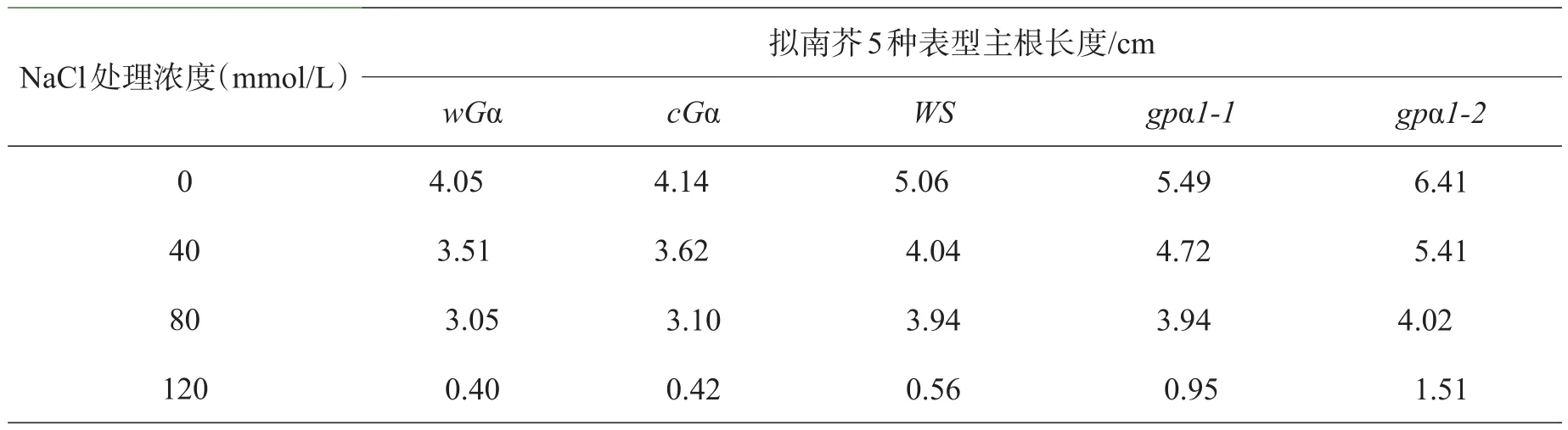
表3 NaCl对不同基因型拟南芥主根长度的影响
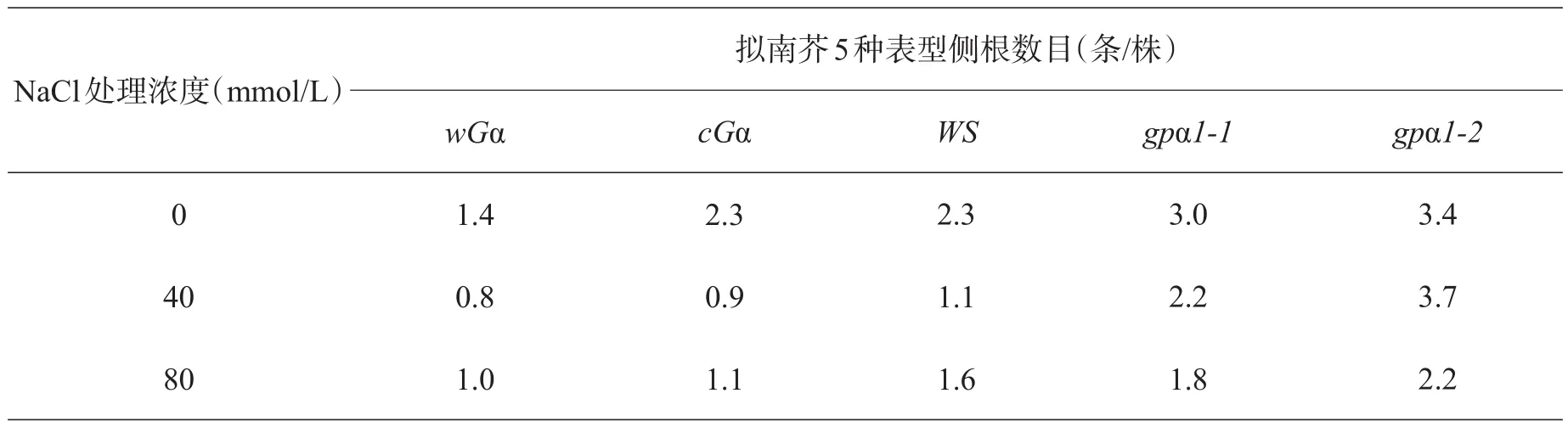
表4 NaCl对不同基因型拟南芥侧根数目的影响
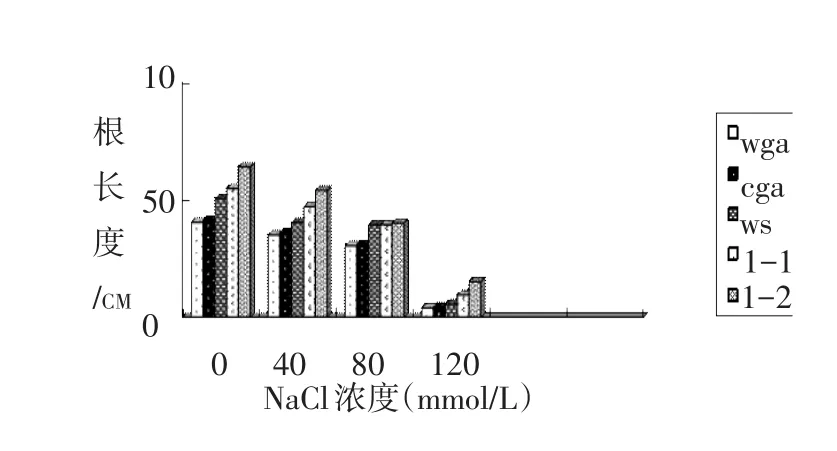
图3 NaCl对不同基因型拟南芥主根长度的影响
2.4 盐胁迫下5种基因型拟南芥的侧根数目

图4 NaCl对不同基因型拟南芥侧根数目的影响
由表4、图4可知,在盐胁迫下,随着NaCl浓度的增加,拟南芥幼苗的侧根数目明显减少,同一浓度下缺失型拟南芥的侧根数目要大于野生型和超表达型,而野生型的也稍多于超表达型拟南芥,说明他们受到的抑制情况为:超表达型>野生型>缺失型。
3 讨论与结论
作为细胞质膜上重要的跨膜信号转导分子,异三聚体G蛋白在植物发育过程中发挥重要作用,已知G蛋白参与种子萌发[2]、光控发育[3]以及包括生长素、赤霉素和脱落酸在内的多种植物激素[4]对植物生长发育的调控。周索等[5,6]曾指出异三聚体G蛋白参与生长素调控的拟南芥侧根生长发育过程。Ullah等[7]曾指出,G蛋白α亚基参与促进叶和茎细胞的细胞分裂,其基因缺失后导致有丝分裂频率降低,超表达突变体细胞分裂速度增加。Okamoto等[3]曾报道G蛋白α亚基超表达可以明显增强光照对植物生长的抑制,推测Gα亚基缺失将会导致这种抑制作用的减弱,而使生长加速。Tanaka等[4]报道G蛋白参与了赤霉素信号转导,并可以增强生长对于赤霉素的敏感性。杨振等[8]发现Gα亚基参与了拟南芥器官(植株高度/叶片形状和长角果)的伸长生长过程,并证实Gα亚基起负调控作用。
本研究结果显示,随NaCl胁迫程度的增加不同基因型拟南芥种子萌发率、存活率、主根长度和侧根数目均明显呈下降趋势。在同一浓度下,缺失型突变体的萌发率和存活率高于超表达突变体和野生型,野生型又稍大于超表达型;缺失型突变体的主根长度和侧根的数目,也大于野生型和超表达突变体,野生型又稍多于超表达型。G蛋白α亚基缺失之后,G蛋白表达减弱,调控作用被减弱,因此导致缺失突变体萌发率,存活率比野生型和超表达突变体高,根长度较他们长,侧根数目较他们多。表明异三聚体G蛋白参与了盐胁迫下拟南芥种子萌发及根生长发育的调节。本研究初步证明盐胁迫下G蛋白α亚基在拟南芥种子萌发及根生长发育中起负调控作用。但G蛋白α亚基是否参与其他离子胁迫下拟南芥种子萌发及根生长发育的信号转导途径以及G蛋白α亚基的下游效应器还不清楚,还有待于进一步研究确定。
[1]柳海龙,杜娟,吴立柱,等.拟南芥种子萌发及幼苗生长对干旱和NaCl胁迫的响应[J].河北农业大学学报,2008(2):12-15.
[2]Ullah H,Chen J G,Wang S.Role of the heterotrimeric G-protein in regulation of Arabidopsis seed germination[J].Plant Physiol,2002(2):897-907.
[3]Okamoto H,Matsui M,Deng XW.Over-expression of the heterotrimeric G protein a-subuint enhances phytochrome-mediated inhibition of hypocotyls elongation Arabidopsis[J].Plant Cell,2001(13):639-651.
[4]Tanaka MU,Fujisawa Y,Kobayashi M.Rice dwarf mutant d1,which is defective in the subunit of the heterotrimeric G protein,affects gibberellin signal transduction[J]. Proceedings of the National Academy of Sciences of the United States ofAmerica,2000(21):11638-11643.
[5]周索,杜丽,赵松峰,等.Ca2+参与异三聚体G蛋白调控的拟南芥侧根生长发育过程.中国农业大学学报,2008(5):5-10.
[6]周索,杜丽,尚忠林,等.异三聚体G蛋白在IAA诱导的拟南芥根生长发育中的作用[J].西北农业学报,2007(6):187-191.
[7]Ullah H,Chen JG,Young JC.Modulation of cell proliferation by heterotrime
ric G-protein in Arabidopsis[J].Science,2001(5524):2066-2069.
[8]杨振,刘婷,武延生,等.异三聚体G蛋白在拟南芥生长发育过程中的作用[J].河北师范大学学报,2005(3):299-304.
S188;Q945
B
1674-7909(2016)33-82-4
