蓄水坑灌条件下不同灌水下限幼龄苹果树叶片光合特性研究
2016-03-23郭向红孙西欢马娟娟
李 波,郭向红,孙西欢,2,马娟娟
(1.太原理工大学水利科学与工程学院,太原 030024;2. 山西水利职业技术学院,山西 运城 044004)
苹果是落叶果树的主要栽培品种之一,具有较高的经济价值。黄土高原地区为我国苹果种植优势区,然而本地区干旱少雨,水资源时空分布不均匀,严重制约了果树种植业的发展。蓄水坑灌法是一种适宜在干旱半干旱地区推广的新型节水灌溉技术,它可以有效缓解水分供需矛盾,提高灌溉水利用率[1]。
土壤水分是影响作物生长的主要限制因子之一,它对植物生长和代谢的影响是多方面的,对光合作用的影响尤为突出[2]。光合作用是植物生长重要的生理过程之一,是构成植物生产力的最主要因素,是生态系统中物质循环与能量流动的基础和重要环节[3]。光合作用受外界环境条件和内部因素的限制,是一个复杂的生物物理化学过程。开展植物光合特性有关研究有助于在农业生产过程中采取适当的经营管理措施提高光合能力、增加经济产量和生物固碳量[4]。
目前有关果树光合特性方面的研究已有大量的报道,对苹果树光合作用的研究主要集中在不同品种光合特性的比较[5],不同水肥组合[6]、干旱等逆境胁迫[7-9]对光合特性的影响,其试验材料多是盛年期果树。而在蓄水坑灌领域,幼龄苹果树叶片光合特性尚方面缺乏系统的研究和报道。本文探讨了蓄水坑灌条件下不同灌水下限幼龄苹果树叶片光合特性及净光合速率与影响因子的相关关系,并以地面灌溉为对照,进一步丰富了蓄水坑灌条件下植物生理生化的研究成果,并为蓄水坑灌法的推广运用提供理论依据。
1 材料与方法
1.1 试验区概况
试验在山西省农业科学院果树研究所内进行。山西省农科院果树研究所位于太谷县西南,东经112°32′,北纬37°23′,平均海拔约800 m。试验区属典型的大陆性半干旱气候,多年平均气温为9.8 ℃,多年平均降雨量为462.9 mm,无霜期175 d。试验区土质以壤土为主,土壤体积质量1.47 g/cm3,饱和含水率(体积)49.21%,0~1 m土层平均田间持水率(体积)30%,灌溉水源为井水。试验材料为3 a生矮砧红富士,株距×行距为2 m×2 m,树形整齐,树势旺盛。
1.2 试验设计
为研究蓄水坑灌条件下不同灌水下限水平幼龄苹果树叶片的光合特性,试验以灌水下限为控制因子,取3个水平,分别为田间持水率的50%、60%、70%,并以地面灌溉为对照组,共4组处理,每组处理5次重复。试验分组情况见表1,试验期间各组处理灌水量及灌水日期见表2。
蓄水坑灌法每株果树下均匀布置4个直径为20 cm的蓄水坑,坑深20 cm,蓄水坑中心距果树树干中心40 cm。蓄水坑使用PVC网布作固壁处理,防止坑壁坍塌,坑底铺设土工膜防止深层渗漏。水分监测点布置在相邻两蓄水坑中心与树干中心的连线上,距树干中心的距离分别为30、60和90 cm,地面灌溉水分监测点的布设形式与蓄水坑灌法相同(见图1)。
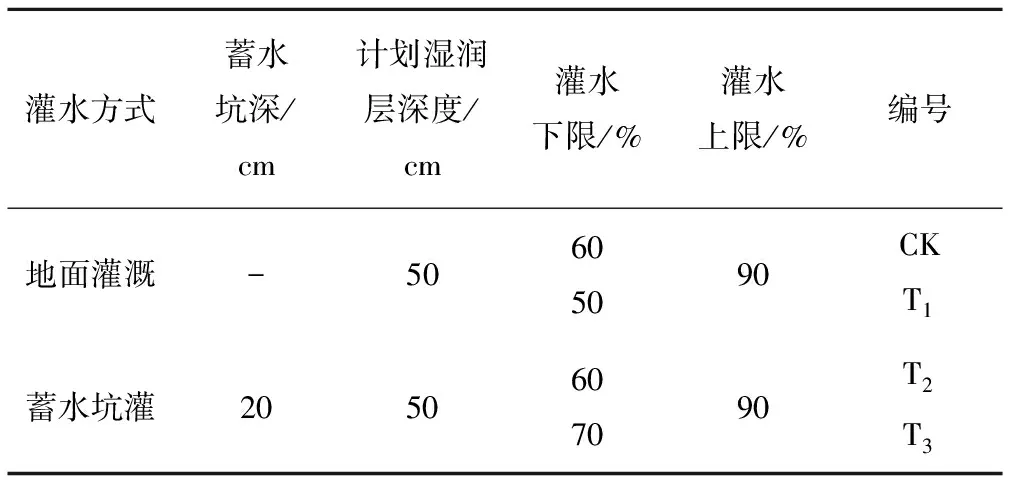
表1 试验设计

表2 灌水量及灌水日期 L
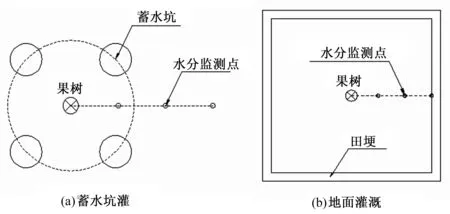
图1 水分监测点及田间工程布置
各小区采用随机排列法排布。相邻小区间设置1棵非实验用树,防止不同实验处理之间的互相干扰。各组处理施肥、修剪、除虫、除草等农艺措施相同。
1.3 测定项目与方法
(1)土壤含水率。采用TRIME-PICO IPH 土壤水分测量系统测定土壤体积含水率,每隔5~7 d测定一次,雨后及灌水后加测。测点测量深度为140 cm,每隔20 cm测定一次。每组处理测量5株果树,并以5株果树计划湿润体(垂向距地表50 cm,径向距树干中心60 cm的圆柱体)内土壤含水率的平均值作为试验处理、数据分析的依据。
(2)叶片光合特性指标。2015年5月20日开始进行试验处理,6月22日开始进行光合指标的测量,9月21日试验结束。选择在天气晴朗,微风无云的典型晴天上午9∶00-11∶00,利用美国LI-COR公司生产的LI-6400XT便携式光合测定仪进行测量。每组处理选择5株果树,在每株果树中上部按东西南北方向选择4片相似且成熟健康的叶子进行测量,每片叶连续采集3个稳定数据取平均值。测定的主要指标包括:叶片净光合速率Pn、胞间CO2浓度Ci、蒸腾速率Tr、气孔导度Cond、光合有效辐射PARi等。每隔15 d左右测定一次。
(3)气象因子。在试验区设立JB-03小型室外气象站,对试验期间的降水量、气温、土壤温度、风速、风向、大气压、相对湿度、太阳辐射等气象因子进行连续监测。
如图2(a)所示,试验期间各月降雨总量分布情况5-7月降雨较少且分布均匀,6月降雨总量仅为15.4 mm,8月降雨总量最大,达95.9 mm,单日最大雨量40.9 mm(8月1日),9月降雨总量76.6 mm,单日最大雨量33.3 mm(9月9日)。试验期间气温变化幅度较大,最低为14.88 ℃(5月21日),最高为28.13 ℃(7月28日)。5月气温较低(平均18.8 ℃),进入6月后(平均21.8 ℃),气温持续上升。7月平均气温23.86℃,为一年中最高水平。7月下旬至8月上旬,连续降雨导致气温持续偏低。由8月中旬至试验结束,气温从平均21.93 ℃(8月)缓慢降至平均17.19 ℃(9月)。
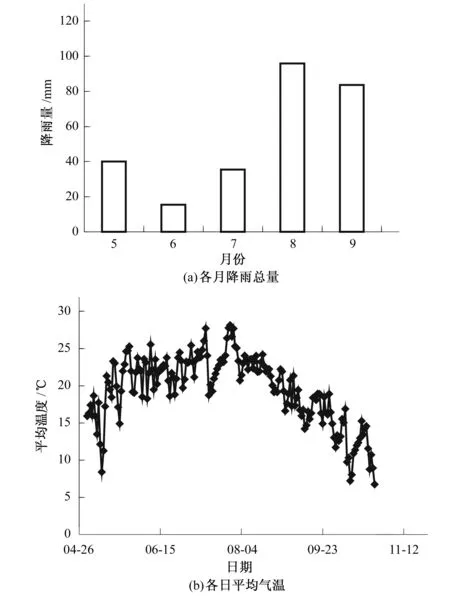
图2 试验期月降雨量与日平均气温变化情况
1.4 数据处理与分析
运用IBM SPSS Statistics 20软件对数据进行相关分析、通径分析和多元逐步回归分析,采用EXCEL 2003软件进行图形处理。
2 结果与分析
2.1 不同灌水下限幼龄苹果树叶片光合特性
2.1.1试验期间土壤含水率及光合有效辐射的变化规律
如图3(a)所示,试验各处理计划湿润层平均含水率变化趋势基本一致,因降雨、灌水等过程升高,随果树生长耗水而降低,在田间持水率的16.3%~27.08%范围内波动。T3处理自7月7日起含水率始终高于其他各组处理,水分充足。T1处理试验期间仅有1次灌水补充,含水率低于其他各组处理。各组处理在7月20日前含水率差异较大,进入8月上旬后,因降雨量增大,土壤水分得到有效补充,处理间差异减小。
各处理光合有效辐射的变化规律基本相同[图3(b)],自6月22日开始呈波动上升的趋势,8月6日略有下降,9月12日达到峰值,9月19日因太阳光照强度减弱而显著下降。因每次测量时各组处理测量时段不可能完全同步,加之野外自然条件复杂多变,使不同处理间出现一定差异。
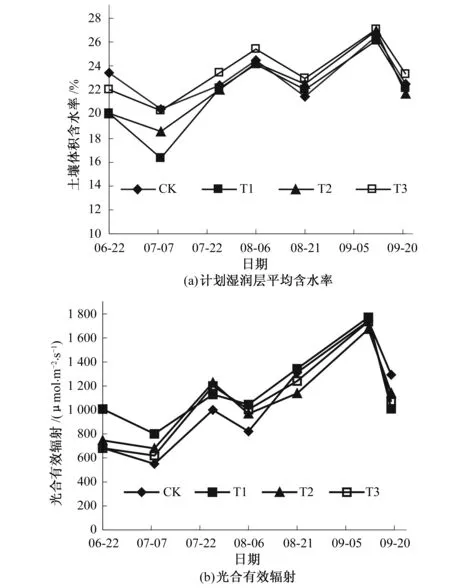
图3 计划湿润层平均含水率及光合有效辐射变化情况
2.1.2不同灌水下限叶片净光合速率的变化规律
净光合速率也称为表观光合速率,是单位时间、单位面积上光合速率与呼吸速率的差值,能够直观反映作物中有机物的累积速率[10]。如图4(a)所示,不同处理幼树叶片净光合速率变化规律基本相同,均随时间推移呈波动上升趋势。7月8日因土壤含水率及光合有效辐射水平降至最低点,各处理净光合速率均为最小值。从8月6日开始,各处理净光合速率处于平台期,随土壤含水率及光合有效辐射水平波动上升而缓慢增大,至9月12日达到峰值后,因土壤含水率及光合有效辐射水平迅速降低而减小。试验期内,各处理净光合速率整体服从T1>T2≥T3>CK的分布规律。T1处理土壤含水率最低,但光有效辐射水平较高,净光合速率一致保持较高水平;T3处理土壤含水率最高,但7月7日净光合速率仅有9.11 μmol/(m2·s),为各处理最小值;9月12日CK处理土壤含水率及光合有效辐射水平均不是最大值,但净光合速率却达到23.31 μmol/(m2·s),比同期最小的T2处理大13.3%。说明光合作用是植物的一种复杂的生理生化反应,土壤水分状况、光合有效辐射、大气温度等因子都对光合作用有一定程度的影响,各因子与净光合速率之间相互作用较为复杂,其具体关系将在下节分析。
2.1.3不同灌水下限叶片气孔导度的变化规律
气孔是叶片和外界环境进行气体交换和水分逸散的门户,气孔的开闭程度对植株的蒸腾和光合作用有重要影响,气孔导度是气孔开闭程度的量化指标[11]。由图4(b)看出,各处理气孔导度的变化规律基本相同。7月7日各处理气孔导度降至最小值,随后迅速增大,CK处理在7月26日达到最大值0.44 mmol/(m2·s),其他处理在8月6日达到峰值,较CK处理的峰值分别增大了16.1%(T1)、10.4%(T2)、6.9%(T3)。随后,因光有效辐射水平增大、温度升高、叶片蒸腾失水等原因,气孔关闭,气孔导度逐渐减小。至9月19日,光合有效辐射水平迅速降低,各处理气孔导度又有不同程度的增大。从8月6日开始,各处理土壤含水率波动上升,且在9月12日达到最大值,但气孔导度却在9月12日降至第2个波谷,此时各处理光合有效辐射水平达到最大值,说明气孔导度对光合有效辐射的敏感程度高于土壤含水率,其变化主要受光合有效辐射的影响。同时注意到,CK处理气孔导度的变化较其他处理超前,说明在光有效辐射基本相同的情况下,地面灌溉条件下气孔导度对土壤水分变化的响应更积极,同时也说明蓄水坑灌具有一定的蓄水保墒作用。
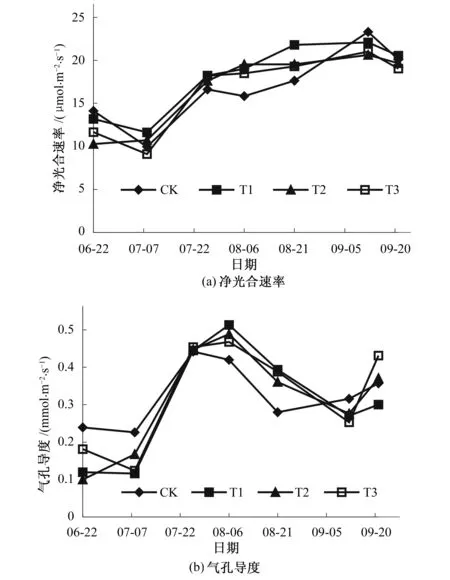
图4 各处理叶片净光合速率及气孔导度变化特征
2.1.4不同灌水下限叶片蒸腾速率的变化规律
蒸腾作用是植株必不可少的代谢过程,它是植物水分吸收和运输的主要动力,也是植物吸收、运输矿物质和有机物的主要动力。另外,蒸腾作用还能降低叶片温度,防止叶片被灼伤[12]。由图5(a)看出,各处理蒸腾速率变化规律基本相同,呈双峰分布,这主要与土壤含水率及光有效辐射的变化有关。7月26日前,光合有效辐射小于1 000~1 200 μmol/(m2·s),各处理蒸腾速率随土壤含水率及光合有效辐射水平的增加基本以直线上升。8月6日,因光合有效辐射水平降低,各处理蒸腾速率出现不同程度的降低,最大降幅超过54%(CK),此时土壤含水率仍处于峰值。至8月21日,土壤含水率下降而光合有效辐射水平上升,各处理蒸腾速率到达第2个峰值,说明蒸腾速率对光合有效辐射的响应较土壤含水率更为积极。当光合有效辐射大于1 000~1 200 μmol/(m2·s)时,各处理蒸腾速率随光合有效辐射的增长而迅速下降,此时因光照过强,植株为保护自己而限制了气孔开度,从而导致了蒸腾速率降低。9月19日光合有效辐射水平降低,气孔开度增大,各处理蒸腾速率增大。试验期内,植株蒸腾速率整体上呈T3>T2≥T1>CK的分布规律,说明在光合有效辐射水平差异不大时,土壤含水率越高,植株蒸腾作用越强烈。自7月26日开始,蓄水坑灌条件下植株蒸腾作用显著大于地面灌溉,说明在水分充足的情况下,蓄水坑灌有效减少了棵间蒸发,提高了灌溉水利用率,这与雷涛等人的研究结果相同[13]。
2.1.5不同灌水下限叶片胞间CO2浓度的变化规律
CO2是植株光合作用的底物,空气中的CO2浓度是限制光合速率的一个重要因素[14]。由图5(b)可见,各处理胞间CO2浓度变化规律基本相同,且与气孔导度的变化规律较为一致,呈单峰分布。8月6日,CK处理胞间CO2浓度达到峰值319.5 μmol/mol,滞后于在7月26日达到峰值的其余各组处理。随后各处理胞间CO2浓度逐渐减小,至9月12日到达波谷,此时各处理净光合速率最大。9月19日因气孔导度增大,叶片净光合速率减小,各处理胞间CO2浓度有不同程度的上升。结合图4(a)分析可知,相同时间段内,净光合速率大的处理胞间CO2浓度小,净光合速率小的处理胞间CO2浓度大。试验期内,各处理整体服从CK>T2≥T3>T1的分布规律。

图5 各处理叶片蒸腾速率及胞间CO2浓度变化特征
2.2 幼龄苹果树叶片净光合速率与影响因子的统计分析
2.2.1净光合速率与影响因子的通径分析
光合作用是植物在光照及一定的温度条件下通过CO2和H2O合成有机物并释放出O2的生理生化反应。在此过程中,光合有效辐射是光合作用的能量来源,CO2和H2O是光合作用的物质基础,温度是光合作用的反应条件,都对光合作用存在着影响[10]。为进一步量化各影响因子对光合作用直接与间接的作用程度,对净光合速率与土壤含水率、气孔导度、胞间CO2浓度、蒸腾速率、光辐射强度等影响因子进行通径分析,见表3。
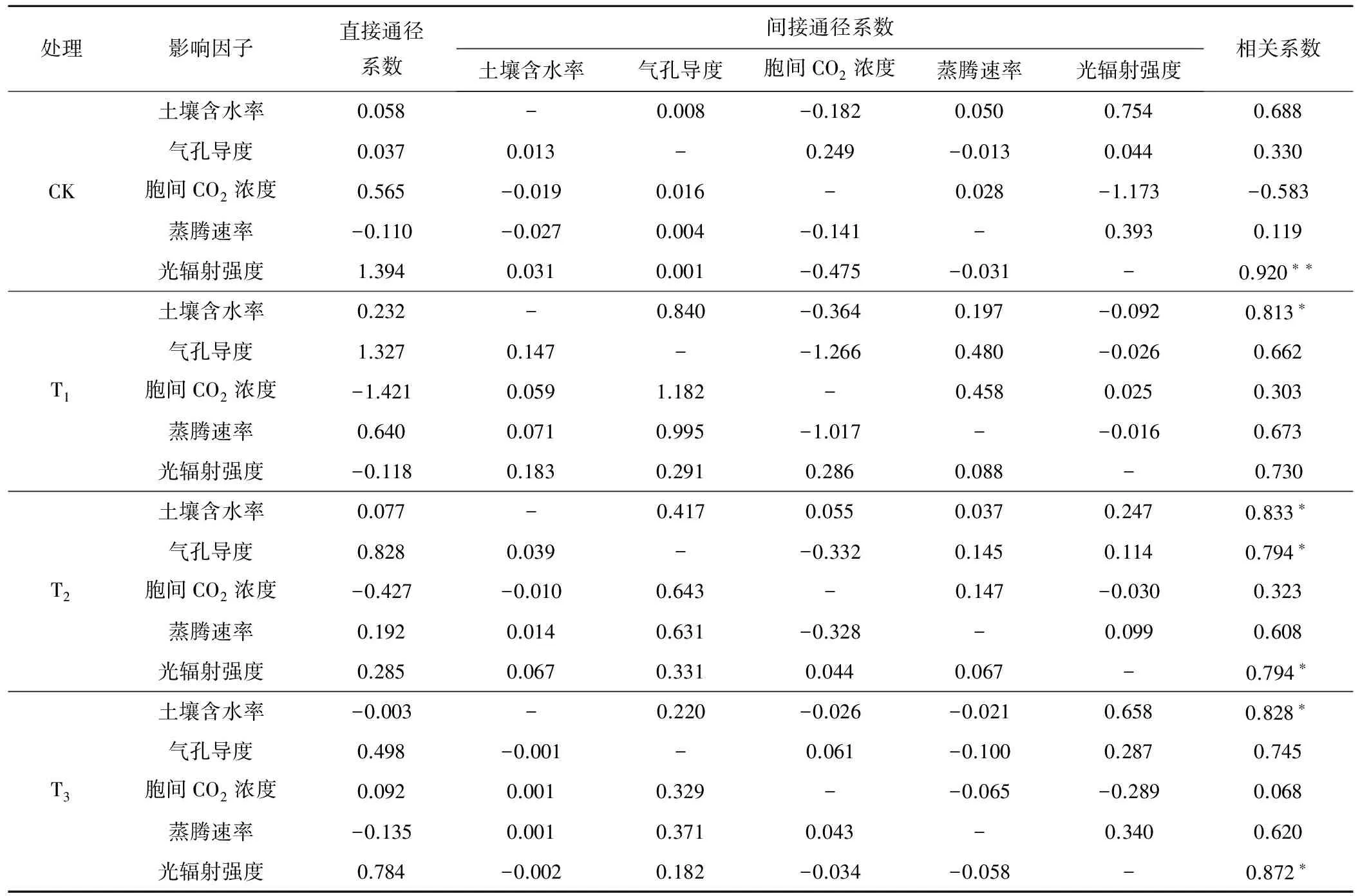
表3 各处理净光合速率与影响因子的相关分析及通径分析
注:**表示在0.01水平上(双侧)显著相关,*表示在0.05水平上(双侧)显著相关。
通径分析由S·Wright于1921年首先提出,其实质是标准化的多元线性回归分析。它将自变量与因变量的简单相关系数加以分解,分解出自变量对因变量的直接作用效应和通过其他变量对因变量的间接作用效应,且根据通径系数大小可以判断各自变量对因变量贡献的相对重要性[15-17],目前在众多研究领域中有着广泛地应用。
对于CK处理,各因子对净光合速率的影响程度为光辐射强度>土壤含水率>胞间CO2浓度>气孔导度>蒸腾速率,直接通径系数按绝对值排列为光辐射强度>胞间CO2浓度>蒸腾速率>土壤含水率>气孔导度。光辐射强度与净光合速率呈极显著相关(0.920)关系,且直接通径系数最大,说明地面灌溉条件下光合有效辐射直接作用于幼树的光合作用,是限制幼树净光合速率的主要因素。土壤含水率与净光合速率的相关系数达0.688,但其直接通径系数仅为0.058,说明土壤水分主要是通过其他因子对净光合速率产生影响。胞间CO2浓度的直接通径系数为0.565,对净光合速率有明显的正效应,但其通过光辐射强度对净光合速率却产生了明显的负效应(-1.173),最终表现为对光合作用的抑制效应(-0.583),说明光照过强时,限制了叶片气孔开度,影响了植株与大气的CO2交换,导致净光合速率下降。
对于T1处理,各因子对净光合速率的影响程度为土壤含水率>光辐射强度>蒸腾速率>气孔导度>胞间CO2浓度,直接通径系数按绝对值排列为胞间CO2浓度>气孔导度>蒸腾速率>土壤含水率>光辐射强度。土壤含水率与净光合速率呈显著相关(0.813)关系,说明在灌水下限为50%时,计划湿润层内土壤含水率较低,对幼树造成了轻微的水分胁迫,土壤水分状况成为限制净光合速率的主要因子。其直接通径系数为0.232,主要通过气孔导度(0.840)和胞间CO2浓度(-0.364)对光合作用产生影响。光辐射强度的直接通径系数为-0.118,间接通径系数均为正数,与净光合速率的相关系数为0.730,说明在轻微水分胁迫的条件下,适当增加光照强度,可以显著促进其他因子对光合作用的正效应,进而提高净光合速率。胞间CO2浓度的直接通径系数为-1.421,被其通过气孔导度(1.182)和蒸腾速率(0.458)产生的正效应所掩盖,最终表现为对光合作用的促进作用。
对于T2处理,各因子对净光合速率的影响程度为土壤含水率>光辐射强度=气孔导度>蒸腾速率>胞间CO2浓度,直接通径系数按绝对值排列为气孔导度>胞间CO2浓度>光辐射强度>蒸腾速率>土壤含水率。土壤含水率(0.833)、光辐射强度(0.794)、气孔导度(0.794)与净光合速率呈显著相关关系,土壤含水率和光辐射强度的直接通径系数分别为0.077和0.285,都明显小于各自的相关系数,但二者各项间接通径系数均为正数,说明在T2处理条件下,适当增加土壤含水率及光照强度,可以显著促进其他因子对光合作用的正效应,进而提高净光合速率。气孔导度的直接通径系数为0.828,与其相关系数差异不大,说明气孔主要通过自身开闭影响净光合速率。各因子通过气孔导度作用于净光合速率的间接通径系数分别为0.417(土壤含水率)、0.643(胞间CO2浓度)、0.631(蒸腾速率)、0.331(光辐射强度),均为各因子间接通径系数的最大值,说明蓄水坑灌条件下,各因子主要通过自身的直接作用及气孔导度的间接作用对净光合速率产生影响,这与张敏的研究成果较为相似[18]。
对于T3处理,各因子对净光合速率的影响程度为光辐射强度>土壤含水率>气孔导度>蒸腾速率>胞间CO2浓度,直接通径系数按绝对值排列为光辐射强度>气孔导度>蒸腾速率>胞间CO2浓度>土壤含水率。光辐射强度(0.872)和土壤含水率(0.828)与净光合速率呈显著相关关系,二者的直接通径系数分别为0.784和-0.003,即光辐射强度对净光合速率有直接、显著的正效应,而土壤含水率主要通过其他因子的间接作用对净光合速率产生正效应,说明在土壤水分供应充足的情况下,光辐射强度是限制净光合速率的主要因素。蒸腾速率直接通径系数为-0.135,对光合作用产生直接的抑制效果,但其通过气孔导度(0.371)和光辐射强度(0.340)对光合作用的正效应,最终表现为促进净光合速率的提高(0.620)。
2.2.2净光合速率与影响因子的回归分析
为进一步研究净光合速率与影响因子间的量化关系,以净光合速率Y为因变量,对各影响因子:土壤含水率X1、气孔导度X2、胞间CO2浓度X3、蒸腾速率X4、光辐射强度X5进行多元逐步回归分析,拟合方程见表4。

表4 净光合速率与影响因子的拟合方程
由表4可见,各组处理逐步回归方程拟合程度高(R≥0.813),显著性强(Sig.≤0.026<0.05),具有明显的统计学意义。回归方程说明,CK处理净光合速率主要与胞间CO2浓度X3和光辐射强度X5有关;T1处理净光合速率仅与土壤含水率X1有关;T2处理净光合速率主要与土壤含水率X1和气孔导度X2有关;T3处理净光合速率主要与和蒸腾速率X4和光辐射强度X5有关。进一步分析发现,当土壤水分成为限制条件时,幼树净光合速率主要受土壤含水率的影响(T1、T2),当幼树受到轻微水分胁迫时,土壤含水率成为了唯一的影响因子(T1),而当计划湿润层土壤水分供应充足时,幼树净光合速率主要受光辐射强度的影响(T3、CK),这与通径分析的结果相互印证。
3 结 语
由于试验条件所限,本文仅探讨了2015年6-9月蓄水坑灌条件下3 a生矮砧红富士叶片光合特性对不同灌水下限的响应,所得结果虽可为幼龄苹果树的水分科学管理提供一定参考,但数据系列较短,仅能代表幼龄苹果树叶片在当地气候条件和试验水平下光合特性的变化规律。在后续的大田试验中,为进一步完善和深入研究幼龄苹果树叶片光合特性对不同灌水下限的响应,还应延长观测年限,取得更长系列的试验数据,以期获得更具普遍性和代表性的响应规律。
通过研究蓄水坑灌条件下不同灌水下限幼龄苹果树叶片光合指标的变化规律,结合通径分析与多元逐步回归分析的结果,并以地面灌溉作为对照,得出以下主要结论。
(1)不同灌水下限幼龄苹果树净光合速率的变化规律基本一致,试验期内呈波动上升趋势,9月12日达到峰值,整体服从T1>T2≥T3>CK的分布规律。由通径分析及多元逐步回归分析可知,净光合速率受多个因子直接和间接的影响,其中,土壤含水率和光合有效辐射是最主要的。当土壤水分成为限制条件时,净光合速率主要受土壤含水率的影响(T1、T2),产生水分胁迫后,土壤含水率成为唯一的影响因子(T1),当土壤水分供应充足时,净光合速率主要受光合有效辐射的影响(T3、CK)。与地面灌溉相比,蓄水坑灌条件下各因子主要通过自身的直接作用及气孔导度的间接作用对净光合速率产生影响。
(2)不同灌水下限幼龄苹果树气孔导度的变化规律基本一致,试验期内呈先增大、后减小、再增大的单峰分布,整体服从CK>T2>T3>T1(7月26日前)和T1>T3>T2>CK(7月26日后)的分布规律。气孔导度的变化主要受光合有效辐射的影响,且灌水下限相同时,地面灌溉对光合有效辐射的响应较蓄水坑灌更积极。
(3)不同灌水下限幼龄苹果树蒸腾速率的变化规律基本一致,试验期内呈典型的双峰分布,整体服从呈T3>T2≥T1>CK的分布规律。当光合有效辐射小于1 000~1 200 μmol/(m2·s)时,各处理蒸腾速率随土壤含水率及光合有效辐射水平的增加而增加,当光合有效辐射大于1 000~1 200 μmol/(m2·s)时,各处理蒸腾速率随光合有效辐射的增加而降低。土壤水分充足时,蒸腾速率对光合有效辐射的响应更敏感,光合有效辐射水平差异不大时,土壤含水率越高蒸腾速率越大。
(4)不同灌水下限幼龄苹果树胞间CO2浓度的变化规律基本一致,试验期内呈先增大,后减小,再增大的单峰分布,整体服从CK>T2≥T3>T1的分布规律。净光合速率越大胞间CO2浓度越小。
[1] 孙西欢,马娟娟,周青云,等.蓄水坑灌法技术要素初探[J].沈阳农业大学学报,2004,35(5/6):405-407.
[2] 王 红,王百田,王 婷,等.不同土壤含水量对山杏盆栽幼苗蒸腾速率和光合速率的影响[J].北方园艺,2010,(2):1-5.
[3] 刘 玲,刘淑明,孙丙寅.不同产地花椒幼苗光合特性研究[J].西北农业学报,2009,18(3):160-165.
[4] 王景燕,龚 伟,包秀兰.水肥耦合对汉源花椒幼苗叶片光合作用的影响[J].生态学报,2016,36(5):1-10.
[5] 郑文君.渭北苹果光合特性的研究[D]. 陕西杨凌:西北农林科技大学,2007.
[6] 张 雯,安贵阳,李翠红.肥水分区调控对苹果光合作用、生长结果和果实品质的影响[J].西北农业学报,2010,19(6):110-114.
[7] 马慧丽,吕德国.干旱-复水处理对寒富苹果光合特性的影响[J].华北农学报,2014,29(4):181-185.
[8] 姜卫兵,高光林,俞开锦,等.水分胁迫对果树光合作用及同化代谢的影响研究进展[J].果树学报,2012,19(6):416-420.
[9] 卢 艳,王 飞,韩明玉,等. NaCl胁迫对4种砧穗组合苹果的生长及光合特性的影响[J].西北农业学报,2011,20(8):106-110.
[10] 张 敏,孙西欢,郭向红,等.蓄水坑灌下苹果树光合日变化与影响因子的分析[J].中国科技论文,2015,10(9):1 095-1 100.
[11] 张西平,赵胜利,张旭东,等.不同灌水处理对温室黄瓜形态及光合作用指标的影响[J].中国农学通报,2007,23(6):622-625.
[12] 李远华,罗金耀.灌溉理论与技术[M].武汉:武汉大学出版社,2003-10:11-17.
[13] 雷 涛,郭向红,孙西欢,等.不同灌溉方式下果园棵间蒸发特性研究[J].灌溉排水学报,2014,33(4/5):179-184.
[14] 张卫强.黄土半干旱区主要树种光合生理与耗水特性研究[D].北京:北京林业大学,2006.
[15] 魏清顺,孙西欢,刘在伦.导流器几何参数对潜水泵性能影响的通径分析[J].排灌机械工程学报,2014,2(3):202-207.
[16] 张 华,徐春华,张 兰,等.基于通径分析的兰州北山三种典型植物光合作用影响因子[J].生态学杂志,2015(5):1 289-1 294.
[17] 蔡甲冰,刘 钰,许 迪,等.基于通径分析原理的冬小麦缺水诊断指标敏感性分析[J].水利学报,2008,39(1):83-90.
[18] 张 敏.蓄水坑灌条件下苹果树光合特性与影响因子的分析研究[D]. 太原:太原理工大学,2015.
