不同淹水条件下河竹叶片光合生理特性的变化
2016-02-16李迎春郭子武陈双林岳永德刘玉芳杨清平
李迎春, 郭子武, 陈双林,①, 岳永德, 刘玉芳, 杨清平
(1. 国际竹滕中心, 北京100102; 2. 中国林业科学研究院亚热带林业研究所, 浙江 杭州 311400)
不同淹水条件下河竹叶片光合生理特性的变化
李迎春1,2, 郭子武2, 陈双林2,①, 岳永德1, 刘玉芳2, 杨清平2
(1. 国际竹滕中心, 北京100102; 2. 中国林业科学研究院亚热带林业研究所, 浙江 杭州 311400)
为阐明淹水条件下河竹(PhyllostachysrivalisH. R. Zhao)叶片的光合生理特性,设置对照组(CK,正常供水)及中度淹水(T1,水面高出基质表面5 cm)和重度淹水(T2,水面高出基质表面10 cm)处理组,对持续淹水360 d内2年生河竹幼苗叶片的光合气体交换参数、光响应曲线、光合特征参数、资源利用效率参数、光合色素含量及比值的变化进行了分析;在此基础上,探讨了河竹对淹水环境的适应性。结果表明:随淹水时间延长,2个处理组的多数光合生理参数呈“升高—降低—升高”的变化趋势,且多数参数在淹水90 d时达到峰值、在淹水270 d时达到谷值;T1组的各项光合生理参数总体上高于T2组。淹水30~180 d,与CK组相比,2个处理组的净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)、胞间CO2浓度(Ci)和叶面饱和水汽压亏缺(Vpdl)总体上降低,表观量子效率(AQY)显著降低;气孔限制值(Ls)升高,而光饱和点(LSP)和光补偿点(LCP)显著升高;T1组的最大净光合速率(Pmax)略降低,而T2组的Pmax值则显著降低;但2个处理组的水分利用效率(WUE)、表观CO2利用效率(CUE)和表观光能利用效率(LUE)却无明显变化。淹水270和360 d,与CK组相比,2个处理组的Pn、Tr、Gs、Ls和Vpdl值降低,Pmax、LSP、 LCP、 WUE、 CUE和LUE值显著降低,仅Ci值升高;T1组的AQY值略降低,而T2组的AQY值则显著降低。2个处理组的光响应曲线变化趋势与CK组相似,但随淹水时间延长,2个处理组的Pn值变幅不同,其中T1组的Pn值总体上高于T2组。淹水30和90 d,与CK组相比,T1组的叶绿素b(Chlb)和总叶绿素(Chl)含量总体上显著升高,而T2组的Chlb和Chl含量总体上显著降低;2个处理组的叶绿素a(Chla)和类胡萝卜素(Car)含量以及Chla/Chlb和Chl/Car值也显著降低。淹水180~360 d,仅2个处理组的Chla/Chlb值显著升高,而其他光合色素含量及比值总体上显著降低。综合分析结果显示:河竹叶片Pn值在淹水前期和中期下降的主要原因为气孔限制,而在淹水后期下降的主要原因为非气孔限制。河竹对淹水环境有较强耐受性,并对长期淹水有一定的适应性;此外,中度淹水条件下河竹叶片的光能转化效率、对光辐射的利用范围和对光变化的适应能力均高于重度淹水条件。
河竹; 淹水; 光合气体交换参数; 光合特征; 资源利用效率参数; 光合色素
光合作用是植物生长发育的能量和物质基础,对植物体内能量的吸收、固定、分配及转化具有重要作用[1]。在淹水胁迫条件下,耐淹植物叶片的光合速率和气孔导度等指标在短时间内呈下降趋势,而不耐淹植物叶片的光合速率和气孔导度等指标则长时间呈下降趋势[2-3]。气孔导度下降、表观量子效率下降、库尺寸减少及光合色素含量和组成比例改变等因素可能是光合速率下降的主要原因[4]。耐淹植物通常可以保持较高的光合速率、气孔导度和蒸腾速率,以维持较高的光合能力,因此,这些光合气体交换参数在一定程度上可以作为判定植物耐淹的指标[5]。气体交换能力是影响植物水分和物质循环(碳水化物及营养物质)的关键因子[6],许多植物在缺氧条件下首先表现为叶片气孔关闭、CO2向叶片扩散的阻力增强,继而相关光合酶活性降低,最终导致叶片光合能力下降[7]。然而,有些植物在淹水过程中气孔能够重新打开,使叶片净光合速率逐渐恢复[8]。光合色素不仅担负着光能吸收与转化的重要作用,而且能够在环境变化过程中通过动态调节各种光合色素间的比例保证光合系统的正常转运[9],但淹水胁迫条件下不同树种叶片光合色素含量及组成比例差异明显[10-12]。
河竹(PhyllostachysrivalisH. R. Zhao)隶属禾本科(Poaceae)刚竹属(PhyllostachysSieb. et Zucc.)水竹组(Sect.HeterocladaeZ. P. Wang),为复轴型混生竹种,主要分布在浙江、福建及广东等地,生于溪涧边或山沟旁,性喜水湿[13]。河竹的鞭根极为发达,其竹鞭韧皮部密生1圈肉眼可见的气孔,呈现出明显的耐淹植物特征[14]。刘玉芳等[15]发现,河竹通过改变根系的形态可塑性和生理可塑性适应淹水环境,并在长期淹水环境下自然生长和更新。目前,关于河竹的研究主要集中在淹水环境下其鞭根系统的生物量分配[15]、生理指标[16]及主要养分含量和积累量[17]等方面,基本上揭示了河竹鞭根对淹水环境的养分生理适应策略,但是对河竹叶片如何通过能量吸收、固定、分配和转化适应长期淹水胁迫的生理机制尚未明确。
通过植物叶片气体交换特征可以明确植物的光合特性和水分利用效率及其与环境之间的关系[18-20]。为此,作者对不同淹水条件下河竹叶片的光合气体交换参数、光响应曲线、光合特征参数、资源利用效率参数、光合色素含量及比值的变化规律进行了比较,初步探讨了淹水条件下河竹对光能过剩的保护机制及其对资源利用的能力,以期为河竹在江、河、湖库区等自然消落带生境中的应用提供基础研究数据。
1 研究地概况和研究方法
1.1 研究地概况
研究地位于浙江省临安市太湖源观赏竹种园内。该区域属中亚热带湿润季风气候,年均温15.4 ℃,极端最低温和极端最高温分别为-13.3 ℃和40.2 ℃,日均温大于10 ℃的活动积温约5 100 ℃,年日照时数1 850~1 950 h, 年均无霜期235 d, 年降水量1 250~ 1 600 mm,年均空气相对湿度在80%以上。
1.2 材料
于2012年2月在研究地内的河竹种苗林中挖取2年生小丛状竹苗,每株保留5或6个盘枝。选择规格基本一致〔地径(1.0±0.2) cm、株高(1.03±0.38) m〕的竹苗,将其移栽到上口径32 cm、下口径23 cm、高27 cm的加仑盆中,每盆栽植10株立竹;以V(红壤)∶V(细沙)=3∶1混合基质作为栽培基质,每盆装栽培基质约15 kg。移栽后采取正常喷灌及清除竹笋和杂草等日常管理措施。
1.3 方法
1.3.1 淹水处理方法 设置3个处理组:CK组(对照,正常供水),定期人工喷灌供水使栽培基质的相对含水率保持在85%左右;T1组(中度淹水),水面高出基质表面5 cm,即相对淹水深度约为竹苗地上部高度的5%;T2组(重度淹水),水面高出基质表面10 cm,即相对淹水深度约为竹苗地上部高度的10%。于2013年4月15日,将供试竹苗连盆置于长4.3 m、宽3.3 m、深0.5 m的水泥池中进行处理。每个处理组40盆,每盆视为1个重复。
1.3.2 光合气体交换参数的测定 在淹水30、90、180、270和360 d,每处理组随机选取3盆,在所有竹苗从顶部向下数第3和第4盘枝的成熟叶片中随机选取10枚叶片测定光合气体交换参数。在测定当日9:00至10:00,采用LI-6400便携式光合仪(美国LI-COR公司)测定叶片的净光合速率(Pn)、蒸腾速率(Tr)、胞间CO2浓度(Ci)、气孔导度(Gs)、空气CO2浓度(Ca)和叶面饱和水汽压亏缺(Vpdl);测定时,光量子通量密度(PPFD)设为1 000 μmol·m-2·s-1,气体流速为500 μmol·s-1。根据所测数据计算气孔限制值(Ls)、水分利用效率(WUE)、表观CO2利用效率(CUE)和表观光能利用效率(LUE)。
1.3.3 光响应曲线的测定 在淹水30、90、180、270和360 d,每处理组随机选取3盆,在所有竹苗从顶部向下数第3和第4盘枝的成熟叶片中随机选取10枚,充分活化后在测定当日9:00至10:00用LI-6400便携式光合仪自带的红蓝光源测定叶片光响应曲线。测定时,PPFD设为2 000、1 800、1 500、1 200、1 000、800、500、200、100、80、50、20和0 μmol·m-2·s-1,叶室CO2浓度设为370~390 mol·m-2·s-1,叶片温度设为25 ℃~30 ℃,空气相对湿度65%~75%。
1.3.4 光合色素含量的测定 在淹水30、90、180、270和360 d,分别采集上述所有样叶及周围成熟叶片,按处理组混匀;分别称取叶片0.5 g,采用改良浸提法[21]提取叶片的光合色素,并用UV-2100型分光光度计(上海美谱达仪器有限公司)测定浸提液在波长663、645和470 nm处的光密度值,重复测定3次。根据所测数据计算叶绿素a(Chla)、叶绿素b(Chlb)、总叶绿素(Chl)和类胡萝卜素(Car)的含量,并计算Chla/Chlb和Chl/Car值。
1.4 数据处理及统计分析
气孔限制值(Ls)、水分利用效率(WUE)、表观CO2利用效率(CUE)和表观光能利用效率(LUE)分别根据公式“Ls=1-Ci/Ca[22]”、“WUE=Pn/Tr[23]”、“CUE=Pn/Ci[24]”和“LUE=Pn/PPFD[25-26]”计算。采用最小二乘法对Pn与PPFD间的关系进行拟合分析,方程式为“Pn=Pmax·(1-C0·e-AQY·PPFD/Pmax)[27]”,式中,Pmax为最大净光合速率;AQY为表观量子效率;C0为度量弱光下净光合速率趋于0的指标。光补偿点(LCP)根据公式“LCP=Pmax·ln(C0)/AQY”计算;假定Pn达到Pmax的99%时PPFD为光饱和点(LSP),计算公式为“LSP=Pmax·ln(100C0)/AQY[28]”。
实验数据采用EXCEL 2003统计分析软件进行整理和制作图表;采用SPSS 20.0统计分析软件对相关数据进行单因素方差分析(one-way ANOVA),并采用Duncan’s新复极差法进行多重比较。
2 结果和分析
2.1 不同淹水条件下河竹叶片光合气体交换参数的变化
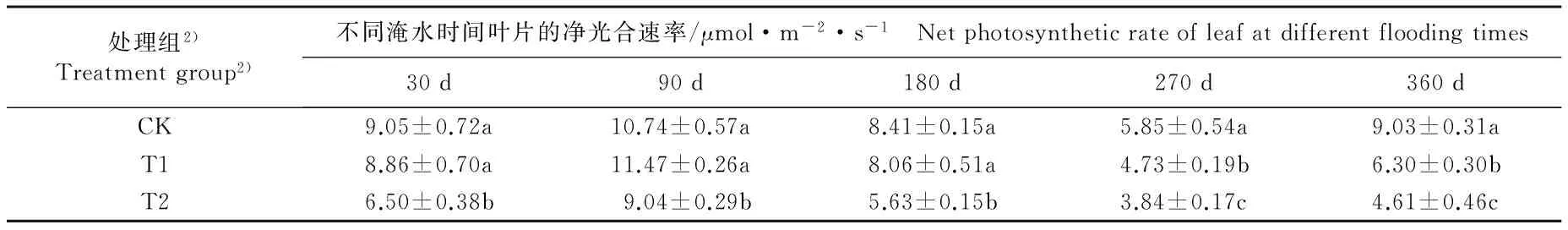
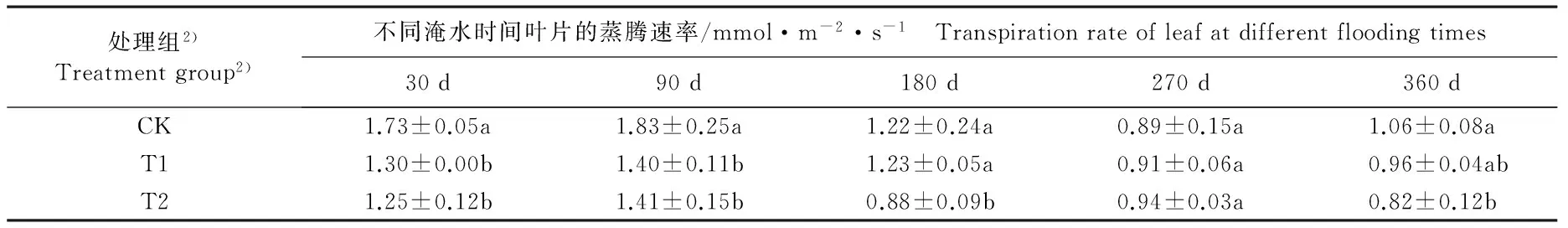
不同淹水条件下河竹叶片光合气体交换参数的变化见表1。由表1可以看出:随淹水时间的延长,各处理组河竹叶片的各项光合气体交换参数大多呈“升高—降低—升高”的变化趋势,且多数在淹水90 d时达到峰值、在淹水270 d时达到谷值。
淹水30 d, T1组(中度淹水,水面高出基质表面 5 cm)仅蒸腾速率(Tr)与CK组(对照组,正常供水)差异显著(P<0.05),且较CK组下降24.9%;其余光合气体交换参数与CK组均无显著差异(P>0.05)。T2组(重度淹水,水面高出基质表面10 cm)的净光合速率(Pn)、Tr值和气孔导度(Gs)分别较CK组下降28.2%、27.7%和30.8%,气孔限制值(Ls)较CK组升高19.4%,且差异均达到显著水平;其余光合气体交换参数与CK组无显著差异。
表1 不同淹水条件下河竹叶片光合气体交换参数的变化
Table 1 Changes in photosynthetic gas exchange parameters of leaf of Phyllostachys rivalis H.R.Zhao under different flooding conditions
处理组2)Treatmentgroup2)不同淹水时间叶片的净光合速率/μmol·m-2·s-1 Netphotosyntheticrateofleafatdifferentfloodingtimes30d90d180d270d360dCK9.05±0.72a10.74±0.57a8.41±0.15a5.85±0.54a9.03±0.31aT18.86±0.70a11.47±0.26a8.06±0.51a4.73±0.19b6.30±0.30bT26.50±0.38b9.04±0.29b5.63±0.15b3.84±0.17c4.61±0.46c
处理组2)Treatmentgroup2)不同淹水时间叶片的蒸腾速率/mmol·m-2·s-1 Transpirationrateofleafatdifferentfloodingtimes30d90d180d270d360dCK1.73±0.05a1.83±0.25a1.22±0.24a0.89±0.15a1.06±0.08aT11.30±0.00b1.40±0.11b1.23±0.05a0.91±0.06a0.96±0.04abT21.25±0.12b1.41±0.15b0.88±0.09b0.94±0.03a0.82±0.12b

处理组2)Treatmentgroup2)不同淹水时间叶片的胞间CO2浓度/μmol·mol-1 IntercellularCO2concentrationofleafatdifferentfloodingtimes30d90d180d270d360dCK256.72±4.88a 256.30±17.9a 250.10±10.26a 304.85±10.61a 243.07±2.61b T1260.74±11.12a241.40±6.40a245.25±13.19a304.83±0.02a301.23±8.97aT2223.59±15.32a234.77±21.02a217.30±11.78b305.10±4.36ab324.50±23.08a

处理组2)Treatmentgroup2)不同淹水时间叶片的气孔导度/mol·m-2·s-1 Stomatalconductanceofleafatdifferentfloodingtimes30d90d180d270d360dCK0.13±0.00a0.14±0.01a0.11±0.01a0.07±0.02b0.10±0.00aT10.13±0.00a0.12±0.01a0.10±0.02ab0.08±0.00a0.06±0.00bT20.09±0.00b0.12±0.01a0.07±0.02b0.05±0.02ab0.06±0.00b

处理组2)Treatmentgroup2)不同淹水时间叶片的气孔限制值 Limitedstomatalvalueofleafatdifferentfloodingtimes30d90d180d270d360dCK0.36±0.01b0.37±0.04a0.36±0.03b0.22±0.02a0.39±0.01aT10.35±0.03b0.40±0.01a0.38±0.03ab0.22±0.00a0.23±0.03bT20.43±0.03a0.42±0.06a0.45±0.03a0.22±0.01a0.17±0.05b
1)同列中不同的小写字母表示差异显著(P<0.05) Different small letters in the same column indicate the significant difference (P<0.05).
2)CK: 对照(正常供水) The control (normal water supplying); T1: 中度淹水(水面高出基质表面5 cm) Moderate flooding (water surface higher than substrate surface 5 cm); T2: 重度淹水(水面高出基质表面10 cm) Severe flooding (water surface higher than substrate surface 10 cm).
淹水90 d,T1组仅Tr值和叶面饱和水汽压亏缺(Vpdl)较CK组显著降低;T2组的Pn、Tr和Vpdl值较CK组显著降低。2个淹水处理组的其余光合气体交换参数与CK组无显著差异。
淹水180 d,T1组仅Vpdl值显著低于CK组;T2组的Pn值、Tr值、胞间CO2浓度(Ci)和Gs值显著低于CK组,Ls值显著高于CK组。2个淹水处理组的其余光合气体交换参数与CK组无显著差异。
淹水270 d,T1组的Pn和Vpdl值显著低于CK组,Gs值显著高于CK组;T2组的Pn和Vpdl值均显著低于CK组。2个淹水处理组的其余光合气体交换参数与CK组无显著差异。
淹水360 d,T1和T2组的Pn、Gs和Ls值以及T2组的Tr值均显著低于CK组,而Ci值显著高于CK组。其中,T1组的Pn、Gs和Ls值较CK组分别下降30.2%、40.0%和41.0%,T2组Pn、Gs、Ls和Tr值较CK组分别下降48.9%、40.0%、56.4%和22.6%,T1和T2组的Ci值较CK组分别升高23.9%和33.5%。2个淹水处理组的其余光合气体交换参数与CK组无显著差异。
由表1还可以看出:不同淹水时间T1和T2组河竹叶片的Pn、Tr、Ci、Gs、Ls和Vpdl 6项光合气体交换参数总体上低于CK组,且T2组叶片的上述6项光合气体交换参数总体上低于T1组。
2.2 不同淹水条件下河竹叶片光响应曲线和光合特征参数的变化
2.2.1 光响应曲线的变化 不同淹水胁迫下河竹叶片光响应曲线的变化见图1。由图1可以看出:在淹水处理的不同时间,随光量子通量密度(PPFD)的提高,各处理组河竹叶片净光合速率(Pn)的变化趋势相似,在PPFD低于100 μmol·m-2·s-1时叶片Pn值近直线升高,在PPFD高于1 000 μmol·m-2·s-1时叶片的Pn值趋于稳定,但在平衡状态下各组叶片的Pn值存在一定差异。

—●—: CK(对照, 正常供水 The control, normal water supplying); —■—: T1(中度淹水,水面高出基质表面5 cm Moderate flooding, water surface higher than substrate surface 5 cm); —▲—: T2(重度淹水,水面高出基质表面10 cm Severe flooding, water surface higher than substrate surface 10 cm).
A-E: 分别为淹水30、90、180、270和360 d时河竹叶片的光响应曲线 Representing light response curve of leaf ofP.rivaliswhen flooding for 30, 90, 180, 270 and 360 d, respectively.
图1 不同淹水条件下河竹叶片光响应曲线的变化
Fig. 1 Change in light response curve of leaf ofPhyllostachysrivalisH. R. Zhao under different flooding conditions
淹水30 d, T1组(中度淹水, 水面高出基质表面5 cm)叶片的光响应曲线与CK组(对照组,正常供水)基本一致,仅Pn值略低于CK组;T2组(重度淹水,水面高出基质表面10 cm)叶片的Pn值明显低于T1组和CK组。淹水90 d,T1组叶片的光响应曲线几乎与CK组完全重合,T2组的Pn值仍明显低于T1组和CK组。淹水180~360 d,随淹水时间的延长,T1组的Pn值与CK组的差异幅度越加明显,而与T2组的差异幅度趋小。
2.2.2 光合特征参数的变化 不同淹水条件下河竹叶片光合特征参数的变化见表2。由表2可以看出:随淹水时间的延长,各处理组河竹叶片的最大净光合速率(Pmax)和表观量子效率(AQY)均呈“升高—降低—升高”的变化趋势,并且均在淹水90 d时达到峰值、在淹水270 d时达到谷值;而叶片的光饱和点(LSP)和光补偿点(LCP)变化各异,但LCP值均在淹水270 d时达到谷值。
表2 不同淹水条件下河竹叶片光合特征参数的变化
Table 2 Changes in photosynthetic characteristic parameters of leaf of Phyllostachys rivalis H.R.Zhao under different flooding conditions

处理组2)Treatmentgroup2)不同淹水时间叶片的最大净光合速率/μmol·m-2·s-1 Themaximumnetphotosyntheticrateofleafatdifferentfloodingtimes30d90d180d270d360dCK9.834±1.370a11.527±0.181a8.908±0.477a6.124±0.131a9.219±0.544aT19.390±0.579a11.471±0.080a8.434±0.535a4.805±0.099b6.005±0.355bT26.263±0.692b8.994±0.221b5.880±0.090b3.721±0.105c5.603±0.331b

处理组2)Treatmentgroup2)不同淹水时间叶片的表观量子效率 Apparentquantumyieldofleafatdifferentfloodingtimes30d90d180d270d360dCK0.054±0.006a0.065±0.002a0.049±0.001a0.033±0.001b0.050±0.003aT10.051±0.006a0.051±0.001b0.043±0.001b0.037±0.001a0.046±0.003aT20.038±0.004b0.044±0.002c0.036±0.001c0.030±0.001c0.038±0.002b

处理组2)Treatmentgroup2)不同淹水时间叶片的光饱和点/μmol·m-2·s-1 Lightsaturationpointofleafatdifferentfloodingtimes30d90d180d270d360dCK1034.311±114.156a 893.651±21.906b872.827±13.377b 845.764±18.027a 901.410±53.234a T1939.704±103.714a1140.074±27.946a1085.229±16.633a636.482±13.566b627.499±37.058bT2893.753±98.642a621.649±15.238c518.437±7.940c364.429±7.767c341.649±20.176c

处理组2)Treatmentgroup2)不同淹水时间叶片的光补偿点/μmol·m-2·s-1 Lightcompensationpointofleafatdifferentfloodingtimes30d90d180d270d360dCK100.330±11.073a23.104±0.566c20.964±0.321c13.886±0.296a20.647±1.219aT178.319±8.644b112.667±2.762a79.906±1.225a6.801±0.145b7.464±0.441aT268.998±7.615b46.412±1.138b24.672±0.378b14.000±0.298a20.708±1.223b
1)同列中不同的小写字母表示差异显著(P<0.05) Different small letters in the same column indicate the significant difference (P<0.05).
2)CK: 对照(正常供水) The control (normal water supplying); T1: 中度淹水(水面高出基质表面5 cm) Moderate flooding (water surface higher than substrate surface 5 cm); T2: 重度淹水(水面高出基质表面10 cm) Severe flooding (water surface higher than substrate surface 10 cm).
淹水30 d,T1组仅LCP值显著低于CK组,T2组的Pmax、AQY和LCP值均显著低于CK组,2个处理组的其余光合特征参数与CK组无显著差异。淹水90和180 d,仅T1组的Pmax值与CK组无显著差异,T1和T2组的其余光合特征参数与CK组均有显著差异。淹水270 d,仅T2组LCP值与CK组无显著差异,T1和T2组其余光合特征参数与CK组均有显著差异。淹水360 d,T1组的Pmax和LCP值及T2组的4个光合特征参数均与CK组差异显著。
由表2还可以看出:不同淹水时间T1和T2组河竹叶片的Pmax、AQY、LSP和LCP 4项光合特征参数总体上低于CK组,且T2组叶片的上述4项光合特征参数总体上低于T1组。
2.3 不同淹水条件下河竹叶片资源利用效率参数的变化
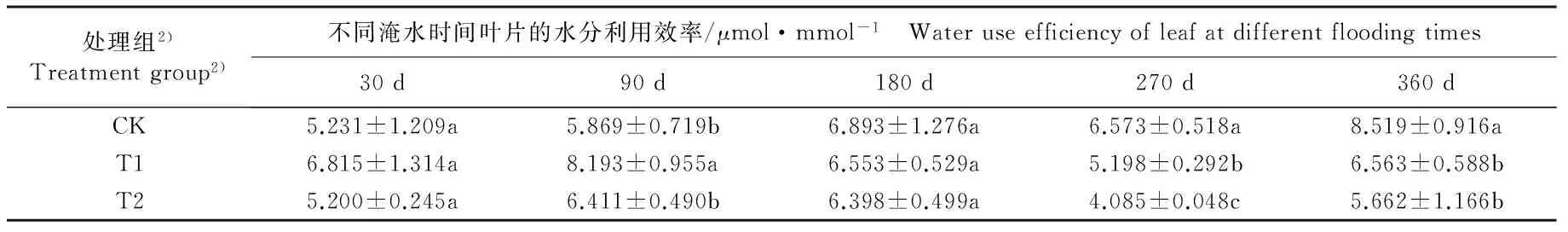
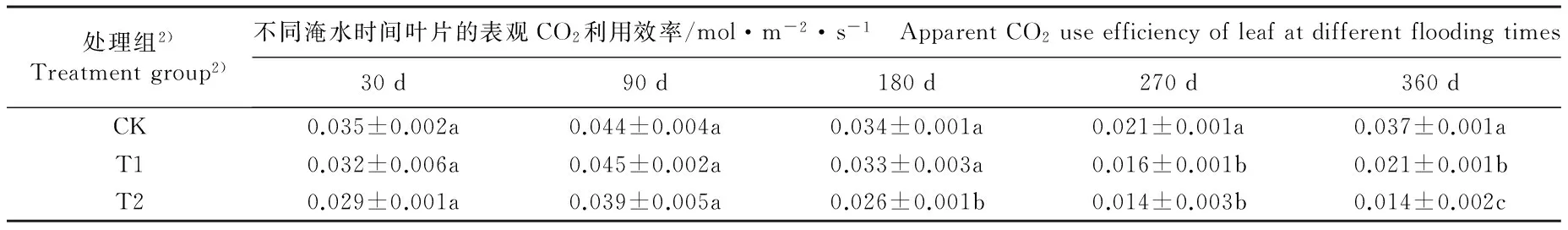
不同淹水条件下河竹叶片资源利用效率参数的变化见表3。由表3可以看出:随淹水时间延长,各处理组河竹叶片的水分利用效率(WUE)、表观CO2利用效率(CUE)和表观光能利用效率(LUE)均呈“升高—降低—升高”的变化趋势,并且均在淹水90 d时达到峰值,在淹水270 d时达到谷值。
淹水30 d,仅T2(重度淹水,水面高出基质表面10 cm)组的LUE值与CK组(对照组,正常供水)差异显著,T1(中度淹水,水面高出基质表面5 cm)和T2组的其余资源利用效率参数与CK组无显著差异。淹水90 d,T1组的WUE值和T2组LUE值与CK组差异显著,T1和T2组的其余资源利用效率参数与CK组无显著差异。淹水180 d,仅T2组的CUE和LUE值与CK组差异显著,T1和T2组的其余资源利用效率参数与CK组无显著差异。淹水270和360 d,T1和T2组的各项资源利用效率参数与CK组均有显著差异。至实验结束时(淹水360 d),T1和T2组的WUE值较CK组分别下降23.0%和34.0%,CUE值较CK组分别下降43.2%和62.2%,LUE值较CK组分别下降33.3%和44.4%。
由表3还可以看出:不同淹水时间T1和T2组河竹叶片的WUE、CUE和LUE 3项资源利用效率参数总体上低于CK组,且T2组叶片的上述3项资源利用效率参数总体上低于T1组。
表3 不同淹水条件下河竹叶片资源利用效率参数的变化
Table 3 Changes in resource use efficiency parameters of leaf of Phyllostachys rivalis H.R.Zhao under different flooding conditions
处理组2)Treatmentgroup2)不同淹水时间叶片的水分利用效率/μmol·mmol-1 Wateruseefficiencyofleafatdifferentfloodingtimes30d90d180d270d360dCK5.231±1.209a5.869±0.719b6.893±1.276a6.573±0.518a8.519±0.916aT16.815±1.314a8.193±0.955a6.553±0.529a5.198±0.292b6.563±0.588bT25.200±0.245a6.411±0.490b6.398±0.499a4.085±0.048c5.662±1.166b
处理组2)Treatmentgroup2)不同淹水时间叶片的表观CO2利用效率/mol·m-2·s-1 ApparentCO2useefficiencyofleafatdifferentfloodingtimes30d90d180d270d360dCK0.035±0.002a0.044±0.004a0.034±0.001a0.021±0.001a0.037±0.001aT10.032±0.006a0.045±0.002a0.033±0.003a0.016±0.001b0.021±0.001bT20.029±0.001a0.039±0.005a0.026±0.001b0.014±0.003b0.014±0.002c
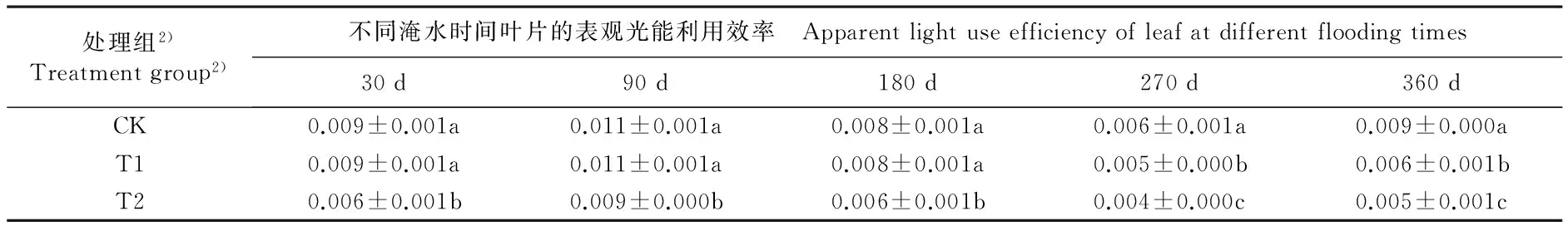
处理组2)Treatmentgroup2)不同淹水时间叶片的表观光能利用效率 Apparentlightuseefficiencyofleafatdifferentfloodingtimes30d90d180d270d360dCK0.009±0.001a0.011±0.001a0.008±0.001a0.006±0.001a0.009±0.000aT10.009±0.001a0.011±0.001a0.008±0.001a0.005±0.000b0.006±0.001bT20.006±0.001b0.009±0.000b0.006±0.001b0.004±0.000c0.005±0.001c
1)同列中不同的小写字母表示差异显著(P<0.05) Different small letters in the same column indicate the significant difference (P<0.05).
2)CK: 对照(正常供水) The control (normal water supplying); T1: 中度淹水(水面高出基质表面5 cm) Moderate flooding (water surface higher than substrate surface 5 cm); T2: 重度淹水(水面高出基质表面10 cm) Severe flooding (water surface higher than substrate surface 10 cm).
2.4 不同淹水条件下河竹叶片光合色素含量及其比值的变化
不同淹水条件下河竹叶片光合色素含量及其比值的变化见表4。由表4可以看出:不同淹水时间,各处理组河竹叶片的叶绿素a(Chla)、叶绿素b(Chlb)、类胡萝卜素(Car)和总叶绿素(Chl)含量及Chla/Chlb和Chl/Car值的变化均存在一定的差异。
淹水30 d, T1组叶片的Chla、 Chlb、 Car和Chl含量分别较CK组升高9.0%、 20.4%、 18.8%和11.5%,其中Chlb、Car和Chl含量与CK组差异显著;而T2组叶片的这4项指标均与CK组无显著差异。淹水90 d,T1组叶片的Chla和Car含量较CK组分别下降27.6%和30.3%, 且Chlb含量较CK组升高55.2%, 且差异均达到显著水平, 而T2组叶片的这 3项指标与CK组无显著差异;T1和T2组叶片的Chl含量均高于CK组,但均无显著差异。淹水180~360 d,T1和T2组叶片的Chla、Chlb、Car和Chl含量基本上显著低于CK组。
由表4还可以看出:淹水30 d,T1和T2组河竹叶片的Chla/Chlb和Chl/Car值均显著低于CK组。淹水90 d,T1组叶片的Chla/Chlb值显著小于CK组,而Chl/Car值显著大于CK组;T2组叶片的Chla/Chlb和Chl/Car值与CK组无显著差异。淹水180 d,T1和T2组叶片的Chla/Chlb值显著大于CK组,而Chl/Car值却显著小于CK组。淹水270 d,仅T1组叶片的Chl/Car值与CK组无显著差异,T1和T2组叶片的Chla/Chlb值以及T2组叶片的Chl/Car值均显著大于CK组。淹水360 d,仅T1组叶片的Chl/Chlb值与CK组无显著差异,T2组叶片的Chla/Chlb值以及2个处理组叶片的Chl/Car值均显著小于CK组。
表4 不同淹水条件下河竹叶片光合色素含量及比值的变化
Table 4 Changes in contents and ratios of photosynthetic pigments in leaf of Phyllostachys rivalis H.R.Zhao under different flooding conditions
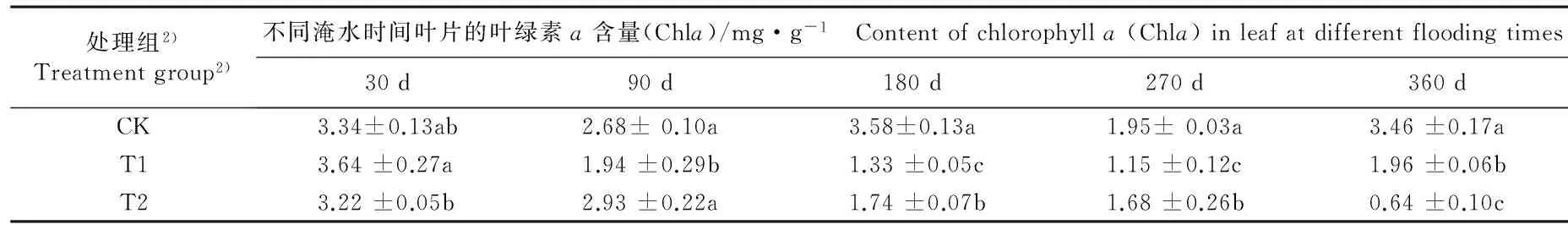
处理组2)Treatmentgroup2)不同淹水时间叶片的叶绿素a含量(Chla)/mg·g-1 Contentofchlorophylla(Chla)inleafatdifferentfloodingtimes30d90d180d270d360dCK3.34±0.13ab2.68±0.10a3.58±0.13a1.95±0.03a3.46±0.17aT13.64±0.27a1.94±0.29b1.33±0.05c1.15±0.12c1.96±0.06bT23.22±0.05b2.93±0.22a1.74±0.07b1.68±0.26b0.64±0.10c
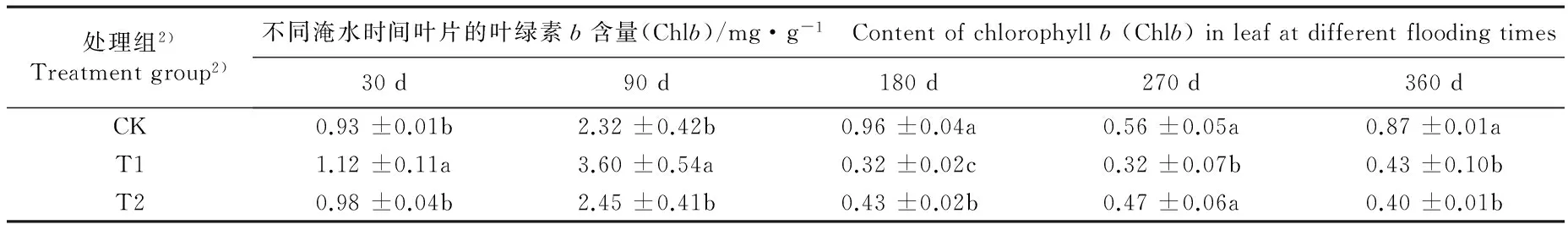
处理组2)Treatmentgroup2)不同淹水时间叶片的叶绿素b含量(Chlb)/mg·g-1 Contentofchlorophyllb(Chlb)inleafatdifferentfloodingtimes30d90d180d270d360dCK0.93±0.01b2.32±0.42b0.96±0.04a0.56±0.05a0.87±0.01aT11.12±0.11a3.60±0.54a0.32±0.02c0.32±0.07b0.43±0.10bT20.98±0.04b2.45±0.41b0.43±0.02b0.47±0.06a0.40±0.01b
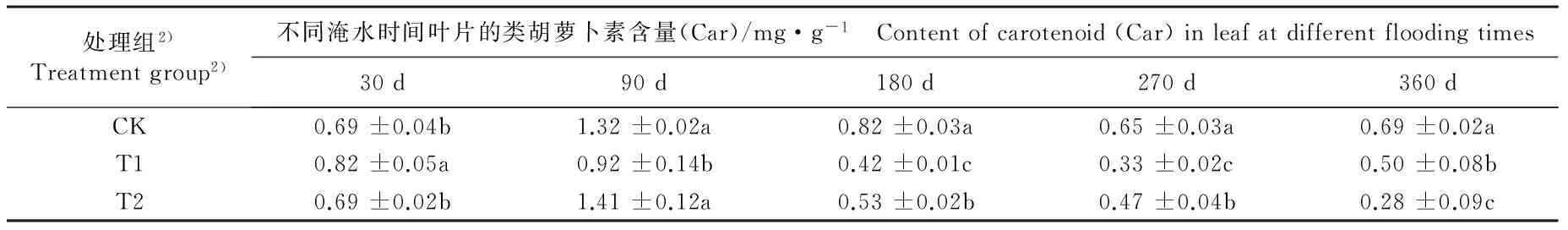
处理组2)Treatmentgroup2)不同淹水时间叶片的类胡萝卜素含量(Car)/mg·g-1 Contentofcarotenoid(Car)inleafatdifferentfloodingtimes30d90d180d270d360dCK0.69±0.04b1.32±0.02a0.82±0.03a0.65±0.03a0.69±0.02aT10.82±0.05a0.92±0.14b0.42±0.01c0.33±0.02c0.50±0.08bT20.69±0.02b1.41±0.12a0.53±0.02b0.47±0.04b0.28±0.09c
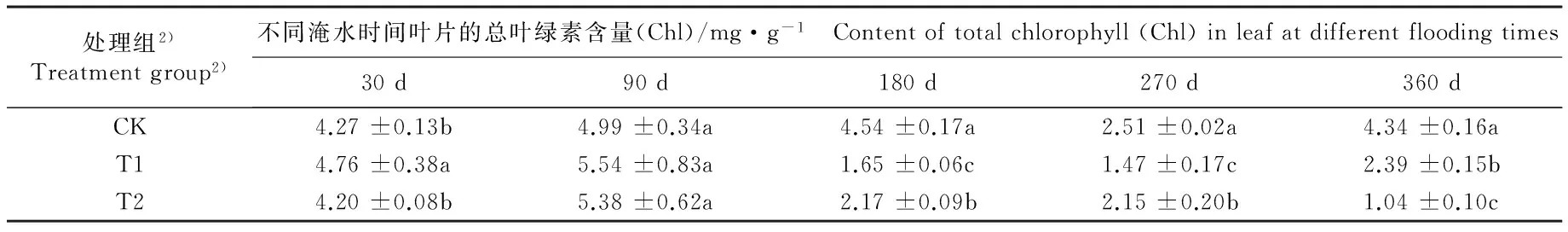
处理组2)Treatmentgroup2)不同淹水时间叶片的总叶绿素含量(Chl)/mg·g-1 Contentoftotalchlorophyll(Chl)inleafatdifferentfloodingtimes30d90d180d270d360dCK4.27±0.13b4.99±0.34a4.54±0.17a2.51±0.02a4.34±0.16aT14.76±0.38a5.54±0.83a1.65±0.06c1.47±0.17c2.39±0.15bT24.20±0.08b5.38±0.62a2.17±0.09b2.15±0.20b1.04±0.10c

处理组2)Treatmentgroup2)不同淹水时间叶片的Chla/Chlb值 Chla/Chlbvalueinleafatdifferentfloodingtimes30d90d180d270d360dCK3.60±0.12a1.16±0.23a3.71±0.02b3.45±0.12b3.96±0.25aT13.24±0.08b0.54±0.00b4.10±0.16a3.59±0.33a4.08±0.14aT23.30±0.09b1.20±0.11a4.09±0.03a3.60±0.37a1.61±1.61b

处理组2)Treatmentgroup2)不同淹水时间叶片的Chl/Car值 Chl/Carvalueinleafatdifferentfloodingtimes30d90d180d270d360dCK6.20±0.18a3.78±0.25b5.55±0.04a3.85±0.13b6.32±0.13aT15.84±0.10b6.03±0.12a3.92±0.06c4.43±0.25ab4.77±0.55bT26.11±0.17ab3.82±0.12b4.12±0.03b4.59±0.54a3.76±0.46b
1)同列中不同的小写字母表示差异显著(P<0.05) Different small letters in the same column indicate the significant difference (P<0.05).
2)CK: 对照(正常供水) The control (normal water supplying); T1: 中度淹水(水面高出基质表面5 cm) Moderate flooding (water surface higher than substrate surface 5 cm); T2: 重度淹水(水面高出基质表面10 cm) Severe flooding (water surface higher than substrate surface 10 cm).
3 讨论和结论
净光合速率(Pn)能够直接反映植物的光合作用能力,其降幅可用于衡量植物对逆境条件的光合响应能力[29-31]。贾中民等[25]的研究结果表明:耐淹能力较强的树种其叶片的Pn值在淹水初期有所下降,但在淹水后期逐步趋于稳定。在淹水深度5 cm的条件下,耐淹能力较强的树种枫杨(PterocaryastenopteraC. DC.)幼苗在持续淹水1 a后,其叶片的Pn值降为对照的54.16%[25];而不耐淹的树种栓皮栎(QuercusvariabilisBl.)的幼苗在根部淹水时间短于50 d情况下,其叶片的Pn值降至对照的0.00%~25.00%[10]。本研究中,随淹水时间延长,T1(中度淹水,水面高出基质表面5 cm)和T2(重度淹水,水面高出基质表面10 cm)组河竹叶片的Pn值均呈“升高—降低—升高”的变化趋势,并在淹水90 d时达到峰值、在淹水270 d时达到谷值;而淹水时间达到270和360 d,T1和T2组叶片的Pn值仍能够达到CK组(对照组,正常供水)的50%以上。由此可见,河竹对于相对淹水深度不超过竹苗地上部高度10%的长期淹水环境具有较强的耐受能力。总体来看,淹水30~180 d,2个处理组河竹叶片的Pn值、蒸腾速率(Tr)、气孔导度(Gs)、胞间CO2浓度(Ci)和叶面水汽压亏缺(Vpdl)均低于CK组,而气孔限制值(Ls)却高于CK组;淹水270和360 d,2个处理组的Pn、Tr、Gs、Ls和Vpdl值均低于CK组,仅Ci值高于CK组。说明在淹水处理的前期(30和90 d)和中期(180 d),河竹叶片Pn值下降的主要原因为气孔限制;而在淹水处理的后期(270和360 d),其Pn值下降的主要原因则是叶肉细胞光合活性下降,即非气孔限制。
最大净光合速率(Pmax)、表观量子效率(AQY)、光饱和点(LSP)和光补偿点(LCP)4项光合特征参数是判定植物光合机构运转状况、光合作用能力、光合利用率及受环境影响程度的重要指标[32-34]。其中,AQY是反映植物光能转化效率的指标之一,其与光饱和时的Pmax值存在显著正相关。本研究中,CK组河竹叶片的AQY值为0.033~0.065 μmol·μmol-1,与自然条件下植物叶片的AQY值(0.03~0.06)[35]基本一致;而在淹水条件下,河竹叶片的AQY值基本上均低于CK组,且不同淹水时间T1组叶片的AQY值与CK组大多无显著差异,但T2组叶片的AQY值与CK组均有显著差异,说明在中度淹水条件下,河竹叶片的光能转化效率高,光合能力强。LSP和LCP分别代表强光与光合作用关系的上限和下限临界指标,能反映植物叶片对光辐射的利用范围[31]。本研究中,淹水90 d时T1组河竹叶片的LSP和LCP值均较CK组显著升高,说明在中度淹水条件下,河竹叶片对光辐射的利用范围增大,对光环境变化的适应能力更强,表明一定程度的淹水处理可能对河竹的光能利用能力无明显影响。
植物的资源利用效率体现在植物对周围环境中光能、水分、氮素和矿质元素等资源的利用水平上。叶片的表观光能利用效率(LUE)反映了植物对光能的捕获和利用效率,能够决定植物能否适应环境并生存[36];叶片的水分利用效率(WUE)是一个衡量植物碳固定和水分消耗能力的较稳定指标[37];而表观CO2利用效率(CUE)则能够反映植物将吸收的碳素转化为生长和贮存的营养物质的能力[38]。在淹水条件下,植物的耐淹能力同样取决于LUE、WUE和CUE值[39-40]。本研究中,淹水30~180 d,2个淹水处理组河竹叶片的WUE、CUE和LUE值均与CK组无显著差异,但在淹水270和360 d则显著低于CK组。表明在淹水条件下,河竹叶片的资源利用效率与淹水时间长短有关,其中,淹水180 d内,河竹叶片的资源利用效率没有受到显著影响;淹水180 d以上,其叶片对水分、光能和CO2的利用效率均受到显著影响。
植物叶片中的叶绿素(Chl)和类胡萝卜素(Car)能够参与光合作用过程中对光能的吸收、传递和转化过程,并能够随着环境的变化动态调整其比例,利于植物叶片更加合理地分配和耗散光能[41-43]。本研究中,淹水30 d时各淹水处理组河竹叶片的Chla含量或高于或低于CK组,但均与CK组无显著差异,说明此时淹水处理对河竹叶片光能转化能力的影响较小;但同一时期,中度淹水可导致河竹叶片的Chlb和Car含量较CK组显著升高,而重度淹水也可使河竹叶片的Chlb和Car含量仅较CK组略升高,说明淹水条件下河竹叶片对光能的吸收和传递能力明显增强。淹水180~360 d,河竹叶片的总叶绿素含量显著低于CK组,表明长期淹水处理可导致河竹叶片中的叶绿素加速分解。淹水30 d,2个处理组间河竹叶片的Chla/Chlb值无显著差异但均显著低于CK组;而淹水360 d,T1组的Chla/Chlb值略高于CK组,而T2组的Chla/Chlb值则显著低于CK组,由于Chla/Chlb值能够反映植物叶片的光合作用强度[44],由此说明在重度淹水条件下,长期淹水可导致河竹叶片光合作用强度下降。淹水30 d,2个处理组河竹叶片的Chl/Car值显著低于或略低于CK组;而淹水360 d,2个处理组的Chl/Car值均显著低于CK组,推测短期淹水造成的河竹叶片Chl/Car值下降可能与叶片中Car含量升高有关,而长期淹水导致的叶片Chl/Car值下降可能与叶片中Chl含量降低、光合色素合成及积累遭到破坏有关。
综上所述,不同程度淹水条件下,河竹叶片对光能的捕获、传递和利用能力均受到一定程度的影响,河竹叶片的光合气体交换参数、光合特征参数、资源利用效率参数和光合色素含量均随淹水时间延长呈“升高—降低—升高”的变化趋势,并且多数参数在淹水90 d时达到峰值、在淹水270 d时达到谷值,其中Pn值的谷值仍达到CK组的50%以上。比较而言,中度淹水(水面高出基质表面5 cm)条件下河竹叶片的各项光合生理参数总体上高于重度淹水(水面高出基质表面10 cm)条件,说明淹水程度越重,河竹叶片的光合生理参数受到的影响越大。总体上看,河竹对淹水环境具有较强的耐受性,对长期淹水也具有一定的耐受能力。
[1] 吴统贵, 周和锋, 吴 明, 等. 旱柳光合作用动态及其与环境因子的关系[J]. 生态学杂志, 2008, 27(12): 2056-2061.
[2] CHAVES M M , PEREIRA J S, MAROCO J, et al. How plants cope with water stress in the field. Photosynthesis and growth[J]. Annals of Botany, 2002, 89: 907-916.
[3] 赵竑绯, 赵 阳, 张 驰, 等. 模拟淹水对杞柳生长和光合特性的影响[J]. 生态学报, 2013, 33(3): 898-906.
[4] 衣英华, 樊大勇, 谢宗强, 等. 模拟淹水对池杉和栓皮栎光合生理生态过程的影响[J]. 生态学报, 2008, 28(12): 6021-6033.
[5] 马瑞娟, 张斌斌, 蔡志翔, 等. 不同桃砧木品种对淹水的光合响应及其耐涝性评价[J]. 园艺学报, 2013, 40(3): 409-416.
[6] 董 蕾, 李吉跃. 植物干旱胁迫下水分代谢、碳饥饿与死亡机理[J]. 生态学报, 2013, 33(18): 5477-5483.
[7] ASHRAF M, ARFAN M. Gas exchange characteristics and water relations in two cultivars ofHibiscusesculentusunder waterlogging[J]. Biologia Plantarum, 2005, 49: 459-462.
[8] KOZLOWSKI T T. Responses of woody plants to flooding and salinity[J]. Tree Physiology Monograph, 1997, 1: 1-29.
[9] 朱桂才, 杨中艺. 水分胁迫下李氏禾叶绿素含量变化研究[J]. 山东农业科学, 2009(11): 48-50.
[10] 衣英华, 樊大勇, 谢宗强, 等. 模拟淹水对枫杨和栓皮栎气体交换、叶绿素荧光和水势的影响[J]. 植物生态学报, 2006, 30(6): 960-968.
[11] PARELLEJ,ZAPATERM,SCOTTI-SAINTAGNEC, et al. Quantitative trait loci of tolerance to waterlogging in a European oak (QuercusroburL.): physiological relevance and temporal effectpatterns[J].Plant,CellandEnvironment, 2007, 30: 422-434.
[12] 吴 静, 王小德, 叶建峰, 等. 涝渍胁迫对鸡爪槭幼苗光合生理特性的影响[J]. 西北林学院学报, 2014, 30(5): 26-30.
[13] 中国科学院中国植物志编辑委员会. 中国植物志: 第九卷第一分册[M]. 北京: 科学出版社, 1996: 311.
[14] ROBERTSON A I, BACON P, HEAGNEY G. The responses of floodplain primary production to flood frequency and timing[J]. Journal of Applied Ecology, 2001, 38: 126-136.
[15] 刘玉芳, 陈双林, 郭子武, 等. 淹水对河竹鞭根系统生物量分配及异速生长模式的影响[J]. 林业科学研究, 2015, 28(4): 502-507.
[16] 刘玉芳, 陈双林, 李迎春, 等. 河竹鞭根对长期淹水环境的生理响应[J]. 林业科学研究, 2014, 27(5): 621-625.
[17] 刘玉芳, 陈双林, 李迎春, 等. 淹水环境下河竹鞭根养分吸收与积累的适应性调节[J]. 生态学报, 2016, 36(10): 2926-2933.
[18] 张卫强, 贺康宁, 田晶会, 等. 不同土壤水分下侧柏苗木光合特性和水分利用效率的研究[J]. 水土保持研究, 2006, 13(6): 44-47.
[19] 朱启红, 徐冬冬, 夏红霞, 等. 淹水胁迫对滴水观音光合气体交换参数的影响[J]. 水生态学杂志, 2014, 35(5): 91-94.
[20] 郭淑红, 薛 立, 杨振意, 等. 水淹胁迫对4种园林绿化树种幼苗气体交换参数的影响[J]. 华南农业大学学报, 2013, 34(4): 548-552.
[21] 张宪政, 陈凤玉, 王荣富. 植物生理学实验技术[M]. 沈阳: 辽宁科学技术出版社, 1994: 164-165.
[22] NIJS I, FERRIS R, BLUM H, et al. Stomatal regulation in a changing climate: a field study using free air temperature increase (FATI) and free air CO2enrichment (FACE)[J]. Plant, Cell and Environment, 1997, 20: 1041-1050.
[23] BERRY J, BJÖRKMAN O. Photosynthesis response and adaptation to temperature in higher plants[J]. Annual Review of Plant Physiology, 1980, 31: 491-543.
[24] 何维明, 马风云. 水分梯度对沙地柏幼苗荧光特征和气孔交换的影响[J]. 植物生态学报, 2000, 24(5): 630-634.
[25] 贾中民, 魏 虹, 田晓峰, 等. 长期水淹对枫杨幼苗光合生理和叶绿素荧光特性的影响[J]. 西南大学学报(自然科学版), 2009, 31(5): 124-129.
[27] BASSMANJH, ZWIERJ C. Gas exchange characteristics ofPopulustrichocarpa,PopulusdeltoidesandPopulustrichocarpa×P.deltoidesclones[J]. Tree Physiology, 1991, 8: 145-159.
[28] 张小全, 徐德应. 杉木中龄林不同部位和叶龄针叶光合特性的日变化和季节变化[J]. 林业科学, 2000, 36(3): 19-26.
[29] 王 瑞, 梁坤伦, 周志宇, 等. 不同淹水梯度对紫穗槐的营养生长和生理响应[J]. 草业学报, 2012, 21(1): 149-155.
[30] 王振夏, 巍 虹, 李昌晓, 等. 土壤水分交替变化对湿地松幼苗光合特性的影响[J]. 西北植物学报, 2012, 32(5): 980-987.
[31] 唐晓芬, 王 彬, 王玉杰, 等. 不同酸度模拟酸雨对重庆缙云山2种阔叶树光合特性的影响[J]. 植物资源与环境学报, 2015, 24(4): 45-51.
[32] 徐伟洲, 徐炳成, 段东平, 等. 不同水肥条件下白羊草光合生理生态特征研究Ⅱ. 光响应曲线[J]. 草地学报, 2010, 18(6): 773-779.
[33] 段爱国, 张建国, 何彩云, 等. 干热河谷主要植被恢复树种干季光合光响应生理参数[J]. 林业科学, 2010, 46(3): 59-73.
[34] 蒋冬月, 钱永强, 刘俊祥, 等. 基于光合-光响应特性的柳树优良无性系光能利用效率的评价[J]. 北京林业大学学报, 2015, 37(5): 49-61.
[35] 蹇洪英, 邹寿青. 地毯草的光合特性研究[J]. 广西植物, 2003, 23(2): 181-184.
[36] 刘玉静, 董周焱, 柏新富, 等. 烟台海岸防护林带6树种资源利用效率比较[J]. 山东农业大学学报(自然科学版), 2015, 46(4): 570-575.
[37] 郑淑霞, 上官周平. 8种阔叶树种叶片气体交换特征和叶绿素荧光特性比较[J]. 生态学报, 2006, 26(4): 1080-1087.
[38] 宋英石, 李 锋, 王效科, 等. 城市地表硬化对银杏生境及生理生态特征的影响[J]. 生态学报, 2014, 34(8): 2164-2171.
[39] CRAWFORD R M M, BRAENDLE R. Oxygen deprivation stress in a changing environment[J]. Journal of Experimental Botany, 1996, 47: 145-159.
[40] 潘 澜, 薛 立. 植物淹水胁迫的生理学机制研究进展[J]. 生态学杂志, 2012, 31(10): 2662-2672.
[41] RONZHINA D A, NEKRASOVA G F, P YANKOV V I. Compara-tive characterization of the pigment complex in emergent, floating, and submerged leaves of hydrophytes[J]. Russian Journal of Plant Physiology, 2004, 51: 21-27.
[42] 李玉灵, 朱 帆, 王俊刚, 等. 水分胁迫下臭柏(SabinavulgarisAnt.)光合特性和色素组成的季节变化[J]. 生态学报, 2009, 29(8): 4346-4352.
[43] 韦继光, 曾其龙, 姜燕琴, 等. 水分胁迫及恢复正常水分供应后兔眼蓝浆果生长及光合特性的变化[J]. 植物资源与环境学报, 2015, 24(3): 77-84.
[44] 何 军, 许 兴, 李树华, 等. 水分胁迫对牛心朴子叶片光合色素及叶绿素荧光的影响[J]. 西北植物学报, 2004, 24(9): 1594-1598.
(责任编辑: 佟金凤)
Changes in photosynthetic and physiological characteristics of leaf of Phyllostachys rivalis under different flooding conditions
LI Yingchun1,2, GUO Ziwu2, CHEN Shuanglin2,①, YUE Yongde1, LIU Yufang2, YANG Qingping2
(1. InternationalCenterforBambooandRattan, Beijing 100102, China; 2. Research Institute of Subtropical Forestry, Chinese Academy of Forestry, Hangzhou 311400, China),J.PlantResour. &Environ., 2016, 25(4): 42-52
In order to clarify photosynthetic and physiological characteristics of leaf ofPhyllostachysrivalisH. R. Zhao under flooding condition, the control group (CK, normal water supplying), moderate flooding (T1, water surface higher than substrate surface 5 cm) and severe flooding (T2, water surface higher than substrate surface 10 cm) treatment groups were set up, changes in photosynthetic gas exchange parameters, light response curve, photosynthetic characteristic parameters, resource use efficiency parameters, contents and ratios of photosynthetic pigments in leaf of two-year-old seedling ofP.rivalisduring continuously flooding for 360 d were analyzed. On this basis, adaptability ofP.rivalisto flooding environment was discussed. The results show that with prolonging of flooding time, most photosynthetic and physiological parameters of two treatment groups appear the changing trend of “increasing-decreasing-increasing”, and most parameters reach the peak value when flooding for 90 d and reach the valley value when flooding for 270 d. Various photosynthetic and physiological parameters of T1 group are generally higher than those of T2 group. When flooding for 30-180 d, compared with CK group, net photosynthetic rate (Pn), transpiration rate (Tr), stomatal conductance (Gs), intercellular CO2concentration (Ci) and vapor pressure deficit of leaf surface (Vpdl) of two treatment groups decrease generally, apparent quantum yield (AQY) decreases significantly; limited stomatal value (Ls) increases, while light saturation point (LSP) and light compensation point (LCP) increase significantly; the maximum net photosynthetic rate (Pmax) of T1 group decreases slightly, while that of T2 group decreases significantly; but there are not obvious changes in water use efficiency (WUE), apparent CO2use efficiency (CUE) and apparent light use efficiency (LUE) of two treatment groups. When flooding for 270 and 360 d, compared with CK group, Pn, Tr, Gs, Ls and Vpdl values of two treatment groups decrease, while their Pmax, LSP, LCP, WUE, CUE and LUE values decrease significantly, only Ci value increases; AQY value of T1 group decreases slightly, while that of T2 group decreases significantly. Changing trend of light response curve of two treatment groups is similar to that of CK group, but changing range of Pn value of two treatment groups is different with prolonging of flooding time, in which, Pn value of T1 group is generally higher than that of T2 group. When flooding for 30 and 90 d, compared with CK group, contents of chlorophyllb(Chlb) and total chlorophyll (Chl) of T1 group increase significantly in general, while those of T2 group decrease significantly in general; contents of chlorophylla(Chla) and carotenoid (Car) and Chla/Chlband Chl/Car values of two treatment groups also decrease significantly. When flooding for 180-360 d, only Chla/Chlbvalue of two treatment groups increases significantly, while other photosynthetic pigment contents and ratios decrease significantly in general. The comprehensive analysis result shows that the main reason for decreasing of Pn value of leaf ofP.rivalisat early and middle flooding stages is stomatal limitation, while that at later stage is mainly non-stomatal limitation.P.rivalishas stronger tolerance to flooding environment, and has a certain adaptability to long-term flooding. In addition, efficiency of light energy conversion, utilization range to light radiation and adaptability to light change of leaf ofP.rivalisunder moderate flooding condition all are higher than those under severe flooding condition.
PhylllostachysrivalisH. R. Zhao; flooding; photosynthetic gas exchange parameter; photosynthetic characteristics; resource use efficiency parameter; photosynthetic pigment
2016-05-04
浙江省省院合作项目(2012SY05); 浙江省自然科学基金资助项目(LY13C160001); 中央级公益性科研院所基本科研业务费专项资金项目(RISF61258)
李迎春(1976—),女,山东高唐人,博士研究生,助理研究员,主要研究方向为竹林生理生态。
①通信作者E-mail: cslbamboo@126.com
Q945.11; Q948.112+.3; S795.9
A
1674-7895(2016)04-0042-11
10.3969/j.issn.1674-7895.2016.04.06
