湖南八面山银杉群落特征及其残遗性和保守性分析
2016-02-16苏乐怡赵万义张记军杨玉鹏郭远飞廖文波
苏乐怡, 赵万义, 张记军, 杨玉鹏, 郭远飞, 凡 强, 廖文波,①
(1. 中山大学生命科学学院, 广东 广州 510275; 2. 首都师范大学资源环境与旅游学院, 北京 100048;3. 湖南八面山国家级自然保护区管理局, 湖南 桂东 423501)
湖南八面山银杉群落特征及其残遗性和保守性分析
苏乐怡1, 赵万义1, 张记军2, 杨玉鹏1, 郭远飞3, 凡 强1, 廖文波1,①
(1. 中山大学生命科学学院, 广东 广州 510275; 2. 首都师范大学资源环境与旅游学院, 北京 100048;3. 湖南八面山国家级自然保护区管理局, 湖南 桂东 423501)
为了解中国特有孑遗植物银杉(CathayaargyrophyllaChun et Kuang)的生存现状,对位于湖南八面山脚盆辽的银杉+南方铁杉〔Tsugachinensisvar.tchekiangensis(Flous) Cheng et L. K. Fu〕+甜槠〔Castanopsiseyrei(Champ.) Tutch.〕-猴头杜鹃(RhododendronsimiarumHance)群落进行调查,分析其群落特征及种群年龄结构,比较分布在不同区域的6个银杉群落中种子植物属的分布区类型及其相似性系数;在此基础上,讨论了银杉群落的残遗性特征。结果显示:脚盆辽银杉群落共有维管植物66种,包括蕨类植物4种和种子植物62种(裸子植物4种、被子植物58种); 该群落乔木层的主要优势种为甜槠、 猴头杜鹃、 银杉和南方铁杉, 重要值分别为13.12%、 9.28%、 8.86%和7.49%; 该群落的Simpson和Shannon-Wiener多样性指数分别为0.94和3.20, Pielou均匀度指数Jsw和Jsi分别为0.81和0.92,说明该群落物种多样性偏低但物种分布均匀,与中亚热带山地的暖性针阔叶混交林特征一致。从生活型谱看,该群落中各频度百分比由高至低依次为A级、B级、C级、D级、E级,与Raunkiaer标准频度图谱基本一致,表明该群落整体上处于较为稳定的亚顶极状态。从银杉种群年龄结构看,种群中幼龄个体占一定比例,但80~180 a的中龄个体比例偏低;从该群落优势种的径级结构看,甜槠、猴头杜鹃和鹿角杜鹃(RhododendronlatoucheaeFranch.)种群为增长型种群,而银杉、南方铁杉和福建柏〔Fokieniahodginsii(Dunn) Henry et Thomas〕种群为衰退型种群。从脚盆辽银杉群落种子植物属的分布区类型看, 温带分布型属占51.06%, 略高于热带分布型属(占48.94%),说明该区域属于热带亚热带过渡区;除纬度外,海拔也能够在一定程度上影响其分布区类型组成。在处于粤北、 粤桂山地和贵州高原3个植物区系亚地区的6个银杉群落中, 种子植物属分布区类型的相似性系数为0.35~0.67,群落建群种所在属如铁杉属〔Tsuga(Endlicher) Carrière〕、润楠属(MachilusNees)、水青冈属(FagusLinn.)和福建柏属(FokieniaHenry et Thomas)等在各群落间有相似性和共通性,且多为古老成分和孑遗成分,表明现存的银杉群落具有相近的变迁历史,分布地域具有明显的亚热带山地避难所特征,体现出其演替过程的残遗性和保守性。研究结果表明:在银杉群落中适度疏通林窗,降低林下郁闭度,有利于银杉幼树的生长,使银杉种群得到可持续更新和演替。
湖南八面山脚盆辽; 银杉; 群落特征; 分布区类型; 种群年龄结构; 残遗性
银杉(CathayaargyrophyllaChun et Kuang)隶属松科(Pinaceae)银杉属(CathayaChun et Kuang)[1],为中国特有孑遗植物,也是国家一级保护植物,堪称“植物界大熊猫”。从化石证据看,银杉曾广泛分布于中新世至上新世的欧洲中部、北美洲及亚洲,第四纪冰川来临前其分布区大幅度缩减,现仅残存于中国南部山区[2-3],因此,银杉也是“活化石”。
据统计,目前在湖南八面山、广西大瑶山、贵州大娄山和湖南越城岭4个山区内的36个分布点共有银杉3 018株,其中八面山地区有879株[4]。湖南八面山是中国银杉群落分布的东界,自从1986年在八面山脚盆辽发现银杉群落,之后10年陆续对八面山的银杉群落状况开展了调查和研究,并探讨了其亚热带常绿针叶林残遗性质以及银杉群落的演替趋势,在此基础上,提出设立银杉定位研究基地的建议[5-7]。近年来研究者对银杉群落的研究热度降低,八面山地区的银杉生存现状需要进一步评价。此外,中国亚热带森林植物区系发生历史古老,而星散分布于亚热带地区的银杉群落结构在演替过程中是否具有保守性,这一问题也未见相关研究报道。
此外,银杉为著名的第三纪孑遗种,在地史时期曾广布于欧洲、亚洲及北美洲,以银杉为特征种的群落在中国亚热带地区形成若干残遗分布地,有着重要的生物地理学意义[2,8]。谢宗强等[9]认为,银杉群系虽地处亚热带,但物种的分布区类型组成却具有较强的温带分布型特点。然而,关于银杉现分布地的群落物种分布区类型组成缺乏进一步的对比分析。
作者采用经典样方调查方法,研究了分布于八面山国家级自然保护区脚盆辽的银杉群落特征及其种群结构。在此基础上,对八面山与大瑶山[10-11]、越城岭[12]、大娄山[13]和金佛山[10]分布的银杉群落的地理分布特征进行比较;并结合谱系地理学研究文献[3],探讨银杉群落演化的残遗性及保守性特征。
1 研究区域自然概况和研究方法
1.1 研究区域自然概况
湖南省八面山属于罗霄山脉南段支脉,地处资兴市、桂东县和炎陵县的交界处,地理坐标为北纬25°54′02″~26°06′59″、东经113°37′39″~113°50′08″,境内山体呈“H”形轮廓, 山涧峡谷纵深, 最高海拔2 051 m,最低海拔800 m。地处中亚热带山地湿润气候区, 夏季凉爽、 冬季稍寒冷; 年平均气温13.5 ℃, 7月平均气温22.6 ℃,1月平均气温4.1 ℃,极端最高气温和极端最低气温分别为34.0 ℃和-6.3 ℃;年降水量1 754 mm,4月和5月降水量较多,占全年降水量的33%,10月至次年1月降水量较少,仅占全年降水量的10%。成土母岩以花岗岩为主,兼有石英砂岩和变质岩;土壤为山地黄壤及黄棕壤,土层疏松且含水量低,腐殖层较薄[7]。
银杉群落主要分布于八面山国家级自然保护区东北侧支脉溪谷两侧山坡的中部和下部,形成典型的针阔叶混交林,邻近局部地区也有零星分布,垂直分布于海拔1 000~1 400 m。脚盆辽为该自然保护区内银杉分布较集中的区域,银杉在河谷内多与猴头杜鹃(RhododendronsimiarumHance)和金叶含笑(MicheliafoveolataMerr. ex Dandy)混生,而在山坡、悬岩等区域则与甜槠〔Castanopsiseyrei(Champ.) Tutch.〕和鹿角杜鹃(RhododendronlatoucheaeFranch.)混生。
1.2 研究方法
1.2.1 样地设置和调查方法 选取银杉、南方铁杉〔Tsugachinensisvar.tchekiangensis(Flous) Cheng et L. K. Fu〕、猴头杜鹃和甜槠混生的群落,划定面积1 600 m2的样地;样地的地理坐标为北纬26°04′10″、东经113°42′58″,海拔1 107 m,坡度约30°~50°,坡向西偏南;土层厚度约30 cm,土壤中多有植物根系,土质疏松,土壤相对含水量低。
[14-15]的方法,将样地划分为16个面积10 m×10 m的大样方,采取每木记账法调查大样方内胸径>1.5 cm、高度≥1.5 m的乔木和灌木的种名、胸围、高度、冠幅和株数。在每个大样方内划出1个面积2 m×2 m的小样方,记录小样方内乔木和灌木的幼苗及草本植物的种名、高度、覆盖度和株数。
1.2.2 群落特征分析方法 参考植物群落学研究方法[14-15],统计样地内各物种的相对多度、相对频度和相对优势度,并根据公式“重要值=(相对多度+相对频度+相对优势度)/3”计算重要值。群落的物种多样性采用Simpson多样性指数(SP)、Shannon-Wiener多样性指数(SW)、Pielou均匀度指数(Jsw和Jsi)进行测度[14-16], 计算公式分别为:SP= 1-∑Pi2;SW= -∑PilnPi;Jsw= (-∑PilnPi)/lnS;Jsi= (1-∑Pi2)/(1-1/S)。式中,S为样方中的物种总数;Pi为第i种的物种数占样方中物种总数的百分数。
参照Raunkiaer[17]的方法进行生活型频度级分析。其中,频度1%~20%为A级;频度21%~40%为B级;频度41%~60%为C级;频度61%~80%为D级;频度81%~100%为E级。
由于脚盆辽银杉群落中部分优势种为小型乔木,Ⅴ级立木划分标准[14]不适合其种群年龄结构分析,因此,依据胸径频度分析群落中各优势种的年龄结构。根据回归方程“A=1.54+5.20D-0.016 4D2(式中,A为年龄;D为胸径)[18]”(R=0.92**)绘制脚盆辽银杉群落中银杉的年龄结构图。
1.2.3 群落种子植物地理分布的分析方法 通过查阅文献,收集其他区域分布的银杉群落的物种组成信息;根据吴征镒等[19-20]对中国种子植物属分布区类型的划分原则,对各银杉群落种子植物属的分布区类型进行划分;采用相似性系数(Sc)衡量各银杉群落间种子植物属分布区类型的相似性,计算公式为Sc=2c/(A+B)。式中,A为甲群落属数;B为乙群落属数;c为甲、乙群落共有的属数[21]。
2 结果和分析
2.1 群落特征分析
2.1.1 种类组成及结构分析 调查结果显示:湖南八面山脚盆辽的银杉+南方铁杉+甜槠-猴头杜鹃群落(简称脚盆辽银杉群落)内共有维管植物27科50属66种,其中蕨类植物2科3属4种,裸子植物2科4属4种,被子植物23科43属58种。从生活型看,木本植物占明显优势,有61种,占总种数的92.42%,其中,乔木35种,灌木(含乔木幼树)26种;草本植物5种,占总种数的7.58%;未发现藤本植物。
脚盆辽银杉群落终年常绿,林冠层起伏度大。乔木层总郁闭度0.8,可分为3个亚层:高大的银杉和南方铁杉突出于林冠层, 为第1亚层, 高度为15.0~ 18.0 m,老龄甜槠高达15.0 m; 第2亚层高度为10.0~15.0 m,主要为福建柏〔Fokieniahodginsii(Dunn) Henry et Thomas〕、银杉、青冈〔Cyclobalanopsisglauca(Thunb.) Oerst.〕、甜槠和猴头杜鹃,伴生种类为日本杜英(ElaeocarpusjaponicusSieb. et Zucc.)和细叶青冈〔Cyclobalanopsisgracilis(Rehd. et Wils.) Cheng et T. Hong〕等;第3亚层高度为5.0~10.0 m,优势种为鹿角杜鹃、猴头杜鹃、辣汁树(CinnamomumtsangiiMerr.)、美丽新木姜子〔Neolitseapulchella(Meissn.) Merr.〕和南岭山矾(SymplocosconfusaBrand)。灌木层高1.5~4.0 m,种类较丰富,主要为毛玉山竹〔Yushaniabasihirsuta(McClure) Z. P. Wang et G. H. Ye〕、赤楠(SyzygiumbuxifoliumHook. et Arn.)、杜鹃(RhododendronsimsiiPlanch.)、鹿角杜鹃和小果珍珠花〔Lyoniaovalifoliavar.elliptica(Sieb. et Zucc.) Hand.-Mazz.〕;其他种类还有少花柏拉木〔Blastuspauciflorus(Benth.) Guillaum.〕和凤凰润楠(MachilusphoenicisDunn)等。草本层种类较少,多为耐旱的狗脊〔Woodwardiajaponica(Linn. f.) Sm.〕、里白〔Diplopterygiumglaucum(Thunb. ex Houtt.) Nakai〕、芒萁〔Dicranopterispedata(Houtt.) Nakaike〕以及鳞毛蕨科(Dryopteridaceae)的种类。
群落中喜阳性的银杉、南方铁杉和甜槠占据群落上层生态位,但大树数量不多,总体覆盖度不高;乔木第2和第3亚层树种较丰富,使群落整体郁闭度增大。群落林下层草本及幼苗较少,藤本植物缺失。在林下仅发现银杉幼苗14株,可能因群落中灌木层覆盖度高使得草本层光线不充足,同时群落地处坡地、土层保水性差以及人类活动等因素制约了林下层草本植物及银杉幼苗的生长。
2.1.2 群落优势种的重要性分析 湖南八面山脚盆辽银杉群落优势种重要性指标的分析结果见表1。脚盆辽银杉群落中共有22个树种的重要值达到1%以上,而重要值大于4%的树种有7种,为甜槠、猴头杜鹃、银杉、南方铁杉、鹿角杜鹃、赤楠和福建柏,它们的重要值之和达到53.89%,为本群落优势种。其中,甜槠、银杉和南方铁杉的相对优势度分别达到29.55%、18.47%和17.81%,明显高于其他树种,在群落结构上占据上层生态位,据此认为三者是本群落的建群种。除以上优势种外,群落中其他树种的重要值均较低,星散分布于群落中下层。
在科级水平,脚盆辽银杉群落的优势科为壳斗科(Fagaceae)、松科和杜鹃花科(Ericaceae),柏科(Cupressaceae)、桃金娘科(Myrtaceae)、山茶科(Theaceae)和山矾科(Symplocaceae)也占据一定地位。综合分析结果表明:该群落主要建群种均为中亚热带优势种,在植被区划上属东部中亚热带常绿阔叶林带南岭山地栲类林、蕈树林区(Ⅳ Aiib-3)[8],南岭山地一带分布的银杉群落具明显的中亚热带山地性质和孑遗植被特征,在植被和区系地理上有着重要的科学意义。
2.1.3 群落物种多样性分析 群落物种多样性分析结果表明:湖南八面山脚盆辽银杉群落的物种丰富度指数为66;木本层Simpson多样性指数(SP)为0.94,Shannon-Wiener多样性指数(SW)为3.20;Pielou均匀度指数较高,Jsw值为0.81,Jsi值为0.92,Jsw和Jsi值均在0.8以上。表明该银杉群落的物种多样性程度偏低,但群落中物种分布均匀,且虽然物种数量不多但群落目前处于较为稳定的状态。
2.1.4 群落物种频度分析 参照Raunkiaer[17]的方法对湖南八面山脚盆辽银杉群落进行物种生活型的频度级分析,并与亚热带针阔叶混交林江西三清山华东黄杉(PseudotsugagausseniiFlous)群落[22]、广东南岭国家级自然保护区疏齿木荷(SchimaremotiserrataH. T. Chang)+福建柏群落[23]及Raunkiaer的标准频度图谱[17]进行比较,结果见图1。
由图1可以看出:脚盆辽银杉群落中,A级、B级、C级、D级和E级的频度百分比分别为56.25%、 17.19%、 12.50%、 10.40%和3.12%,A级占明显优势。与Raunkiaer标准频度图谱相比,该银杉群落A级、B级、C级和D级的频度百分比略大,而E级的频度百分比则降低13%。E级植物是群落中的优势种和建群种,在脚盆辽银杉群落中,频度级达到E级的只有甜槠和鹿角杜鹃,二者属于中小型乔木,常作为亚热带常绿阔叶林伴生种出现;而其他优势种和建群种中,银杉和猴头杜鹃的频度级为D级,南方铁杉的频度级为C级,说明该银杉群落中银杉和南方铁杉这2种针叶树种在群落中的分布不均匀,优势度也不十分明显,群落呈现出向以甜槠为建群种的常绿阔叶林演替的趋势。
表1 湖南八面山脚盆辽银杉群落优势种重要性指标的分析结果1)
Table 1 Analysis result on important index of dominant species inCathayaargyrophyllaChun et Kuang community at Jiaopenliao of Bamianshan in Hu’nan Province1)
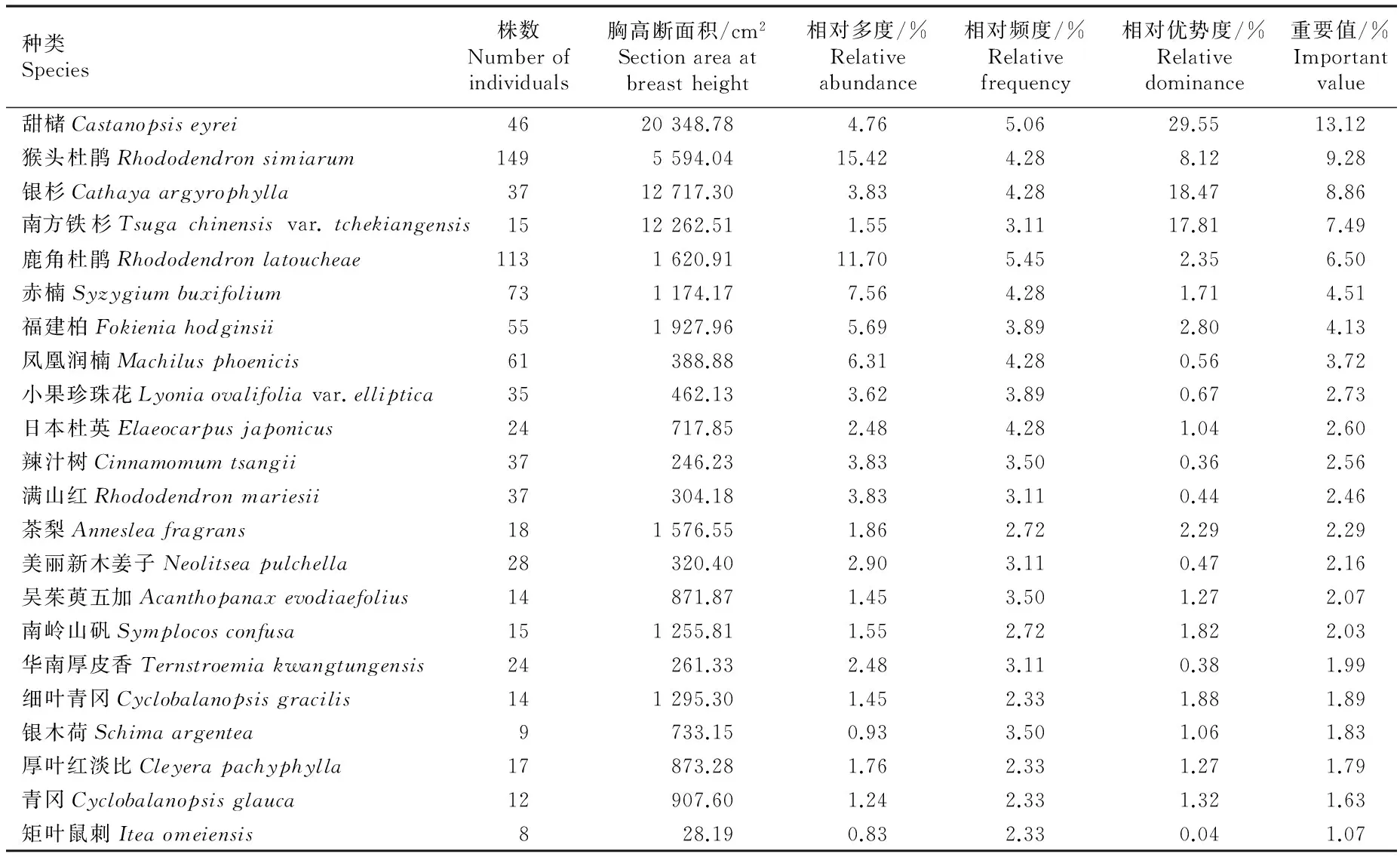
种类Species株数Numberofindividuals胸高断面积/cm2Sectionareaatbreastheight相对多度/%Relativeabundance相对频度/%Relativefrequency相对优势度/%Relativedominance重要值/%Importantvalue甜槠Castanopsiseyrei4620348.784.765.0629.5513.12猴头杜鹃Rhododendronsimiarum1495594.0415.424.288.129.28银杉Cathayaargyrophylla3712717.303.834.2818.478.86南方铁杉Tsugachinensisvar.tchekiangensis1512262.511.553.1117.817.49鹿角杜鹃Rhododendronlatoucheae1131620.9111.705.452.356.50赤楠Syzygiumbuxifolium731174.177.564.281.714.51福建柏Fokieniahodginsii551927.965.693.892.804.13凤凰润楠Machilusphoenicis61388.886.314.280.563.72小果珍珠花Lyoniaovalifoliavar.elliptica35462.133.623.890.672.73日本杜英Elaeocarpusjaponicus24717.852.484.281.042.60辣汁树Cinnamomumtsangii37246.233.833.500.362.56满山红Rhododendronmariesii37304.183.833.110.442.46茶梨Annesleafragrans181576.551.862.722.292.29美丽新木姜子Neolitseapulchella28320.402.903.110.472.16吴茱萸五加Acanthopanaxevodiaefolius14871.871.453.501.272.07南岭山矾Symplocosconfusa151255.811.552.721.822.03华南厚皮香Ternstroemiakwangtungensis24261.332.483.110.381.99细叶青冈Cyclobalanopsisgracilis141295.301.452.331.881.89银木荷Schimaargentea9733.150.933.501.061.83厚叶红淡比Cleyerapachyphylla17873.281.762.331.271.79青冈Cyclobalanopsisglauca12907.601.242.331.321.63矩叶鼠刺Iteaomeiensis828.190.832.330.041.07
1)群落中重要值小于1%的物种被忽略Species with important value less than 1% in the community are omitted.

A: 1%-20%; B: 21%-40%; C: 41%-60%; D: 61%-80%; E: 81%-100%.
1: 湖南八面山脚盆辽银杉群落CathayaargyrophyllaChun et Kuang community at Jiaopenliao of Bamianshan in Hu’nan Province; 2: 江西三清山华东黄杉群落PseudotsugagausseniiFlous community in Sanqing Mountain of Jiangxi Province; 3: 广东南岭国家级自然保护区疏齿木荷+福建柏群落SchimaremotiserrataH. T. Chang+Fokieniahodginsii(Dunn) HenryetThomascommunityinNanlingNationalNature Reserve of Guangdong Province; 4: Raunkiaer标准频度图谱Raunkiaer’s standard frequency diagram.
图1 湖南八面山脚盆辽银杉群落与2个亚热带针阔混交林群落及Raunkiaer标准频度图谱的频度级对比
Fig. 1 Comparison on frequency class ofCathayaargyrophyllaChun et Kuang community at Jiaopenliao of Bamianshan in Hu’nan Province with that of two communities of mixed coniferous and broad-leaved mixed forest in subtropics and Raunkiaer’s standard frequency diagram
与江西三清山华东黄杉群落和广东南岭国家级自然保护区疏齿木荷+福建柏群落这2个亚热带地区的针阔混交林群落的频度级相比,脚盆辽银杉群落的频度结构优于华东黄杉群落,但较纬度偏低的疏齿木荷+福建柏群落的稳定性差,可能与脚盆辽银杉群落优势种具有较强的种内及种间竞争有关。
2.2 银杉种群年龄结构及群落优势种径级结构分析
2.2.1 银杉种群的年龄结构分析 湖南八面山脚盆辽银杉群落中银杉种群的年龄结构图见图2。由图2可以看出:脚盆辽银杉群落中年龄小于20 a的银杉有19株,为1~3年生幼树,其中胸径达到1.5 cm以上的幼树只有6株。该群落中银杉的成年树可存活240 a以上,但年龄80~180 a的银杉植株比例偏低,这一现象在其他区域的银杉群落中也存在[4],可见银杉在长成大树的过程中阻碍因素较多。银杉为喜阳树种,生长期短,光饱和点和光补偿点均较高,光照因素对其幼树的生长明显有利,并且其在成年阶段更需要充足的光照[24-26]。而脚盆辽银杉群落的郁闭度较高,且中下层乔灌木丰富,虽然林下层存在一定数量的银杉和福建柏幼苗,但林窗缺失,这在很大程度上影响了林下幼树的生长,导致银杉种群表现出中龄级壮树较少的格局。
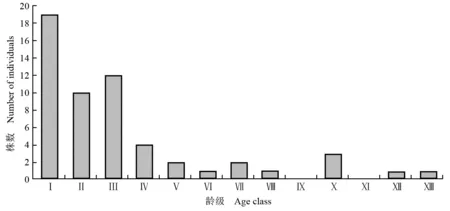
Ⅰ: 0-19 a; Ⅱ: 20-39 a; Ⅲ: 40-59 a; Ⅳ: 60-79 a; Ⅴ: 80-99 a; Ⅵ: 100-119 a; Ⅶ: 120-139 a; Ⅷ: 140-159 a; Ⅸ: 160-179 a; Ⅹ: 180-199 a; Ⅺ: 200-219 a; Ⅻ: 220-239 a;: ≥240 a.
图2 湖南八面山脚盆辽银杉群落中银杉的年龄结构图
Fig. 2 Diagram of age structure ofCathayaargyrophyllaChun et Kuang inC.argyrophyllacommunity at Jiaopenliao of Bamianshan in Hu’nan Province
2.2.2 群落优势种的径级结构分析 湖南八面山脚盆辽银杉群落中部分优势种(包括甜槠、猴头杜鹃、银杉、鹿角杜鹃、南方铁杉和福建柏)的径级结构见图3。由图3可以看出:在脚盆辽银杉群落中,银杉种群的年龄结构与径级结构基本一致。在脚盆辽银杉群落中,处于乔木第1亚层的高大乔木为银杉和南方铁杉,二者的胸径可达50.0 cm以上,常突出于群落林冠层,但这2个树种的幼龄个体较多,老树(胸径≥30.0 cm)个体极少,且中龄个体也较少;该群落中福建柏的径级结构也与二者相似。脚盆辽银杉群落中,在重要值最高的甜槠种群的径级结构中,壮年个体(20 cm≤胸径<40 cm)占据较高比例,数量明显多于老龄个体(胸径≥40 cm);胸径<5 cm的个体多达20余株,显示其幼树丰富度高,因此,甜槠种群为稳定增长型。
在脚盆辽银杉群落的乔木下层及灌木层中,杜鹃花科的猴头杜鹃和鹿角杜鹃占据明显优势,二者的径级结构也均呈增长状态,中龄个体的比例大于幼龄和老树个体,它们在乔木下层的生态位中占据主导地位。此外,在脚盆辽银杉群落中,银杉、南方铁杉和福建柏种群规模可能会随甜槠、猴头杜鹃和鹿角杜鹃种群的增长而逐步消退。

A: 甜槠Castanopsiseyrei(Champ.) Tutch.; B: 猴头杜鹃RhododendronsimiarumHance; C: 银杉CathayaargyrophyllaChun et Kuang; D: 鹿角杜鹃RhododendronlatoucheaeFranch.; E: 南方铁杉Tsugachinensisvar.tchekiangensis(Flous) Cheng et L. K. Fu; F: 福建柏Fokieniahodginsii(Dunn) Henry et Thomas.
图3 湖南八面山脚盆辽银杉群落优势种的径级结构图
Fig. 3 Diagram of diameter-class structure of dominant species inCathayaargyrophyllaChun et Kuang community at Jiaopenliao of Bamianshan in Hu’nan Province
2.3 不同区域银杉群落属的分布区类型比较及相似性分析
2.3.1 属的分布区类型比较 根据吴征镒等[19-20]对中国种子植物属分布区类型的划分原则,以群落为结构单元,对分布于湖南八面山的脚盆辽和小桃辽、广西大瑶山、湖南越城岭、贵州大娄山以及重庆金佛山的银杉群落(6个银杉群落分布地的基本概况见表2)中种子植物属的分布区类型进行比较,结果见表3。
由表3可知:6个银杉群落的种子植物属的分布区类型组成均体现出中亚热带植被区系成分交汇的性质。6个银杉群落中,大瑶山银杉群落处于最南部,但温带分布型属所占比例居中,推测与该群落所处的海拔较高以及山地黄壤土壤贫瘠有关;在纬度相对偏北的越城岭银杉群落中,热带分布型所占比例略高于温带分布型,分别为53.84%和46.16%,与该群落的海拔较低及所在区域降雨丰富有关;位于八面山脚盆辽和小桃辽的银杉群落纬度居中,其热带分布型属与温带分布型属的比例基本持平;在大娄山和金佛山银杉群落中,温带分布型所占比例分别为64.10%和72.73%,明显高于热带分布型。一般来说,随着纬度北移,温带分布型表现出逐渐增多的趋势,但从上述分析可见,银杉群落属的分布区类型除与纬度相关外,也受海拔的影响。
表2 分布于不同区域的6个银杉群落的基本概况
Table 2 Basic information of six communities ofCathayaargyrophyllaChun et Kuang distributing in different regions
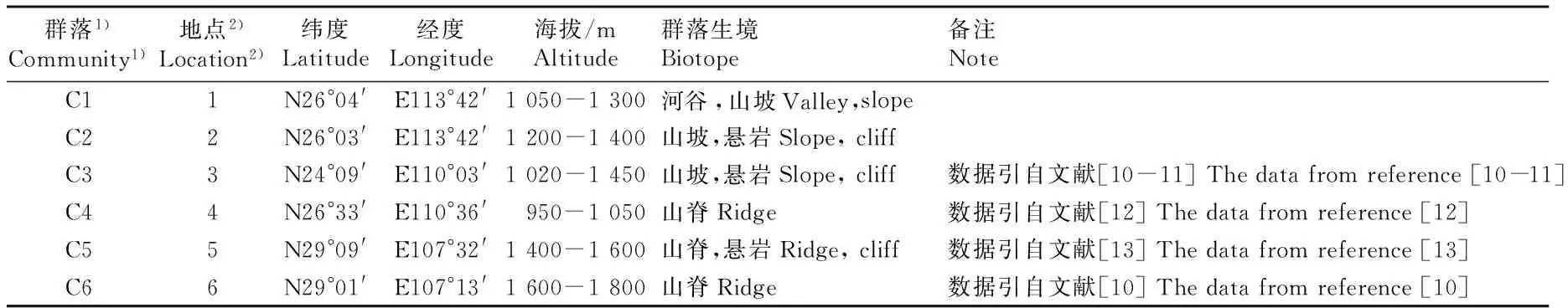
群落1)Community1)地点2)Location2)纬度Latitude经度Longitude海拔/mAltitude群落生境Biotope备注NoteC11N26°04'E113°42'1050-1300河谷,山坡Valley,slopeC22N26°03'E113°42'1200-1400山坡,悬岩Slope,cliffC33N24°09'E110°03'1020-1450山坡,悬岩Slope,cliff数据引自文献[10-11]Thedatafromreference[10-11]C44N26°33'E110°36'950-1050山脊Ridge数据引自文献[12]Thedatafromreference[12]C55N29°09'E107°32'1400-1600山脊,悬岩Ridge,cliff数据引自文献[13]Thedatafromreference[13]C66N29°01'E107°13'1600-1800山脊Ridge数据引自文献[10]Thedatafromreference[10]
1)C1: 脚盆辽 Jiaopenliao; C2: 小桃辽Xiaotaoliao; C3: 大瑶山Dayaoshan; C4: 越城岭Yuechengling; C5: 大娄山Daloushan; C6: 金佛山Jinfoshan.
2)1: 湖南八面山国家级自然保护区脚盆辽 Jiaopenliao of Bamianshan National Nature Reserve of Hu’nan Province; 2: 湖南八面山国家级自然保护区小桃辽Xiaotaoliao of Bamianshan National Nature Reserve of Hu’nan Province; 3: 广西大瑶山国家级自然保护区Dayaoshan National Nature Reserve of Guangxi Province; 4: 湖南越城岭Yuechengling of Hu’nan Province; 5: 贵州大沙河省级自然保护区大娄山Daloushan of Dashahe Provincial Nature Reserve of Guizhou Province; 6: 重庆金佛山国家级自然保护区Jinfoshan National Nature Reserve of Chongqing City.
表3 分布于不同区域的6个银杉群落中种子植物属的分布区类型1)
Table 3 Generic areal-type of spermatophytes in six communities ofCathayaargyrophyllaChun et Kuang distributing in different regions1)

分布区类型Areal-typeC1NP/%C2NP/%C3NP/%世界分布Cosmopolitan—2)—2)—2)泛热带分布Pantropic510.641121.15713.46热带亚洲和热带美洲间断分布Trop.Asia&Trop.Amer.disjuncted48.5123.8547.69旧世界热带分布OldWorldTropics12.1311.9211.92热带亚洲至热带大洋洲分布Trop.AsiatoTrop.Australasia12.1323.8523.85热带亚洲至热带非洲分布Trop.AsiatoTrop.Africa12.1300.0023.85热带亚洲(印度-马来西亚)分布Trop.Asia(Indo-Malesia)1123.401121.15917.31 热带分布型合计Totaloftropicareal-type2348.942751.922548.08北温带分布N.Temperate919.15611.54815.38东亚和北美洲间断分布E.Asia&N.Amer.disjuncted919.151121.15815.38旧世界温带分布OldWorldTemperate00.0011.9200.00东亚分布E.Asia510.6459.62713.46中国特有分布EndemictoChina12.1323.8547.69 温带分布型合计Totaloftemperateareal-type2451.062548.082751.92合计Total47100.0052100.0052100.00

分布区类型Areal-typeC4NP/%C5NP/%C6NP/%世界分布Cosmopolitan—2)—2)—2)泛热带分布Pantropic820.51717.9549.09热带亚洲和热带美洲间断分布Trop.Asia&Trop.Amer.disjuncted12.5625.1312.27旧世界热带分布OldWorldTropics25.1312.5600.00热带亚洲至热带大洋洲分布Trop.AsiatoTrop.Australasia25.1325.1312.27热带亚洲至热带非洲分布Trop.AsiatoTrop.Africa12.5600.0012.27热带亚洲(印度-马来西亚)分布Trop.Asia(Indo-Malesia)717.9525.13511.36 热带分布型合计Totaloftropicareal-type2153.841435.901227.26北温带分布N.Temperate512.821128.211227.27东亚和北美洲间断分布E.Asia&N.Amer.disjuncted820.51615.38920.45旧世界温带分布OldWorldTemperate00.0000.0000.00东亚分布E.Asia410.26410.261022.73中国特有分布EndemictoChina12.56410.2612.27 温带分布型合计Totaloftemperateareal-type1846.162564.103272.74合计Total39100.0039100.0044100.00
1)C1: 脚盆辽 Jiaopenliao; C2: 小桃辽Xiaotaoliao; C3: 大瑶山Dayaoshan; C4: 越城岭Yuechengling; C5: 大娄山Daloushan; C6: 金佛山Jinfoshan. N: 属数Number of genera; P: 各分布型所占比例Percentage of each areal-type.
2)未统计 Not counting.
2.3.2 属的分布区类型的相似性分析 植物区系格局与物种的形成过程和植被发展历史密切相关,6个银杉群落的演替历史可能与亚热带地区区系变迁进程相一致,从种子植物属的分布区类型的相似性系数(表4)看,脚盆辽、小桃辽、大瑶山和越城岭的银杉群落地处华南区系和华东南区系的交汇区,相似性系数均大于0.5;与地处北部的大娄山和金佛山银杉群落的相似性系数均小于0.5。从地理位置及区系区划来看,大娄山和金佛山银杉群落同位于贵州高原亚地区西北部,二者距离较接近,且所处纬度较高,因此与南部4个银杉群落区别较大;金佛山银杉群落的海拔高于大娄山银杉群落,金佛山银杉群落中落叶性植物明显多于后者,因此二者间的相似性系数也小于0.5。小桃辽和脚盆辽银杉群落同处于湖南八面山国家级自然保护区内, 在植物区划上属粤北亚地区[8], 这 2个群落的地理距离最近,它们的种子植物属分布区类型的相似性系数也最高,达到0.67;此外,这2个群落与地处粤桂山地亚地区的大瑶山和越城岭银杉群落的相似性系数也达0.55以上,反映出这4个银杉群落在植物区划上的一致性,即它们均属于岭南山地地区[8]。
尽管6个银杉群落在植物区划上属于3个不同的植物区系亚地区,但它们的种子植物属的分布区类型仍然有一定的相似性,相似性系数为0.35~0.49,且南部4个群落的相似性系数则达到0.55~0.67(表4),表明银杉群落虽经历漫长的演替历程而成为残存群落,但在属级水平上它们的分布区类型组成仍具有较高的相似度,特别是分布区类型亚型在地理区上极为相似。从表3可见:6个银杉群落种子植物属的主要分布区类型的优势度基本相似,如泛热带分布型、热带亚洲分布型、北温带分布型、东亚和北美洲间断分布型及东亚分布型等,仅各分布型所占比例有差异,其中,金佛山银杉群落中东亚分布型占22.73%,明显高于另5个银杉群落(仅占10.26%~13.46%),推测主要原因是金佛山山体高峻,亚热带山地成分特别丰富,具有明显的东亚区域特征,如双盾木属(DipeltaMaxim.)、 红果树属(StranvaesiaLindl.)、 吊钟花属(EnkianthusLour.)和石椒草属(BoenninghauseniaReichenbach ex Meisner)等东亚分布型属的大量分布。
表4 分布于不同区域的6个银杉群落中种子植物属分布区类型的相似性系数1)
Table 4 Similarity coefficient of generic areal-type of spermatophytes in six communities ofCathayaargyrophyllaChun et Kuang distributing in different regions1)

群落Community6个群落间的相似性系数SimilaritycoefficientamongsixcommunitiesC1C2C3C4C5C6C11.000.670.670.560.490.42C21.000.560.590.460.35C31.000.550.420.40C41.000.460.43C51.000.46C61.00
1)C1: 脚盆辽 Jiaopenliao; C2: 小桃辽Xiaotaoliao; C3: 大瑶山Dayaoshan; C4: 越城岭Yuechengling; C5: 大娄山Daloushan; C6: 金佛山Jinfoshan.
从6个银杉群落的优势种来看,银杉群落伴生种以松科的南方铁杉、长苞铁杉(TsugalongibracteataCheng)和华南五针松(PinuskwangtungensisChun ex Tsiang),樟科(Lauraceae)的润楠属(MachilusNees)和新木姜子属(NeolitseaMerr.)种类,壳斗科的水青冈属(FagusLinn.)、柯属(LithocarpusBlume)、青冈属(CyclobalanopsisOerst.)和鹅耳枥属(CarpinusLinn.)种类,山茶科的厚皮香属(TernstroemiaMutis ex Linn. f.)和柃木属(EuryaThunb.)种类,冬青科(Aquifoliaceae)的冬青属(IlexLinn.)种类,槭树科(Aceraceae)的槭属(AcerLinn.)种类,杜鹃花科的杜鹃属(RhododendronLinn.)种类为主;其他伴生种有青檀属(PteroceltisMaxim.)、蜡瓣花属(CorylopsisSieb. et Zucc.)、马蹄荷属(ExbucklandiaR. W. Brown)和含笑属(MicheliaLinn.)种类等,这些植物在第三纪大多已广泛存在,具有明显的孑遗性质[2]。化石研究结果[27]表明:在第三纪银杉群落的生存环境较温暖,随后在冰川作用下其分布区逐步退缩,现代的银杉群落均分布于山脊或山坡中部等光照充足的阳坡,其中,脚盆辽及大瑶山等地的银杉群落还分布在温暖湿润的河谷、山坡地区,这类生境与其在地质演化时期的生境相似。
3 讨论和结论
上述研究结果表明:湖南八面山国家级自然保护区是银杉分布的东界,该保护区内脚盆辽银杉群落中共有维管植物27科50属66种,种子植物属的分布区类型以温带分布型稍占优势,与其地处亚热带气候区相一致;群落的主要优势种为松科、杜鹃花科和壳斗科的种类,建群种为甜槠、银杉和南方铁杉,主要伴生种类有猴头杜鹃、鹿角杜鹃、福建柏和赤楠等。
脚盆辽银杉群落的物种多样性水平和Pielou均匀度指数均较高,与其具有典型的亚热带针阔叶混交林的特征相符[22],但该群落的物种多样性程度低于典型的亚热带常绿阔叶林[28]。群落中各生活型频度百分比由高到低依次为A级、B级、C级、D级、E级,E级树种仅甜槠和鹿角杜鹃,而高大乔木银杉和南方铁杉的频度级分别为D级和C级,表明脚盆辽银杉群落呈多优势种的格局。从径级结构上看,6个建群种中,甜槠、猴头杜鹃和鹿角杜鹃的种群结构为稳定型,在群落中处于优势地位;银杉、南方铁杉和福建柏种群中老龄个体占据群落的上层生态位,林下层郁闭度高的区域幼龄个体极少,而在灌木较少且光照较充足的坡地则生长有一定数量的银杉及南方铁杉幼龄个体,但它们的中龄个体较少,因此,推测林下层较高的郁闭度阻碍了这些种类的幼树成长为大树。
银杉在生命周期中可多次结实并产生种子,其花粉活力高、适宜于长距离传粉[29],在一定程度上证实银杉这一濒危植物的种子库具有稳定性,并在漫长的演替过程中得到延续。但银杉传粉期在雨季,且结实周期长达3年,加之动物对其种子的破坏等,这些不利因素均阻碍了银杉种群的扩张[18]。对脚盆辽银杉群落的实地考察结果表明:该群落中有3株银杉大树处于结实期,林下层也有较多的落果及银杉幼苗,但群落郁闭度太高,导致银杉幼树和幼苗死亡率高[9],限制了银杉种群的更新和发展。银杉作为一种长龄树种,适于生长在山脊和悬岩等光线充足的生境中,而在林下其幼苗的竞争力降低,因此,对银杉幼苗周围树木进行适当间伐,形成人为林窗,可在一定程度上促进银杉种群复壮。
植物群落作为具有特定特征的生态系统的主体,是物种与环境长期相互演化的结果,符合植物区系研究的整体性及自然性原则[30]。植物群落的演替具有明显的延续性和保守性,群落物种的组成存在进化的滞后性及生态位的保守性[31];对南半球45个生物区[32]以及东亚和北美维管植物[33]的研究结果也证实了大尺度生物区格局下植物群落的这种保守性;对桫椤〔Alsophilaspinulosa(Wall. ex Hook.) R. M. Tryon〕群落区系组成的研究结果也表明在群落水平上植被进化的保守性依然存在[34]。银杉作为东亚植物区的重要代表种,群落内伴生物种多具有古老历史,因而以银杉为特征种的植物群落在物种组成上可能体现着东亚植物区系的孑遗性及古老性。对6个银杉群落物种组成的比对结果显示:各群落在种子植物属的分布区类型组成上呈现出较高的一致性,相似性系数多在0.5以上,而组成成分的差异主要表现在林下草本以及群落的偶见种;种子植物属的分布区类型也体现着银杉群落的古老性,如各银杉群落的优势种、建群种多为泛热带分布的冬青属和厚皮香属,热带亚洲分布的含笑属、福建柏属(FokieniaHenry et Thomas)、青冈属和润楠属,北温带分布的槭属、水青冈属和杜鹃属,东亚和北美洲间断分布的柯属、铁杉属〔Tsuga(Endlicher) Carrière〕和锥属〔Castanopsis(D. Don) Spach〕,以及东亚分布的蜡瓣花属,这些属均起源古老,在第三纪大多已在中国大陆广布,其中部分属为著名的孑遗属[8,19]。
对欧洲奥地利晚中新世古生物群的研究结果[35]表明:银杉属的孢粉在第三纪沉积物中广泛存在,同地层的孢粉组合类型以木兰科(Magnoliaceae)、樟科和壳斗科类群占优势,第三纪银杉的生存环境表现出温暖湿润的特点,与其现代中亚热带暖性针阔叶混交林的生存环境一致。在更新世冰期中随气候变冷,分布于较寒冷地区的银杉均已灭绝[13],因而,银杉现仅残存于中国气候较温暖的亚热带地区;对银杉的谱系地理学研究结果表明:银杉的现存分布地在第四纪冰川时期为其避难所[3]。此外,银杉群落的伴生种华南五针松的冰期避难所也存在于大瑶山和越城岭等南岭山脉附近的中亚热带山地[36]。总体上看,银杉群落不仅在现生环境上与第三纪以来的古老生境明显相似,在群落的种系组成上也表现出古老性和孑遗性,可认为其在群落演替、物种进化和环境适应性上也存在明显的保守性,即银杉群落的演替进程与东亚植物区系的发展历史相一致,各地的现存银杉群落在第三纪晚期已经存在,随后因冰期气候波动等原因导致群落中的物种组成发生一定分化。
致谢: 湖南八面山国家级自然保护区工作人员、湖南师范大学刘克明教授在野外考察中给予大力支持,中山大学2014级本科生仇博元、陈冬夏、邵千芊、揭敏文、谢雁、湛霞和林心怡等参加了野外考察,中山大学植物标本馆助理王晓阳协助制作群落分布图,在此一并致谢!
参考文献:
[1] CHUN W Y, KUANG K Z. A new genus of Pinaceae,CathayaChun et Kuang, General Nov, from southern and western China[J]. Botaniceskii Žurnal SSSR, 1958, 43: 461-470.
[2] 周浙昆, MOMOHARA A. 一些东亚特有种子植物的化石历史及其植物地理学意义[J]. 云南植物研究, 2005, 27(5): 449-470.
[3] WANGHW,GES. Phylogeography of the endangeredCathayaargyrophylla(Pinaceae) inferred from sequence variation of mitochondrial and nuclear DNA[J]. Molecular Ecology, 2006, 15: 4109-4122.
[4] QIAN S H, YANG Y C, TANG C Q, et al. Effective conservation measures are needed for wildCathayaargyrophyllapopulations in China: insights from the population structure and regeneration characteristics[J]. Forest Ecology and Management, 2016, 361: 358-367.
[5] 祁承经, 肖育檀. 湖南省八面山银杉林的群落学分析[J]. 植物研究, 1988, 8(4): 169-182.
[6] 贺军辉. 八面山银杉林的调查研究[J]. 湖南林业科技, 1991(1): 42-45.
[7] 谢宗强, 陈伟烈, 江明喜, 等. 八面山银杉林种群的初步研究[J]. 植物学报, 1995, 37(1): 58-65.
[8] 陈灵芝. 中国植物区系与植被地理[M]. 北京: 科学出版社, 2014: 1-573.
[9] 谢宗强, 陈伟烈. 濒危植物银杉的群落特征及其演替趋势[J]. 植物生态学报, 1999, 23(1): 48-55.
[10] 应俊生, 马成功, 李良千, 等. 中国银杉林的初步研究[J]. 植物学报, 1983, 25(2): 157-170.
[11] 莫新礼, 钟业聪, 李法炎, 等. 广西大瑶山的银杉研究[J]. 广西植物, 1992, 12(3): 254-268.
[12] 祁承经, 曹铁如, 罗仲春. 湖南省越城岭北部罗汉洞的银杉与长苞铁杉混交林[J]. 植物生态学与地植物学丛刊, 1983, 7(1): 58-66.
[13] 李爱民. 大沙河自然保护区银杉种群和群落生态学研究[D]. 贵阳: 贵州大学林学院, 2011.
[14] 王伯荪, 余世孝, 彭少麟, 等. 植物群落学实验手册[M]. 广州: 广东高等教育出版社, 1996: 24-27.
[15] 方精云, 王襄平, 沈泽昊, 等. 植物群落清查的主要内容、方法和技术规范[J]. 生物多样性, 2009, 17(6): 533-548.
[16] PIELOU E C. 数学生态学引论[M]. 卢泽愚, 译. 北京: 科学出版社, 1978: 1-200.
[17] RAUNKIAER C. The Life Forms of Plants and Statistical Plant Geography[M]. Oxford: Oxford University Press, 1934: 623.
[18] 谢宗强, 陈伟烈, 路 鹏, 等. 濒危植物银杉的种群统计与年龄结构[J]. 生态学报, 1999, 19(4): 523-528.
[19] 吴征镒. 中国种子植物属的分布区类型[J]. 云南植物研究, 1991, 13(增刊Ⅳ): 1-179.
[20] 吴征镒, 孙 航, 周浙昆, 等. 中国种子植物区系地理[M]. 北京: 科学出版社, 2010: 1-485.
[21] 张镱锂, 张雪梅. 植物区系地理研究中的重要参数: 相似性系数[J]. 干旱区研究, 1998, 15(1): 59-63.
[22] 郭 微, 沈如江, 吴金火, 等. 江西三清山华东黄杉群落的组成及结构分析[J]. 植物资源与环境学报, 2007, 16(3): 46-52.
[23] 陈 林, 董安强, 王发国, 等. 广东南岭国家级自然保护区疏齿木荷+福建柏群落结构与物种多样性研究[J]. 热带亚热带植物学报, 2010, 18(1): 59-67.
[24] 张向峰, 王玉杰, 王云琦, 等. 银杉的光合特性[J]. 华北农学报, 2011, 26(增刊): 75-79.
[25] 张旺锋, 樊大勇, 谢宗强, 等. 濒危植物银杉幼树对生长光强的季节性光合响应[J]. 生物多样性, 2005, 13(5): 387-397.
[26] 谢宗强. 银杉(Cathayaargyrophylla)林林窗更新的研究[J]. 生态学报, 1999, 19(6): 775-779.
[27] LIU Y S, BASINGER J F. FossilCathaya(Pinaceae) pollen from the Canadian high arctic[J]. International Journal of Plant Sciences, 2000, 161: 829-847.
[28] 王伯荪, 彭少麟. 鼎湖山森林群落分析: Ⅶ. 生态优势度[J]. 中山大学学报, 1986(2): 93-97.
[29] 王红卫, 邓辉胜, 谭海明, 等. 银杉花粉生命力及其变异[J]. 植物生态学报, 2007, 31(6): 1199-1204.
[30] 朱 华. 中国植物区系研究文献中存在的几个问题[J]. 云南植物研究, 2007, 29(5): 489-491.
[31] RICKLEFS R E, LATHAM R E. Intercontinental correlation of geographical ranges suggests stasis in ecological traits of relict genera of temperate perennial herbs[J]. The American Naturalist, 1992, 139: 1305-1321.
[32] CRISP M D, ARROYO M T K, COOK L G, et al. Phylogenetic biomeconservatismonaglobalscale[J]. Nature, 2009, 458: 754-758.
[33] QIAN H, RICKLEFS R E. Geographical distribution and ecological conservatism of disjunct genera of vascular plants in eastern Asia and eastern North American[J]. Journal of Ecology, 2004, 92: 253-265.
[34] 郝云庆, 江 洪, 余树全, 等. 桫椤植物群落区系进化保守性[J]. 生态学报, 2009, 29(8): 4102-4111.
[35] MELLER B, KOVAR-EDER J, ZETTER R. Lower Miocene leaf, palynomorph, and diaspore assemblages from the base of the lignite-bearing sequence in the opencast mine Oberdorf, N Voitsberg (Styria, Austria) as an indication of “Younger Mastixioid” vegetation[J]. Palaeontographica Abteilung B, 1999, 252: 123-179.
(责任编辑: 张明霞)
Analyses on community characteristics and its relict and conservation of Cathaya argyrophylla at Bamianshan in Hu’nan Province
SU Leyi1, ZHAO Wanyi1, ZHANG Jijun2, YANG Yupeng1, GUO Yuanfei3, FAN Qiang1, LIAO Wenbo1,①
(1. School of Life Sciences, Sun Yat-sen University, Guangzhou 510275, China; 2. College of Resource Environment and Tourism, Capital Normal University, Beijing 100048, China; 3. Administration Bureau of Bamianshan National Nature Reserve of Hu’nan Province, Guidong 423501, China),J.PlantResour. &Environ., 2016, 25(4): 76-86
In order to understand current survival status of Chinese endemic relic plantCathayaargyrophyllaChun et Kuang,C.argyrophylla+Tsugachinensisvar.tchekiangensis(Flous) Cheng et L. K. Fu+Castanopsiseyrei(Champ.) Tutch.-RhododendronsimiarumHance community located at Jiaopenliao of Bamianshan in Hu’nan Province was investigated, its community characteristics and population age structure were analyzed, and generic areal-type and its similarity coefficient of spermatophytes in six communities distributing in different regions were compared. On this basis, relic characteristics ofC.argyrophyllacommunity was discussed. The results show that there are 66 species of vascular plants inC.argyrophyllacommunity at Jiaopenliao, including 4 species of ferns and 62 species of spermatophytes (4 species of gymnosperm, 58 species of angiosperm). Main dominant species in arbor layer of the community areC.eyrei,R.simiarum,C.argyrophyllaandT.chinensisvar.tchekiangensis, with important values of 13.12%, 9.28%, 8.86% and 7.49%, respectively. Simpson and Shannon-Wiener diversity indexes of the community are 0.94 and 3.20, respectively, and Pielou evenness indexJswandJsiare 0.81 and 0.92, respectively. It means that species diversity of the community is lower, but species distributes well, and its characteristics is consistent with that of warm coniferous and broad-leaved mixed forest in mid-subtropical mountainous region. Based on life-form spectrum, percentage of frequency in the community from high to low in sequence is A class, B class, C class, D class, E class, which is consistent with Raunkiaer’s standard frequency diagram, it means that the community is totally in relatively stable subclimax state. Based on age structure ofC.argyrophyllapopulation, middle and young age individuals in population occupy a certain ratio, while ratio of middle age individuals with 80-180 a is lower. Based on diameter-class structure of dominant species in the community,C.eyrei,R.simiarumandRhododendronlatoucheaeFranch. populations are belonged to growing type, whileC.argyrophylla,T.chinensisvar.tchekiangensisandFokieniahodginsii(Dunn) Henry et Thomas populations are belonged to declining type. Based on generic areal-type of spermatophytes inC.argyrophyllacommunity at Jiaopenliao, temperate areal-type genera account for 51.06%, which is slightly higher than tropic areal-type genera (accounting for 48.94%), meaning that this region is located in the transition area of tropical and subtropical. Besides latitude, altitude also has a certain effect on areal-type elements. In sixC.argyrophyllacommunities located in three floristic subregions of North Guangdong, Guangdong and Guangxi mountainous region and Guizhou Plateau, similarity coefficient of generic areal-type of spermatophytes is 0.35-0.67, the genera containing constructive species in community have similarity and intercommunity among different communities, such asTsuga(Endlicher) Carrière,MachilusNees,FagusLinn. andFokieniaHenry et Thomas, etc., and most of them are relic and conservative elements, indicating that existing communities ofC.argyrophyllahave similar vicissitude history, the distribution area has obvious characteristics of subtropical mountain shelter, showing relict and conservation of its succession process. It is suggested that moderately exploring forest gap inC.argyrophyllacommunity and decreasing canopy density in forest understory will be beneficial to growth of young tree and keepingC.argyrophyllapopulation continuous renewal and succession.
Jiaopenliao of Bamianshan in Hu’nan Province;CathayaargyrophyllaChun et Kuang; community characteristics; areal-type; population age structure; relict
2016-03-10
国家科技基础性工作专项(2013FY111500); 2014年度广东省教育教学成果奖(高等教育)培育第二类项目; 2015年广东省本科高校教学质量与教学改革工程立项建设项目
苏乐怡(1996—),女,广东佛山人,本科,专业为生物科学。
①通信作者E-mail: lsslwb@mail.sysu.edu.cn
Q948.15; S791.19
A
1674-7895(2016)04-0076-11
10.3969/j.issn.1674-7895.2016.04.10
