胡桃科6个树种花粉形态的扫描电镜观察
2016-02-16李晓春刘晶晶洑香香
毛 霞, 李晓春, 刘晶晶, 洑香香,①
(南京林业大学: a. 南方现代林业协同创新中心, b. 林学院, 江苏 南京210037)
胡桃科6个树种花粉形态的扫描电镜观察
毛 霞a,b, 李晓春a,b, 刘晶晶a,b, 洑香香a,b,①
(南京林业大学: a. 南方现代林业协同创新中心, b. 林学院, 江苏 南京210037)
胡桃科; 花粉; 形态特征; 扫描电镜; 演化地位
中国有胡桃科(Juglandaceae)植物约7属27种1变种,南方和北方均有分布[1];该科植物通常为落叶或半常绿乔木或小乔木,大部分种类具有药用、材用和食用等价值,其中,青钱柳〔Cyclocaryapaliurus(Batal.) Iljinsk.〕为重要的药用植物,其叶的多糖复合物可增强糖尿病小鼠对葡萄糖的耐受力[2-3];胡桃(JuglansregiaLinn.)和美国山核桃〔Caryaillinoinensis(Wangenh.) K. Koch〕为重要的干果和油料作物,前者也是重要的硬木产品,果皮可用于制造活性炭[4];枫杨(PterocaryastenopteraC. DC.)是优良的家具用材,其树皮富含纤维[5]。随胡桃科应用价值的不断挖掘,人们对胡桃科植物潜在资源的研究也不断深入。
在植物世代相传过程中,花粉因能保持其固有的形态特征而成为物种进化研究的重要证据,也成为鉴别植物种的重要依据之一,通过扫描电镜观察花粉形态的差异可在一定程度上阐释园艺植物种或品种的分类和系统进化关系[6-7]。目前,已有研究者采用孢粉及其化石证据对胡桃科山核桃属(CaryaNutt.)、胡桃属(JuglansLinn.)和化香树属(PlatycaryaSieb. et Zucc.)的系统演化地位进行了探讨[8-9],但对胡桃科各属代表性种类的花粉形态特征尚无系统的研究报道。鉴于此,作者采用扫描电镜观察胡桃科4属6个主要栽培树种花粉的亚显微特征,并据此初步探讨各属种的系统演化地位。
1 材料和方法
1.1 材料
供试树种包括枫杨属(PterocaryaKunth)的枫杨,胡桃属的胡桃、华东野核桃〔Juglanscathayensisvar.formosana(Hayata) A. M. Lu et R. H. Chang〕和美国黑核桃(JuglansnigraLinn.),青钱柳属(CyclocaryaIljinsk.)的青钱柳及山核桃属的美国山核桃,均种植于南京林业大学校园内。于2010年3月至5月从各树种成年植株上采集盛花期的雄花序,置于室内进行水培,收集花粉并置于0 ℃~4 ℃条件下保存、备用。
1.2 方法
1.2.1 扫描电镜观察 供试6个树种花粉用质量体积分数4%戊二醛固定12 h以上,随后用0.1 mol·L-1磷酸缓冲液漂洗3次,每次5 min;然后用质量体积分数1%锇酸固定2~3 h,随后用0.1 mol·L-1磷酸缓冲液漂洗3次;经无水乙醇脱水,随后依次用体积分数50%和100%丙酮溶液各置换1次,每次5 min;用EMITECH K850型临界点干燥仪(英国EMITECH公司)干燥4 h,随后用E-1010型离子溅射镀膜仪(日本HITACHI公司)在花粉表面喷金属银膜(厚度15 nm),然后用QUANTA-200型扫描电子显微镜(美国FEI公司)观察花粉形态的亚显微特征并拍照。
1.2.2 数据测定和分析 每个树种随机选取20粒花粉,分别测量极轴长(P)和赤道轴长(E),并计算极轴长与赤道轴长的比值(P/E,表示花粉形状)及极轴长与赤道轴长的乘积(P×E,表示花粉大小)[10-11]。花粉形态描述术语参见文献[12]。采用EXCEL 2010和SAS 18.1软件对数据进行处理和分析。
2 结果和分析
扫描电镜下胡桃科6个树种花粉的形态特征见图1,各树种花粉的相关形态参数见表1。
由图1可见:6个树种的花粉均为单粒花粉,表面纹饰均为分布均匀的颗粒状雕纹,且较为密集。萌发孔为复式萌发孔,主要分布在赤道面上;萌发孔稍凸出于花粉表面,外壁2层在萌发孔处分离形成孔室,且变化复杂。6个树种的花粉直径为25~50 μm,根据花粉分级标准,皆属于中等型花粉[13]。
由表1可见:供试6个树种花粉的极轴长(P)为25.00~37.61 μm,赤道轴长(E)为21.90~36.55 μm,其中,美国山核桃花粉的极轴长和赤道轴长均最大,枫杨花粉均最小。根据花粉P/E值可将6个树种花粉形状划分为3类:扁球形(胡桃、美国黑核桃、华东野核桃和美国山核桃)、扁五角形(青钱柳)和扁多角形(枫杨),3类花粉对应的极面观分别为圆形、五角形和多边形;枫杨花粉P/E值变异系数最大,达12%,说明枫杨种内花粉形状差异较大;而其余5个树种花粉P/E值变异系数为2%~4%,说明它们的花粉形状在种内差异较小。6个树种花粉的P×E值由小至大依次为枫杨、华东野核桃、青钱柳、美国黑核桃、胡桃、美国山核桃,其中,美国黑核桃与华东野核桃、华东野核桃与青钱柳间花粉的P×E值无显著差异;枫杨花粉P×E值的变异系数最大,青钱柳次之,其余4个树种花粉P×E值的变异系数为3%~6%,表明枫杨种内花粉大小差异较大,青钱柳次之,而其他树种的花粉大小差异较小。
由表1还可见:美国黑核桃、胡桃和华东野核桃的花粉萌发孔数较多(8~26),均为散孔类型;青钱柳、枫杨和美国山核桃的花粉萌发孔数均较少(3~6),均为赤道孔类型;其中,美国山核桃花粉萌发孔数最少(具3个萌发孔),美国黑核桃花粉萌发孔数最多(平均值为22)。此外,根据花粉对称型可将6个树种花粉分为2类:左右对称型(枫杨、青钱柳和美国山核桃)和完全对称型(胡桃、美国黑核桃和华东野核桃)。
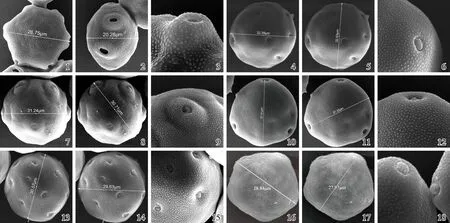
1-3. 枫杨PterocaryastenopteraC. DC.: 1. 极轴长Polar axis length; 2. 赤道轴长Equatorial axis length; 3. 表面纹饰Exine ornamentation. 4-6. 胡桃JuglansregiaLinn.: 4. 极轴长Polar axis length; 5. 赤道轴长Equatorial axis length; 6. 表面纹饰Exine ornamentation. 7-9. 华东野核桃Juglanscathayensisvar.formosana(Hayata) A. M. Lu et R. H. Chang: 7. 极轴长Polar axis length; 8. 赤道轴长Equatorial axis length; 9. 表面纹饰Exine ornamentation. 10-12. 美国山核桃Caryaillinoinensis(Wangenh.) K. Koch: 10. 极轴长Polar axis length; 11. 赤道轴长Equatorial axis length; 12. 表面纹饰Exine ornamentation. 13-15. 美国黑核桃JuglansnigraLinn.: 13. 极轴长Polar axis length; 14. 赤道轴长Equatorial axis length; 15. 表面纹饰Exine ornamentation. 16-18. 青钱柳Cyclocaryapaliurus(Batal.) Iljinsk: 16. 极轴长Polar axis length; 17. 赤道轴长Equatorial axis length; 18. 表面纹饰Exine ornamentation.
图1 扫描电镜下胡桃科6个树种的花粉形态
Fig. 1 Morphology of pollen of six tree species in Juglandaceae under scanning electron microscope
表1 胡桃科6个树种花粉的相关形态参数
Table 1 Related morphological parameters of pollen of six tree species in Juglandaceae
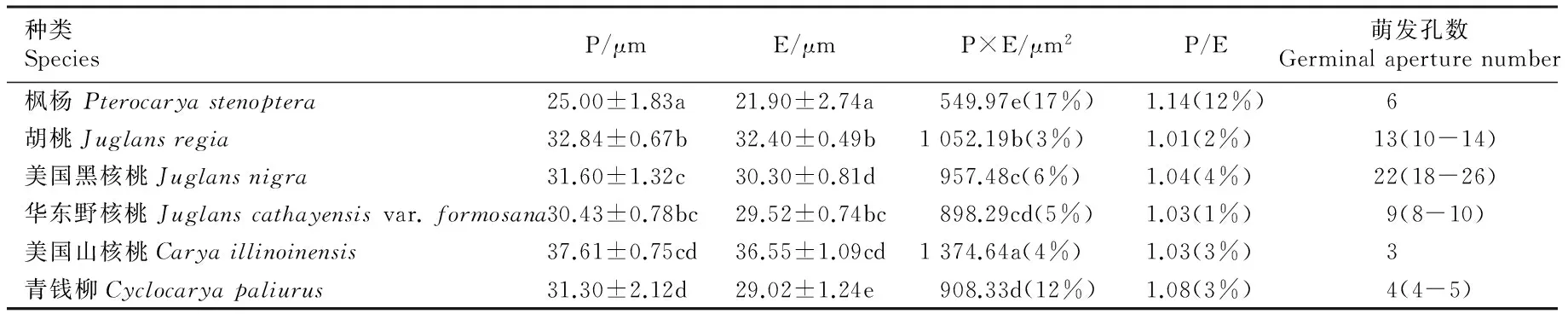
种类SpeciesP/μmE/μmP×E/μm2P/E萌发孔数Germinalaperturenumber枫杨Pterocaryastenoptera25.00±1.83a21.90±2.74a549.97e(17%)1.14(12%) 6胡桃Juglansregia32.84±0.67b32.40±0.49b1052.19b(3%)1.01(2%) 13(10-14)美国黑核桃Juglansnigra31.60±1.32c30.30±0.81d957.48c(6%)1.04(4%) 22(18-26)华东野核桃Juglanscathayensisvar.formosana30.43±0.78bc29.52±0.74bc898.29cd(5%)1.03(1%) 9(8-10)美国山核桃Caryaillinoinensis37.61±0.75cd36.55±1.09cd1374.64a(4%)1.03(3%) 3青钱柳Cyclocaryapaliurus31.30±2.12d29.02±1.24e908.33d(12%)1.08(3%) 4(4-5)
1)P: 极轴长 Polar axis length; E: 赤道轴长 Equatorial axis length; P×E: 极轴长与赤道轴长的乘积 The product of polar axis length and equatorial axis length; P/E: 极轴长与赤道轴长的比值 Ratio of polar axis length to equatorial axis length. 同列中不同的小写字母表示差异显著(P<0.05) Different small letters in the same column indicate the significant difference (P<0.05). 括号中的百分数为变异系数 Percentages in brackets are the coefficient of variance.
3 讨论和结论
在植物世代遗传过程中,由于花粉形态的遗传保守性,其形态特征受外界环境的影响较少且相对稳定,为研究物种的亲缘关系和系统演化提供了依据。Wodehouse[14]认为,进化程度高的花粉调节功能强,而花粉的调节功能则随其体积和表面积比值的减小而增强。一般认为,较原始的被子植物花粉体积较大,花粉体积从大向小演化。据此推测:在供试的6个树种中,枫杨(枫杨属)的演化程度最高,青钱柳(青钱柳属)及胡桃属的3个树种(华东野核桃、美国黑核桃、胡桃)均处于中等演化水平,美国山核桃(山核桃属)的演化程度最低。刘成运[15]认为,周孔类花粉最进化,3孔花粉次之,3孔沟花粉则较原始。本研究中,美国山核桃花粉为3孔型,相对于胡桃、青钱柳和枫杨较为原始。原始植物花粉均具有光滑且无定型的外壁,花粉表面纹饰的演化趋势为表面光滑→表面具小穴、小沟状雕纹→粗糙并具疣状纹饰→表面颗粒状→表面网状[16]。本研究中,供试的6种胡桃科植物的花粉均为单粒花粉,表面纹饰均为密集分布的颗粒状并分布在整个花粉表面,由此可以推测胡桃科植物进化较快。
韩丽娟等[17]对胡桃科植物次生木质部导管分子的解剖研究结果表明:在胡桃科中,山核桃属最进化,核桃属和枫杨属均处于中等进化水平,且枫杨属比核桃属稍进化,与本文结果存在差异。Manning[18]根据胡桃科的花序形态推断胡桃属、山核桃属和枫杨属较进化,与本文观点相一致。由此可见,仅以单一性状判断植物的演化顺序具有一定局限性,需依据多学科研究证据分析胡桃科植物的演化关系。
[1] 郑万钧. 中国树木志: 第二卷[M]. 北京: 中国林业出版社, 1985: 2359-2395.
[2] 方升佐, 洑香香. 青钱柳资源培育与开发利用的研究进展[J]. 南京林业大学学报(自然科学版), 2007, 31(1): 95-100.
[3] 李 磊, 谢明勇, 易 醒, 等. 青钱柳多糖组分及其降血糖活性研究[J]. 江西农业大学学报, 2001, 23(4): 484-486.
[4] 李一帆, 江 莎, 陈旭辉. 胡桃科植物研究进展[J]. 内蒙古农业大学学报, 2006, 27(3): 148-152.
[5] 张占敏, 侯利峰. 枫杨育苗技术及其在园林绿化中的作用[J]. 安徽农学通报, 2013, 19(11): 108, 145.
[6] 冯立国, 邵大伟, 生利霞, 等. 野生玫瑰的花粉形态及其起源与演化的探讨[J]. 林业科学, 2007, 43(12): 76-80.
[7] 邓小燕, 周颂东, 何兴金. 中国黄精属13种植物花粉形态及系统学研究[J]. 武汉植物学研究, 2007, 25(1): 11-18.
[8] WHITEHEAD D R. Pollen morphology in the Juglandaceae, Ⅱ: survey of the family[J]. Journal of the Arnold Arboretum, 1965, 46(4): 369-410.
[9] LEOPOLD E B, MACGINITIE H D. Development and affinities of Tertiary floras in the Rocky Mountains[M]∥GRAHAM A. Floristics and Paleofloristics of Asia and Eastern North America. Amsterdam: Elsevier, 1972: 147-200.
[10] 牛立新, 张延龙. 中国野生葡萄花粉形态学研究[J]. 园艺学报, 2000, 27(5): 361-363.
[11] 埃尔特曼 G. 孢粉学手册[M]. 中国科学院植物研究所古植物研究室孢粉组, 译. 北京: 科学出版社, 1978: 13-26.
[12] 王伏雄, 钱南芬, 张玉龙, 等. 中国植物花粉形态[M]. 2版. 北京: 科学出版社, 1995: 218-219.
[13] 宁万军, 廖 康, 王国安, 等. 新疆主栽核桃品种花粉量及花粉形态研究[J]. 新疆农业科学, 2014, 51(2): 220-226.
[14] WODEHOUSE R P. Pollen grains: their structure, indentification and significance in science and medicine[M]. New York: McGraw-Hill Book Company, Inc., 1935: 323-340.
[15] 刘成运. 马尾树科及其近缘科花粉形态研究[J]. 云南植物研究, 1987, 9(2): 181-186.
[16] 贝 克. 被子植物的起源和早期演化[M]. 张芝玉, 陶君蓉, 张玉龙, 等, 译. 北京: 科学出版社, 1981: 190-194.
[17] 韩丽娟, 周春丽, 吴树明. 国产胡桃科次生木质部导管分子的比较解剖及其系统位置的讨[J]. 西北植物学报, 2002, 22(6): 1426-1431.
[18] MANNING W E. The morphology of the flowers of the Juglandaceae. Ⅰ. The inflorescence[J]. American Journal of Botany, 1938, 25(6): 407-419.
(责任编辑: 郭严冬)
Scanning electron microscope observation on pollen morphology of six tree species in Juglandaceae
MAO Xiaa,b, LI Xiaochuna,b, LIU Jingjinga,b, FU Xiangxianga,b,①
(Nanjing Forestry University: a. Co-Innovation Center for Sustainable Forestry in Southern China, b. College of Forestry, Nanjing 210037, China),J.PlantResour. &Environ., 2016, 25(4): 104-106
Pollen morphology of six tree species belonging to four genera in Juglandaceae was observed by scanning electron microscope. The results show that pollens of six tree species are all single grain with medium size, the exine ornamentation is densely distributed by granular glyph with compound germinal apertures scattering or along equatorial; there are great differences in number of germinal apertures (average 3-22), number of germinal apertures ofJuglansnigraLinn. is the most, while that ofCaryaillinoinensis(Wangenh.) K. Koch is the least. Polar axis length of pollen is 25.00-37.61 μm, equatorial axis length is 21.90-36.55 μm. The order of pollen size from small to big isPterocaryastenopteraC. DC.,Juglanscathayensisvar.formosana(Hayata) A. M. Lu et R. H. Chang,Cyclocaryapaliurus(Batal.) Iljinsk.,J.nigra,J.regiaLinn. andC.illinoinensis. Pollen shape ofC.paliurusandP.stenopterais flat pentagonal and flat polygonal, respectively, while that of other four tree species is flat spheroid. According to pollen size, exine ornamentation and germinal aperture morphology, evolutionary relationships of six tree species are discussed.
Juglandaceae; pollen; morphological characteristics; scanning electron microscope; evolutionary status
2015-11-25
国家自然科学基金资助项目(31470637; 31270673); 江苏高校优势学科建设工程项目(PAPD)
毛 霞(1991—),女,江苏南京人,博士研究生,主要从事花芽分化机制的研究。
①通信作者E-mail: xxfu@njfu.edu.cn
Q944.571; Q949.735
A
1674-7895(2016)04-0104-03
10.3969/j.issn.1674-7895.2016.04.13
